A Novel Noncoding RNA dsr11 Involved in Heat Stress Tolerance in Deinococcus radiodurans

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Growth Conditions
2.2. RNA Extraction
2.3. Construction of Gene Deletion Mutant Strain
2.4. Quantitative Real-Time PCR (qRT-PCR)
2.5. Bacterial Growth Curve and Heat Stress Tolerance Assays
2.6. RNA-seq and Data Analysis
2.7. Bioinformatics Analysis
2.8. Microscale Thermophoresis (MST) Analysis
3. Results and Discussion
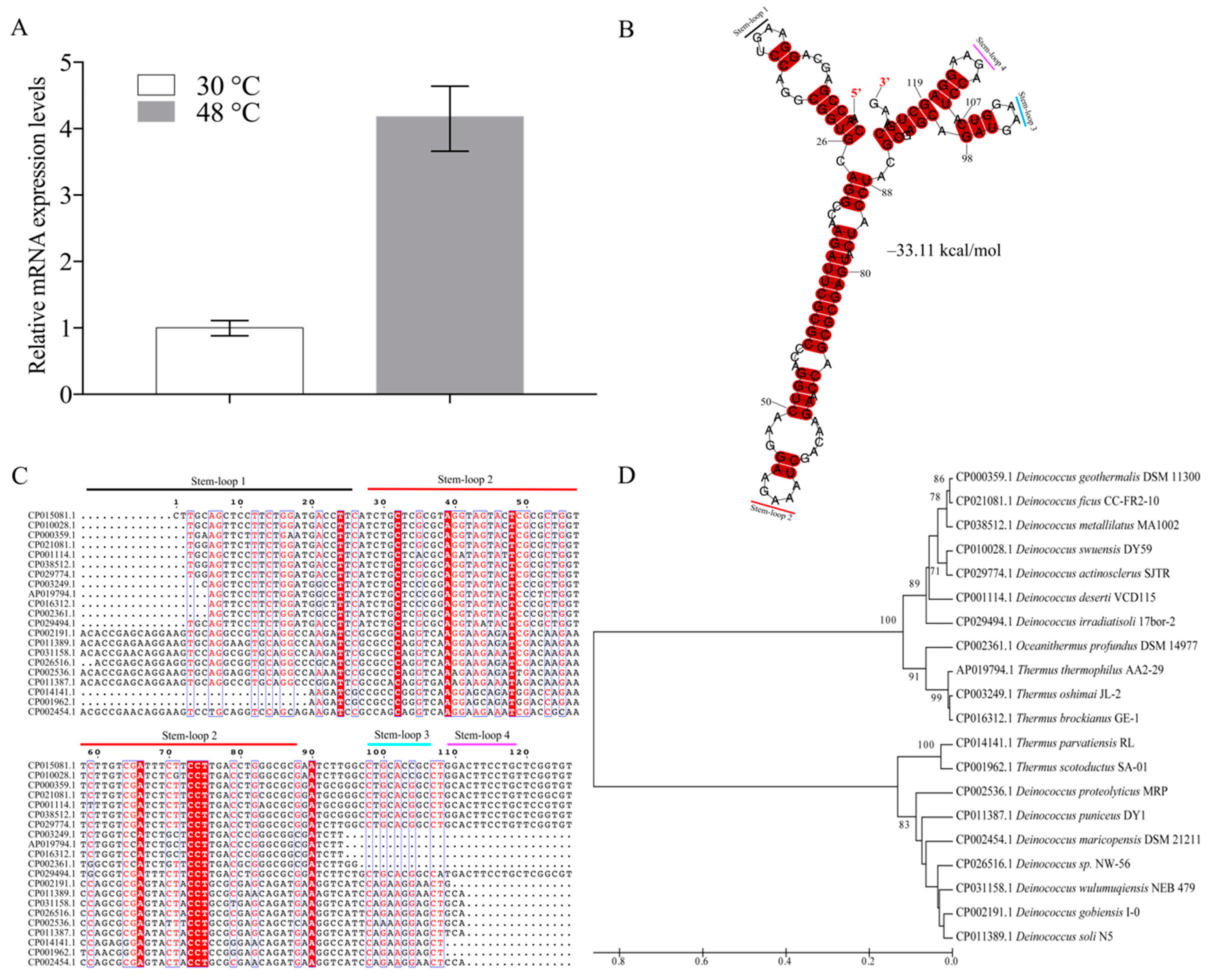
3.1. Characterization of the Novel ncRNA dsr11
3.2. dsr11 is Required for Heat Tolerance of D. radiodurans
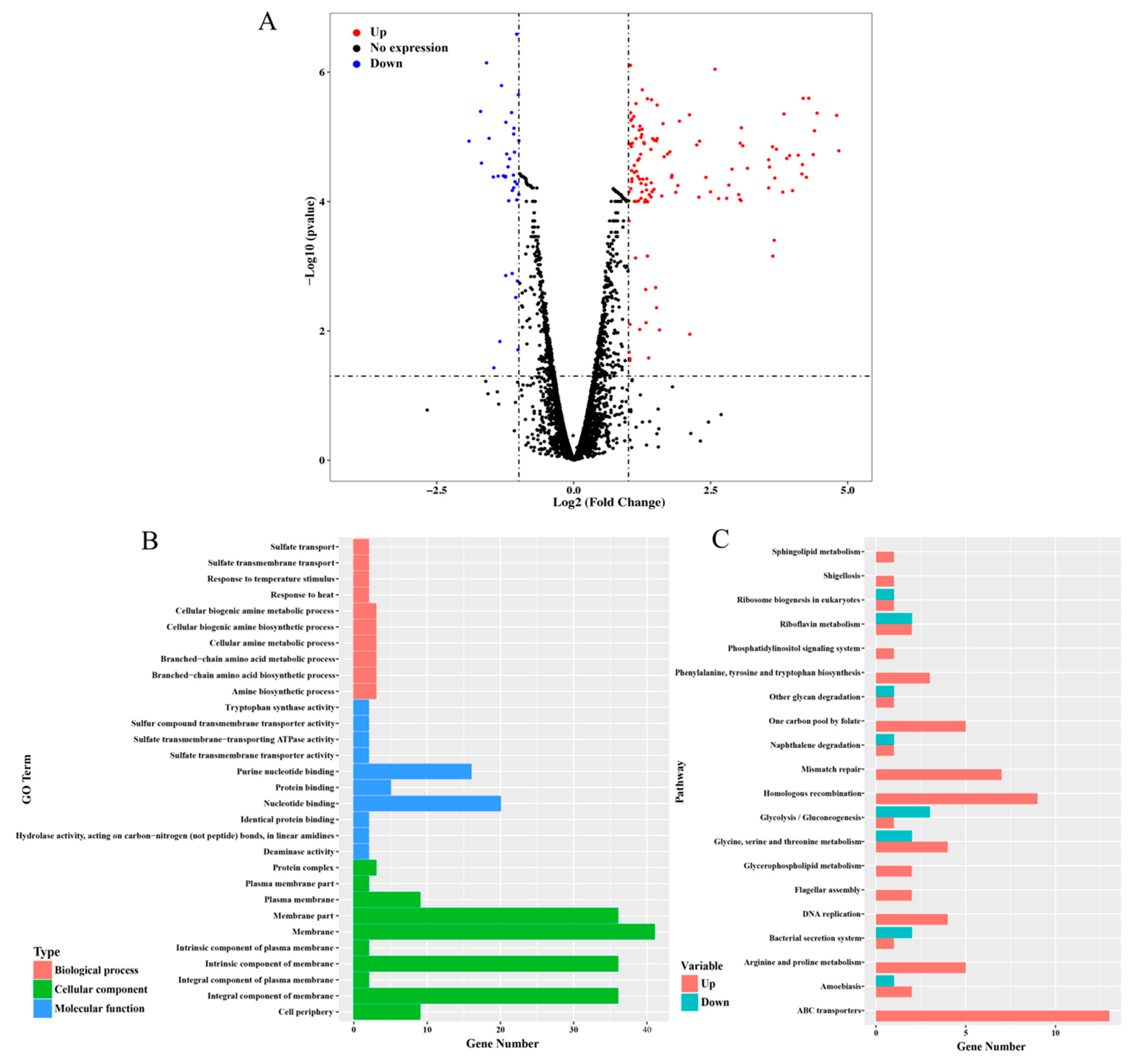
3.3. RNA-seq Analysis Sheds New Light on the Mechanisms of dsr11-Mediated Heat Tolerance
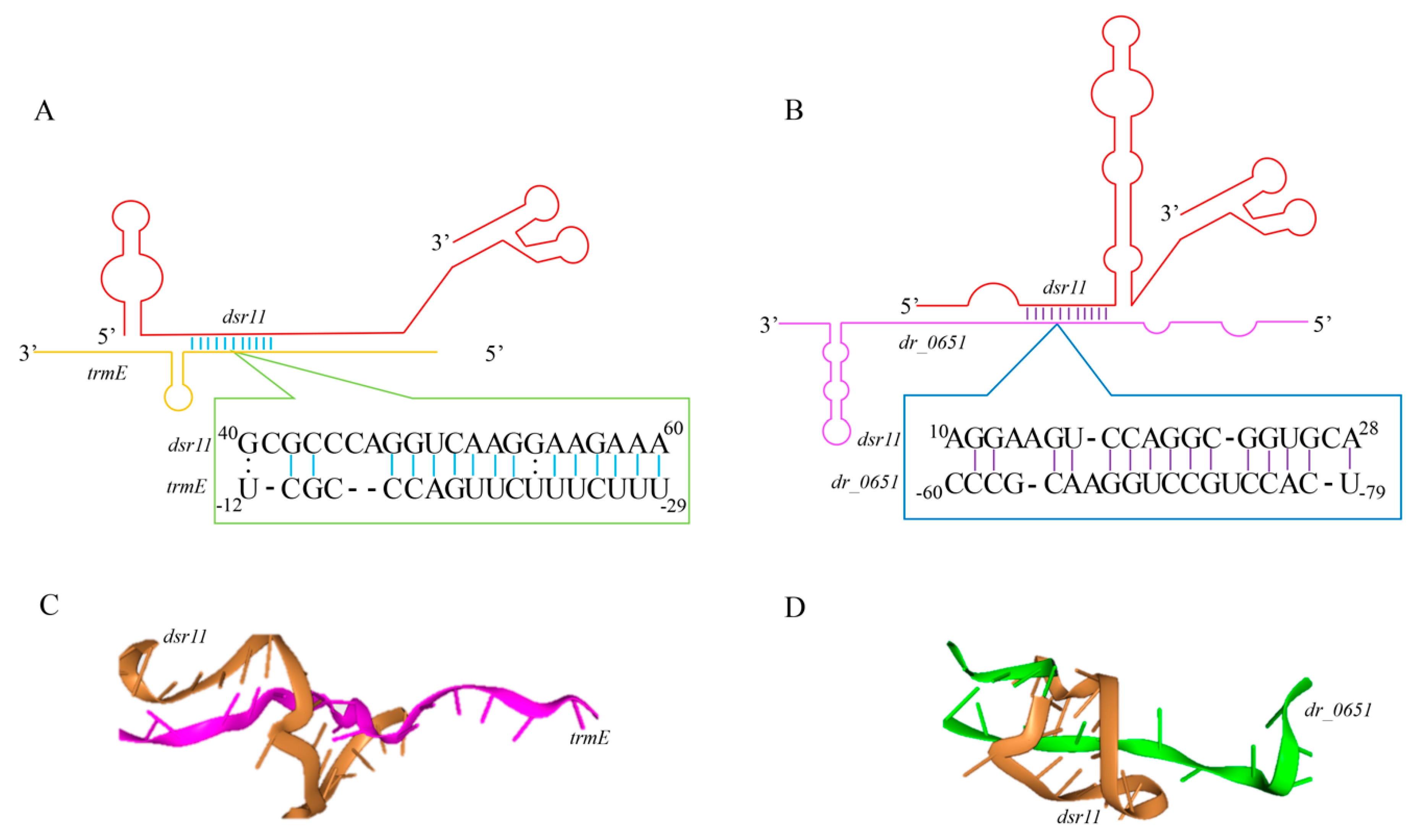
3.4. Identification of the dsr11 Targets
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, M.V.; Wolf, Y.I.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Daly, M.J.; Koonin, E.V.; Makarova, K.S. Comparative genomics of Thermus thermophilus and Deinococcus radiodurans: Divergent routes of adaptation to thermophily and radiation resistance. BMC Evol. Biol. 2005, 5, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Hua, Y. Improved complete genome sequence of the extremely radioresistant bacterium Deinococcus radiodurans R1 obtained using PacBio singlemolecule sequencing. Genome Announc. 2016, 4, e00886-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, L.; Coste, G.; Sommer, S.; Oberto, J.; Confalonieri, F.; Servant, P.; Pasternak, C. DdrI, a cAMP receptor protein family member, acts as a major regulator for adaptation of Deinococcus radiodurans to various stresses. J. Bacteriol. 2018, 200, e00129-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [Green Version]
- Airo, A.; Chan, S.L.; Martinez, Z.; Platt, M.O.; Trent, J.D. Heat shock and cold shock in Deinococcus radiodurans. Cell Biochem. Biophys. 2004, 40, 277–288. [Google Scholar] [CrossRef]
- Patel, B.A.; Moreau, M.; Widom, J.; Chen, H.; Yin, L.; Hua, Y.; Crane, B.R. Endogenous nitric oxide regulates the recovery of the radiation-resistant bacterium Deinococcus radiodurans from exposure to UV light. Proc. Natl. Acad. Sci. USA 2009, 106, 18183–18188. [Google Scholar] [CrossRef] [Green Version]
- Mattimore, V.; Battista, J.R. Radioresistance of Deinococcus radiodurans: Functions necessary to survive ionizing radiation are also necessary to survive prolonged desiccation. J. Bacteriol. 1996, 178, 633–637. [Google Scholar] [CrossRef] [Green Version]
- Blasius, M.; Sommer, S.; Hübscher, U. Deinococcus radiodurans: What belongs to the survival kit? Crit. Rev. Biochem. Mol. Biol. 2008, 43, 221–238. [Google Scholar] [CrossRef]
- Gottesman, S. Micros for microbes: Non-coding regulatory RNAs in bacteria. Trends Genet. 2005, 21, 399–404. [Google Scholar] [CrossRef]
- Waters, L.S.; Storz, G. Regulatory RNAs in bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by small RNAs in bacteria: Expanding frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, B.; van Opijnen, T.; Wang, J.; Obert, C.; Wang, Y.D.; Carter, R.; McGoldrick, D.J.; Ridout, G.; Camilli, A.; Tuomanen, E.I.; et al. Control of virulence by small RNAs in Streptococcus pneumoniae. PLoS Pathog. 2012, 8, e1002788. [Google Scholar] [CrossRef] [PubMed]
- Opdyke, J.A.; Kang, J.G.; Storz, G. GadY, a small-RNA regulator of acid response genes in Escherichia coli. J. Bacteriol. 2004, 186, 6698–6705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhan, Y.; Yan, Y.; Liu, Y.; Hu, G.; Wang, S.; Yang, H.; Qiu, X.; Liu, Y.; Li, J.; et al. The Pseudomonas stutzeri-specific regulatory ncRNA, NfiS, targets the katB mRNA encoding a catalase essential for optimal oxidative resistance and nitrogenase activity. J. Bacteriol. 2019, 201, e00334-19. [Google Scholar] [CrossRef] [Green Version]
- Lybecker, M.C.; Samuels, D.S. Temperature-induced regulation of RpoS by a small RNA in Borrelia burgdorferi. Mol. Microbiol. 2007, 64, 1075–1089. [Google Scholar] [CrossRef]
- Eyraud, A.; Tattevin, P.; Chabelskaya, S.; Felden, B. A small RNA controls a protein regulator involved in antibiotic resistance in Staphylococcus aureus. Nucleic Acids Res. 2014, 42, 4892–4905. [Google Scholar] [CrossRef]
- Sharma, C.M.; Vogel, J. Experimental approaches for the discovery and characterization of regulatory small RNA. Curr. Opin. Microbiol. 2009, 12, 536–546. [Google Scholar] [CrossRef]
- Puerta-Fernandez, E.; Barrick, J.E.; Roth, A.; Breaker, R.R. Identification of a large noncoding RNA in extremophilic eubacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 19490–19495. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.J.; Misulovin, Z.; Eddy, S.R. Noncoding RNA genes identified in AT-rich hyperthermophiles. Proc. Natl. Acad. Sci. USA 2002, 99, 7542–7547. [Google Scholar] [CrossRef] [Green Version]
- Gelsinger, D.R.; Diruggiero, J. The non-coding regulatory RNA revolution in archaea. Genes 2018, 9, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelsinger, D.R.; DiRuggiero, J. Transcriptional landscape and regulatory roles of small noncoding RNAs in the oxidative stress response of the haloarchaeon Haloferax volcanii. J. Bacteriol. 2018, 200, e00779-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliemt, J.; Jaschinski, K.; Soppa, J. A haloarchaeal small regulatory RNA (sRNA) is essential for rapid adaptation to phosphate starvation conditions. Front. Microbiol. 2019, 10, 1219. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Liao, R.; Chou, B.; Contreras, L.M. Transcriptional analysis of Deinococcus radiodurans reveals novel small RNAs that are differentially expressed under ionizing radiation. Appl. Environ. Microbiol. 2015, 81, 1754–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, D.; Gao, G.; Tian, B.; Xu, Z.; Zheng, Z.; Hua, Y. RecX is involved in antioxidant mechanisms of the radioresistant bacterium Deinococcus radiodurans. FEMS Microbiol. Lett. 2005, 244, 251–257. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder-Open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef] [Green Version]
- Bernhart, S.H.; Hofacker, I.L.; Will, S.; Gruber, A.R.; Stadler, P.F. RNAalifold: Improved consensus structure prediction for RNA alignments. BMC Bioinform. 2008, 9, 474. [Google Scholar] [CrossRef] [Green Version]
- Kery, M.B.; Feldman, M.; Livny, J.; Tjaden, B. TargetRNA2: Identifying targets of small regulatory RNAs in bacteria. Nucleic Acids Res. 2014, 42, W124–W129. [Google Scholar] [CrossRef] [Green Version]
- Beckert, B.; Kedrov, A.; Sohmen, D.; Kempf, G.; Wild, K.; Sinning, I.; Stahlberg, H.; Wilson, D.N.; Beckmann, R. Translational arrest by a prokaryotic signal recognition particle is mediated by RNA interactions. Nat. Struct. Mol. Biol. 2015, 22, 767. [Google Scholar] [CrossRef] [PubMed]
- Buddeweg, A.; Sharma, K.; Urlaub, H.; Schmitz, R.A. sRNA41 affects ribosome binding sites within polycistronic mRNAs in Methanosarcina mazei Gö1. Mol. Microbiol. 2018, 107, 595–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerabek-Willemsen, M.; André, T.; Wanner, R.; Roth, H.M.; Duhr, S.; Baaske, P.; Breitsprecher, D. MicroScale Thermophoresis: Interaction analysis and beyond. J. Mol. Struct. 2014, 1077, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Lippok, S.; Seidel, S.A.I.; Duhr, S.; Uhland, K.; Holthoff, H.P.; Jenne, D.; Braun, D. Direct detection of antibody concentration and affinity in human serum using microscale thermophoresis. Anal. Chem. 2012, 84, 3523–3530. [Google Scholar] [CrossRef]
- Peer, A.; Margalit, H. Accessibility and evolutionary conservation mark bacterial small-RNA target-binding regions. J. Bacteriol. 2011, 193, 1690–1701. [Google Scholar] [CrossRef] [Green Version]
- Miroshnichenko, M.L.; L’Haridon, S.; Jeanthon, C.; Antipov, A.N.; Kostrikina, N.A.; Tindall, B.J.; Schumann, P.; Spring, S.; Stackebrandt, E.; Bonch-Osmolovskaya, E.A. Oceanithermus profundus gen. nov., sp. nov., a thermophilic, microaerophilic, facultatively chemolithoheterotrophic bacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2003, 53, 747–752. [Google Scholar] [CrossRef]
- Bauermeister, A.; Hahn, C.; Rettberg, P.; Reitz, G.; Moeller, R. Roles of DNA repair and membrane integrity in heat resistance of Deinococcus radiodurans. Arch. Microbiol. 2012, 194, 959–966. [Google Scholar] [CrossRef]
- Kawamoto, H.; Koide, Y.; Morita, T.; Aiba, H. Base-pairing requirement for RNA silencing by a bacterial small RNA and acceleration of duplex formation by Hfq. Mol. Microbiol. 2006, 61, 1013–1022. [Google Scholar] [CrossRef]
- Prévost, K.; Salvail, H.; Desnoyers, G.; Jacques, J.F.; Phaneuf, É.; Massé, E. The small RNA RyhB activates the translation of shiA mRNA encoding a permease of shikimate, a compound involved in siderophore synthesis. Mol. Microbiol. 2007, 64, 1260–1273. [Google Scholar] [CrossRef]
- Yamanaka, K.; Hwang, J.; Inouye, M. Characterization of GTPase activity of TrmE, a member of a novel GTPase superfamily, from Thermotoga maritima. J. Bacteriol. 2000, 182, 7078–7082. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Pindi, P.K.; Dube, S.; Sundareswaran, V.R.; Shivaji, S. Importance of trmE for growth of the psychrophile Pseudomonas syringae at low temperatures. Appl. Environ. Microbiol. 2009, 75, 4419–4426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, S.; Ma, Z.; Foster, J.W. The Era-like GTPase TrmE conditionally activates gadE and glutamate-dependent acid resistance in Escherichia coli. Mol. Microbiol. 2004, 54, 948–961. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Shivaji, S. A cold-active heat-labile tRNA modification GTPase from a psychrophilic bacterium Pseudomonas syringae (Lz4W). Res. Microbiol. 2010, 161, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, R.B.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An old enzyme with new tricks. Trends Pharmacol. Sci. 2015, 36, 395–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGee, D.J.; Zabaleta, J.; Viator, R.J.; Testerman, T.L.; Ochoa, A.C.; Mendz, G.L. Purification and characterization of Helicobacter pylori arginase, RocF: Unique features among the arginase superfamily. Eur. J. Biochem. 2004, 271, 1952–1962. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wang, J.; Tao, H.; Xiao, Y.; Huang, S.Y. HNADOCK: A nucleic acid docking server for modeling RNA/DNA-RNA/DNA 3D complex structures. Nucleic Acids Res. 2019, 47, W35–W42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Log2 (FC) * | p Value | Description |
|---|---|---|---|
| Down-regulated in ∆Dsr11 strain | |||
| dr_0828 | −1.9173 | 1.17×10−5 | Isocitrate lyase |
| dr_2464 | −1.38547 | 4.06×10−5 | Peptidyl-prolyl cis-trans isomerase, FKBP-type |
| dr_1325 | −1.2897 | 4.02×10−5 | Endopeptidase-related protein |
| dr_0815 | −1.1394 | 4.23×10−6 | Transcriptional regulator, GntR family |
| dr_A0367 | −1.12989 | 1.3×10−4 | UDP-galactopyranose mutase |
| dr_0907† | −1.08722 | 1.73×10−5 | Cold shock protein, CSD family |
| Up-regulated in ∆Dsr11 strain | |||
| dr_0128† | 1.00099 | 1.29×10−5 | GrpE protein |
| dr_0166 | 1.03439 | 4.26×10−6 | Acyl-CoA-binding protein |
| dr_0651 | 1.10117 | 3.56×10−5 | Arginase |
| dr_1359 | 1.13075 | 3.08×10−6 | Outer membrane protein |
| dr_0139 | 1.1902 | 7.89×10−6 | GTP-binding protein HflX |
| dr_1424† | 1.23589 | 9.26×10−6 | DnaJ protein |
| dr_1046† | 1.24258 | 7.63×10−6 | ATP-dependent Clp protease, ATP-binding subunit ClpB |
| dr_2328 | 1.25488 | 4.53×10−6 | Sensor histidine kinase |
| dr_0440 | 1.30399 | 9.42×10−5 | Holliday junction resolvase |
| dr_0942 | 1.31442 | 7.5×10−4 | Tryptophan synthase, alpha subunit |
| dr_1114† | 1.51777 | 1.07×10−5 | Heat shock protein, HSP20 family |
| dr_1473 | 1.64409 | 2.04×10−5 | Phage shock protein A |
| dr_1016 | 1.70629 | 1.85×10−5 | trmE; tRNA modification GTPase TrmE |
| dr_1994 | 4.24133 | 4.23×10−5 | Hypothetical protein |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, D.; Chen, Y.; Li, J.; Han, J.; Zhou, Z.; Zhang, W.; Chen, M.; Lin, M.; Ongena, M.; Wang, J. A Novel Noncoding RNA dsr11 Involved in Heat Stress Tolerance in Deinococcus radiodurans. Biomolecules 2020, 10, 22. https://doi.org/10.3390/biom10010022
Xue D, Chen Y, Li J, Han J, Zhou Z, Zhang W, Chen M, Lin M, Ongena M, Wang J. A Novel Noncoding RNA dsr11 Involved in Heat Stress Tolerance in Deinococcus radiodurans. Biomolecules. 2020; 10(1):22. https://doi.org/10.3390/biom10010022
Chicago/Turabian StyleXue, Dong, Yun Chen, Jiang Li, Jiahui Han, Zhengfu Zhou, Wei Zhang, Ming Chen, Min Lin, Marc Ongena, and Jin Wang. 2020. "A Novel Noncoding RNA dsr11 Involved in Heat Stress Tolerance in Deinococcus radiodurans" Biomolecules 10, no. 1: 22. https://doi.org/10.3390/biom10010022
APA StyleXue, D., Chen, Y., Li, J., Han, J., Zhou, Z., Zhang, W., Chen, M., Lin, M., Ongena, M., & Wang, J. (2020). A Novel Noncoding RNA dsr11 Involved in Heat Stress Tolerance in Deinococcus radiodurans. Biomolecules, 10(1), 22. https://doi.org/10.3390/biom10010022