The Absence of Pyruvate Kinase Affects Glucose-Dependent Carbon Catabolite Repression in Bacillus subtilis



Abstract
:1. Introduction
2. Results
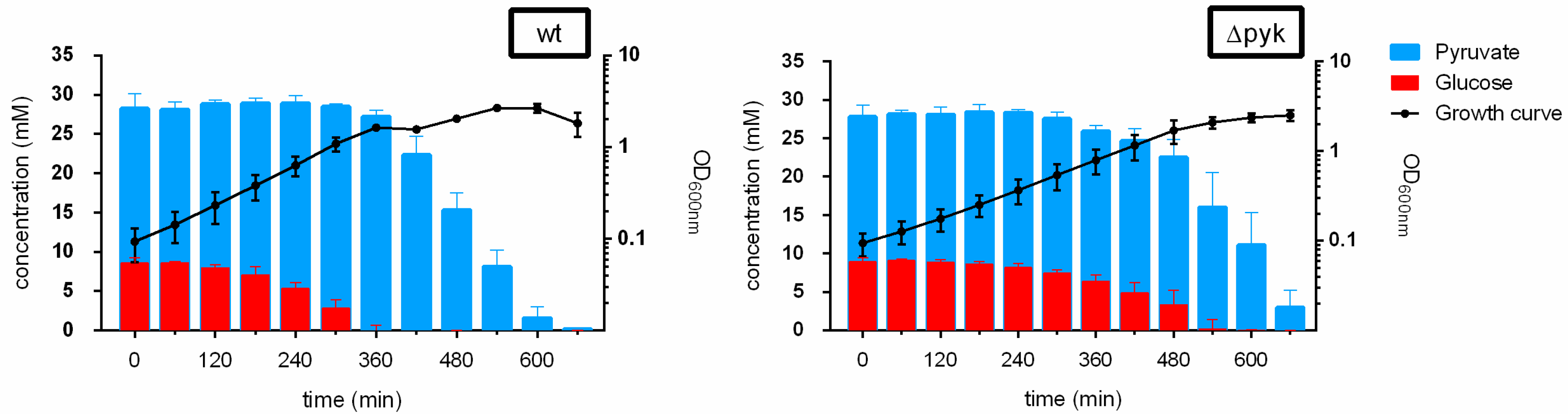
2.1. Concomitant Consumption of Glucose and Pyruvate by ΔPyk—Carbon Catabolite Repression of Pyruvate Uptake in the Presence of Glucose Was Relieved
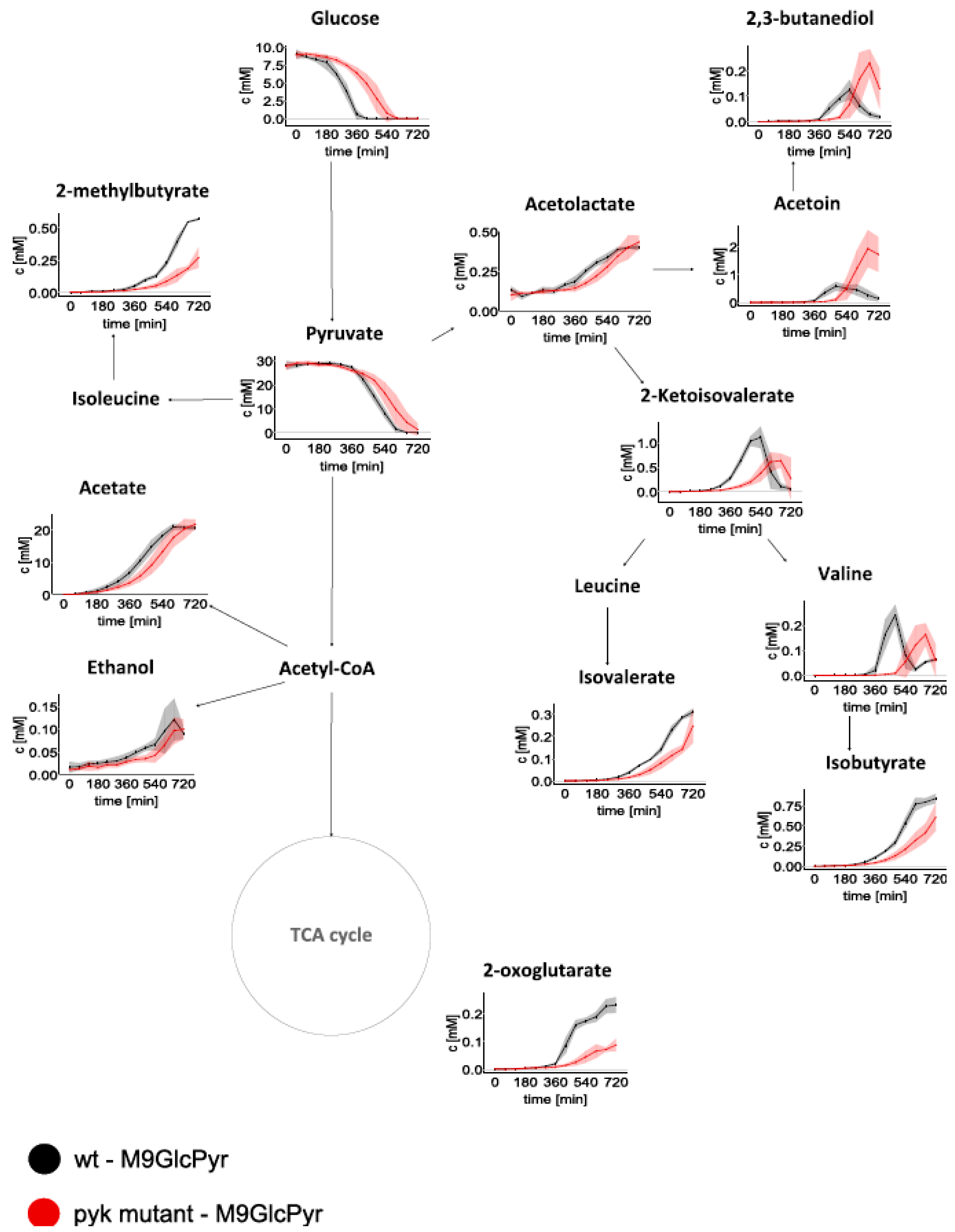
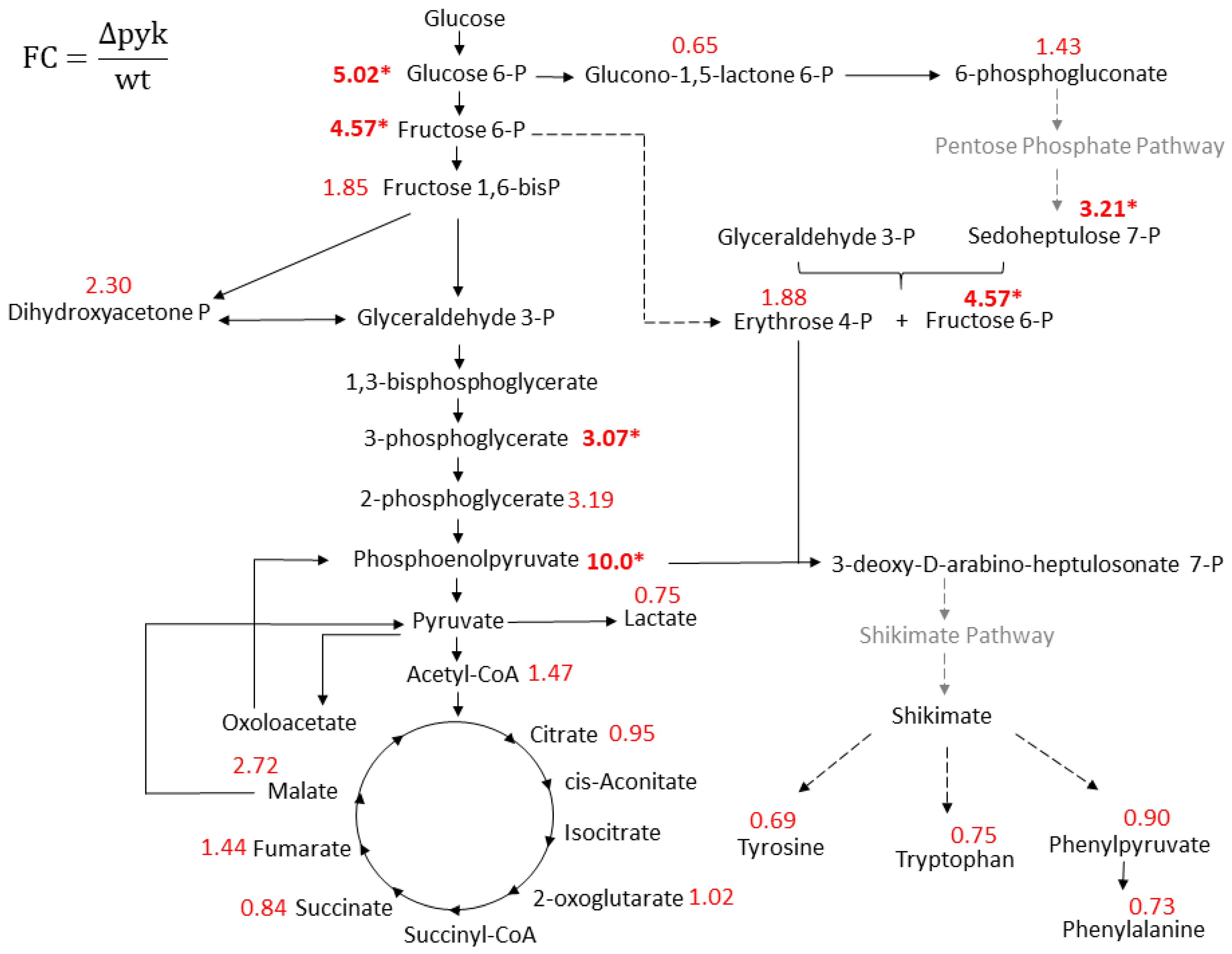
2.2. Intracellular Metabolome Revealed Several Central Metabolic Pathways Altered in ΔPyk
3. Discussion
3.1. Carbon Catabolite Repression of Pyruvate Uptake in ΔPyk Cells Was Relieved
3.2. The TCA cycle and the Pentose Phosphate Pathway (PPP) Are Possibly Altered in Δpyk
3.3. Overflow Metabolites: Increased Secretion of Acetoin and 2,3-Butanediol by Δpyk
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Cultivation
4.3. Sampling of Extracellular and Intracellular Metabolites
4.4. H-NMR Spectroscopy Measurement and Data Analysis of Extracellular Metabolites
4.5. GC-MS Measurement and Data Analysis of Intracellular Metabolites
4.6. HPLC-MS Measurement and Data Analysis of Intracellular Metabolites
4.7. LC-MS/MS Measurement and Data Analysis of Intracellular Metabolites
4.8. Statistical Analysis and Visualization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kunst, F.; Ogasawara, N.; Moszer, I.; Albertini, A.M.; Alloni, G.O.; Azevedo, V.; Bertero, M.G.; Bessieres, P.; Bolotin, A.; Borchert, S.; et al. The complete genome sequence of the gram-positive bacterium Bacillus subtilis. Nature 1997, 390, 249–256. [Google Scholar] [CrossRef]
- Barbe, V.; Cruveiller, S.; Kunst, F.; Lenoble, P.; Meurice, G.; Sekowska, A.; Vallenet, D.; Wang, T.; Moszer, I.; Médigue, C.; et al. From a consortium sequence to a unified sequence: The Bacillus subtilis 168 reference genome a decade later. Microbiology 2009, 155, 1758–1775. [Google Scholar] [CrossRef]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Modern analytical techniques in metabolomics analysis. Analyst 2012, 137, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Sauer, U.; Eikmanns, B.J. The PEP—pyruvate—oxaloacetate node as the switch point for carbon flux distribution in bacteria: We dedicate this paper to Rudolf K. Thauer, Director of the Max-Planck-Institute for Terrestrial Microbiology in Marburg, Germany, on the occasion of his 65th birthday. FEMS Microbiol. Rev. 2005, 29, 765–794. [Google Scholar] [PubMed]
- Goel, A.; Lee, J.; Domach, M.M.; Ataai, M.M. Metabolic fluxes, pools, and enzyme measurements suggest a tighter coupling of energetics and biosynthetic reactions associated with reduced pyruvate kinase flux. Biotechnol. Bioeng. 1999, 64, 129–134. [Google Scholar] [CrossRef]
- Pan, Z.; Zhu, T.; Domagalski, N.; Khan, S.; Koepsel, R.R.; Domach, M.M.; Ataai, M.M. Regulating expression of pyruvate kinase in Bacillus subtilis for control of growth rate and formation of acidic byproducts. Biotechnol. Prog. 2006, 22, 1451–1455. [Google Scholar] [CrossRef] [PubMed]
- Charbonnier, T.; Le Coq, D.; McGovern, S.; Calabre, M.; Delumeau, O.; Aymerich, S.; Jules, M. Molecular and Physiological Logics of the Pyruvate-Induced Response of a Novel Transporter in Bacillus subtilis. MBio 2017, 8, e00976-17. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, D.; De Mendoza, D. FapR: From Control of Membrane Lipid Homeostasis to a Biotechnological Tool. Front. Mol. Biosci. 2016, 3, 64. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y. Carbon catabolite control of the metabolic network in Bacillus subtilis. Biosci. Biotechnol. Biochem. 2009, 73, 245–259. [Google Scholar] [CrossRef]
- Görke, B.; Stülke, J. Carbon catabolite repression in bacteria: Many ways to make the most out of nutrients. Nat. Rev. Microbiol. 2008, 6, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Kleijn, R.J.; Buescher, J.M.; Le Chat, L.; Jules, M.; Aymerich, S.; Sauer, U. Metabolic fluxes during strong carbon catabolite repression by malate in Bacillus subtilis. J. Biol. Chem. 2010, 285, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.D.; Schmalisch, M.H.; Stülke, J.; Görke, B. Carbon catabolite repression in Bacillus subtilis: Quantitative analysis of repression exerted by different carbon sources. J. Bacteriol. 2008, 190, 7275–7284. [Google Scholar] [CrossRef] [PubMed]
- van den Esker, M.H.; Kovács, Á.T.; Kuipers, O.P. YsbA and LytST are essential for pyruvate utilization in Bacillus subtilis. Environ. Microbiol. 2017, 19, 83–94. [Google Scholar] [CrossRef] [PubMed]
- van den Esker, M.H.; Kovács, Á.T.; Kuipers, O.P. From Cell Death to Metabolism: Holin-Antiholin Homologues with New Functions. MBio 2017, 8, e01963-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutscher, J.; Francke, C.; Postma, P.W. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 939–1031. [Google Scholar] [CrossRef] [PubMed]
- Postma, P.; Lengeler, J.; Jacobson, G. Phosphoenolpyruvate: Carbohydrate phosphotransferase systems of bacteria. Microbiol. Rev. 1993, 57, 543–594. [Google Scholar]
- Li, Z.; Nimtz, M.; Rinas, U. The metabolic potential of Escherichia coli BL21 in defined and rich medium. Microb. Cell Fact. 2014, 13, 45. [Google Scholar] [CrossRef]
- Sonenshein, A.L. Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Microbiol. 2007, 5, 917–927. [Google Scholar] [CrossRef]
- Ponce, E.; Martínez, A.; Bolívar, F.; Valle, F. Stimulation of glucose catabolism through the pentose pathway by the absence of the two pyruvate kinase isoenzymes in Escherichia coli. Biotechnol. Bioeng. 1998, 58, 292–295. [Google Scholar] [CrossRef]
- Emmerling, M.; Dauner, M.; Ponti, A.; Fiaux, J.; Hochuli, M.; Szyperski, T.; Wüthrich, K.; Bailey, J.E.; Sauer, U. Metabolic flux responses to pyruvate kinase knockout in Escherichia coli. J. Bacteriol. 2002, 184, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Meza, E.; Becker, J.; Bolivar, F.; Gosset, G.; Wittmann, C. Consequences of phosphoenolpyruvate: Sugar phosphotranferase system and pyruvate kinase isozymes inactivation in central carbon metabolism flux distribution in Escherichia coli. Microb. Cell Fact. 2012, 11, 127. [Google Scholar] [CrossRef] [PubMed]
- Toya, Y.; Ishii, N.; Nakahigashi, K.; Hirasawa, T.; Soga, T.; Tomita, M.; Shimizu, K. 13C-metabolic flux analysis for batch culture of Escherichia coli and its pyk and pgi gene knockout mutants based on mass isotopomer distribution of intracellular metabolites. Biotechnol. Prog. 2010, 26, 975–992. [Google Scholar] [PubMed]
- Schilling, O.; Frick, O.; Herzberg, C.; Ehrenreich, A.; Heinzle, E.; Wittmann, C.; Stülke, J. Transcriptional and metabolic responses of Bacillus subtilis to the availability of organic acids: Transcription regulation is important but not sufficient to account for metabolic adaptation. Appl. Environ. Microbiol. 2007, 73, 499–507. [Google Scholar] [CrossRef]
- Dauner, M.; Storni, T.; Sauer, U. Bacillus subtilis metabolism and energetics in carbon-limited and excess-carbon chemostat culture. J. Bacteriol. 2001, 183, 7308–7317. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Altman, E.; Sangurdekar, D.P.; Khodursky, A.B.; Eiteman, M.A. Overflow metabolism in Escherichia coli during steady-state growth: Transcriptional regulation and effect of the redox ratio. Appl. Environ. Microbiol. 2006, 72, 3653–3661. [Google Scholar] [CrossRef]
- Szenk, M.; Dill, K.A.; de Graff, A.M.R. Why do fast-growing bacteria enter overflow metabolism? Testing the membrane real estate hypothesis. Cell Syst. 2017, 5, 95–104. [Google Scholar] [CrossRef]
- Wolfe, A.J. The acetate switch. Microbiol. Mol. Biol. Rev. 2005, 69, 12–50. [Google Scholar] [CrossRef]
- Grundy, F.; Waters, D.; Allen, S.; Henkin, T. Regulation of the Bacillus subtilis acetate kinase gene by CcpA. J. Bacteriol. 1993, 175, 7348–7355. [Google Scholar] [CrossRef]
- Meyer, H.; Weidmann, H.; Mäder, U.; Hecker, M.; Völker, U.; Lalk, M. A time resolved metabolomics study: The influence of different carbon sources during growth and starvation of Bacillus subtilis. Mol. Biosyst. 2014, 10, 1812–1823. [Google Scholar] [CrossRef]
- Cabrera-Valladares, N.; Martínez, L.M.; Flores, N.; Hernández-Chávez, G.; Martínez, A.; Bolívar, F.; Gosset, G. Physiologic consequences of glucose transport and phosphoenolpyruvate node modifications in Bacillus subtilis 168. J. Mol. Microbiol. Biotechnol. 2012, 22, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Xu, P. Acetoin metabolism in bacteria. Crit. Rev. Microbiol. 2007, 33, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Ramos, H.C.; Hoffmann, T.; Marino, M.; Nedjari, H.; Presecan-Siedel, E.; Dreesen, O.; Glaser, P.; Jahn, D. Fermentative metabolism of Bacillus subtilis: Physiology and regulation of gene expression. J. Bacteriol. 2000, 182, 3072–3080. [Google Scholar] [CrossRef] [PubMed]
- Turinsky, A.J.; Grundy, F.J.; Kim, J.-H.; Chambliss, G.H.; Henkin, T.M. Transcriptional activation of the Bacillus subtilis ackA gene requires sequences upstream of the promoter. J. Bacteriol. 1998, 180, 5961–5967. [Google Scholar]
- Härtig, E.; Jahn, D. Regulation of the anaerobic metabolism in Bacillus subtilis. Adv. Microb. Physiol. 2012, 61, 195–216. [Google Scholar]
- Harwood, C.R.; Cutting, S.M. Molecular Biological Methods for Bacillus; Wiley: Hoboken, NJ, USA, 1990. [Google Scholar]
- Sambrook, J. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 2001. [Google Scholar]
- Wach, A. PCR-synthesis of marker cassettes with long flanking homology regions for gene disruptions in S. cerevisiae. Yeast 1996, 12, 259–265. [Google Scholar] [CrossRef]
- Pietak, N.; Becher, D.; Schmidl, S.R.; Saier, M.H.; Hecker, M.; Commichau, F.M.; Stülke, J. In vitro Phosphorylation of Key Metabolic Enzymes from Bacillus subtilis: PrkC Phosphorylates Enzymes from Different Branches of Basic Metabolism. J. Mol. Microbiol. Biotechnol. 2010, 18, 129–140. [Google Scholar] [CrossRef]
- Guerout-Fleury, A.-M.; Shazand, K.; Frandsen, N.; Stragier, P. Antibiotic-resistance cassettes for Bacillus subtilis. Genes 1995, 167, 335–336. [Google Scholar] [CrossRef]
- Meyer, H.; Weidmann, H.; Lalk, M. Methodological approaches to help unravel the intracellular metabolome of Bacillus subtilis. Microb. Cell Fact. 2013, 12, 69. [Google Scholar] [CrossRef]
- Atkinson, D.E. Energy charge of the adenylate pool as a regulatory parameter. Interaction with feedback modifiers. Biochemistry 1968, 7, 4030–4034. [Google Scholar] [CrossRef]
- Dörries, K.; Schlueter, R.; Lalk, M. Impact of antibiotics with various target sites on the metabolome of Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 7151–7163. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | wt | Δpyk | ||||||
|---|---|---|---|---|---|---|---|---|
| Glucose | Pyruvate | Glucose | Pyruvate | |||||
| Concentration (mM) | SD | Concentration (mM) | SD | Concentration (mM) | SD | Concentration (mM) | SD | |
| 0 | 8.59 | 0.65 | 28.25 | 1.85 | 8.93 | 0.53 | 27.80 | 1.52 |
| 60 | 8.62 | 0.17 | 28.10 | 0.98 | 9.09 | 0.20 | 28.16 | 0.48 |
| 120 | 7.98 | 0.38 | 28.80 | 0.49 | 8.88 | 0.33 | 28.10 | 0.93 |
| 180 | 7.05 | 1.04 | 28.93 | 0.62 | 8.63 | 0.31 | 28.41 | 0.99 |
| 240 | 5.31 | 0.73 | 28.91 | 0.98 | 8.20 | 0.48 | 28.33 | 0.44 |
| 300 | 2.82 | 1.09 | 28.49 | 0.38 | 7.49 | 0.41 | 27.55 | 0.82 |
| 360 | 0.00 | 0.68 | 27.24 | 0.81 | 6.41 | 0.80 | 25.94 | 0.73 |
| 420 | 0.00 | 0.00 | 22.34 | 2.35 | 4.90 | 1.33 | 24.69 | 1.55 |
| 480 | 0.00 | 0.04 | 15.32 | 2.24 | 3.35 | 1.91 | 22.53 | 2.35 |
| 540 | 0.01 | 0.01 | 8.14 | 2.10 | 0.20 | 1.20 | 16.03 | 4.51 |
| 600 | 0.00 | 0.02 | 1.59 | 1.39 | 0.05 | 0.04 | 11.14 | 4.20 |
| 660 | 0.02 | 0.02 | 0.20 | 0.04 | 0.03 | 0.03 | 2.99 | 2.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, J.; Westhoff, P.; Methling, K.; Lalk, M. The Absence of Pyruvate Kinase Affects Glucose-Dependent Carbon Catabolite Repression in Bacillus subtilis. Metabolites 2019, 9, 216. https://doi.org/10.3390/metabo9100216
Sousa J, Westhoff P, Methling K, Lalk M. The Absence of Pyruvate Kinase Affects Glucose-Dependent Carbon Catabolite Repression in Bacillus subtilis. Metabolites. 2019; 9(10):216. https://doi.org/10.3390/metabo9100216
Chicago/Turabian StyleSousa, Joana, Philipp Westhoff, Karen Methling, and Michael Lalk. 2019. "The Absence of Pyruvate Kinase Affects Glucose-Dependent Carbon Catabolite Repression in Bacillus subtilis" Metabolites 9, no. 10: 216. https://doi.org/10.3390/metabo9100216
APA StyleSousa, J., Westhoff, P., Methling, K., & Lalk, M. (2019). The Absence of Pyruvate Kinase Affects Glucose-Dependent Carbon Catabolite Repression in Bacillus subtilis. Metabolites, 9(10), 216. https://doi.org/10.3390/metabo9100216