Elucidating the Binding Mechanism of a Novel Silica-Binding Peptide

 , , , ,
, , , , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Circular Dichroism (CD) Spectroscopy
2.3. Fluorescence Spectroscopy
2.4. Surface Plasmon Resonance (SPR)
2.5. Quartz Crystal Microbalance with Dissipation Monitoring (QCM-D)
2.6. Adsorption Isotherms
2.7. Viscoelastic Properties of Adsorbed Proteins
2.8. Chemical Biofunctionalization
3. Results and Discussion
3.1. Effect of the Linker on the Structure and Stability of Protein G
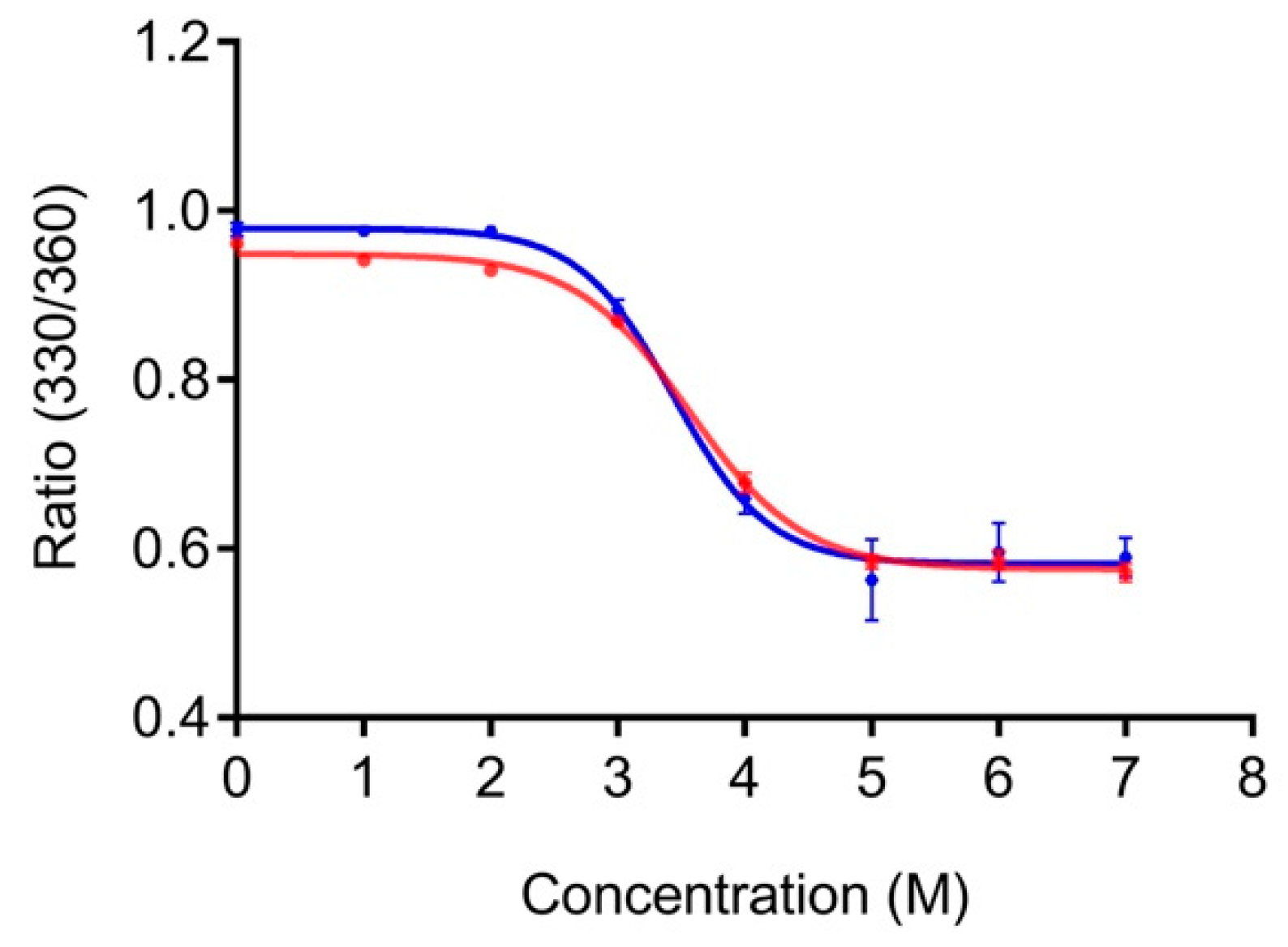
3.2. Effect of the Linker on the Antibody-Binding Function of Protein G
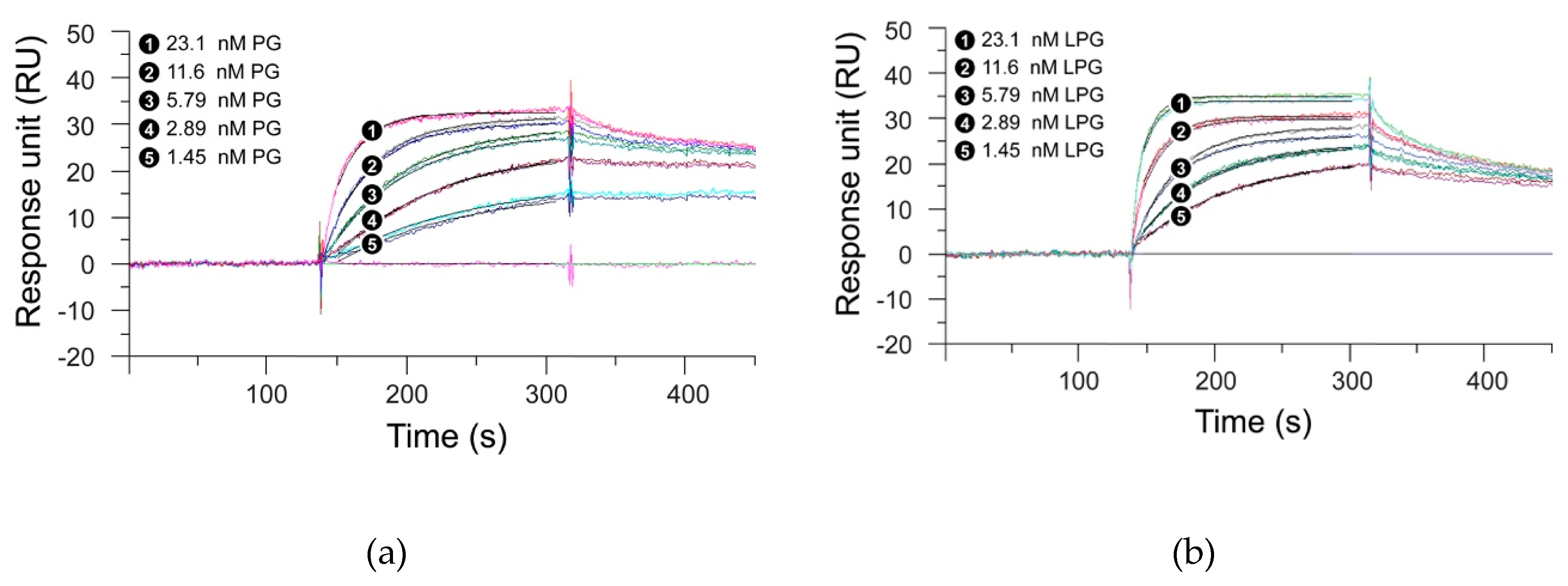
3.3. Binding Kinetics of LPG
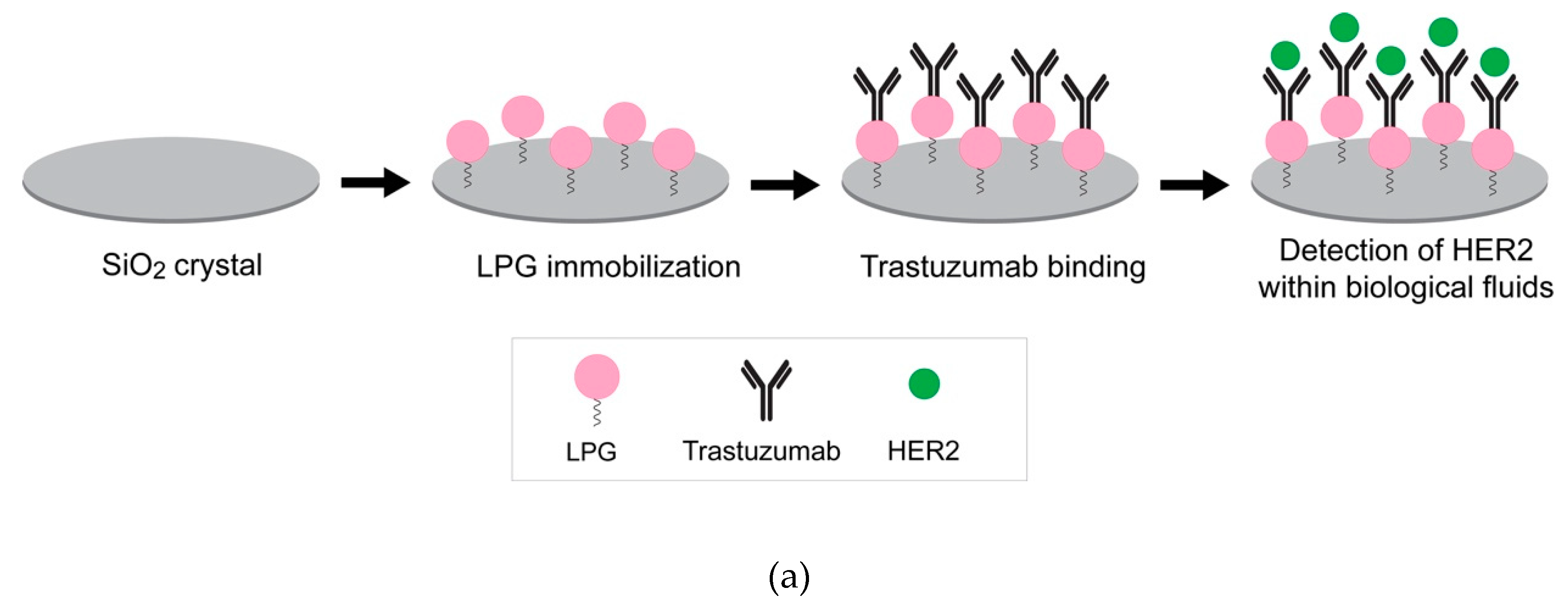
3.4. Linker and Covalent Functionalization of Silica Surface
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, G.Y.; Amro, N.A. Positioning protein molecules on surfaces: A nanoengineering approach to supramolecular chemistry. Proc. Natl. Acad. Sci. USA 2002, 99, 5165–5170. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.C.W.; Nie, S. Quantum dot bioconjugates for ultrasensitive nonisotopic detection. Science 1998, 281, 2016–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phizicky, E.; Bastiaens, P.I.; Zhu, H.; Snyder, M.; Fields, S. Protein analysis on a proteomic scale. Nature 2003, 422, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, H.; Sekine, J.; Yajima, H.; Serizawa, T. Biological selection of peptides for poly (l-lactide) substrates. Langmuir 2008, 24, 6399–6403. [Google Scholar] [CrossRef] [PubMed]
- Hnilova, M.; Oren, E.E.; Seker, U.O.S.; Wilson, B.R.; Collino, S.; Evans, J.S. Effect of molecular conformations on the adsorption behavior of gold-binding peptides. Langmuir 2008, 24, 12440–12445. [Google Scholar] [CrossRef] [PubMed]
- Serizawa, T.; Sawada, T.; Matsuno, H. Highly specific affinities of short peptides against synthetic polymers. Langmuir 2007, 23, 11127–11133. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Care, A.; Lord, M.S.; Walsh, T.R.; Sunna, A. Experimental and theoretical tools to elucidate the binding mechanisms of solid-binding peptides. N. Biotechnol. 2019, 52, 9–18. [Google Scholar] [CrossRef]
- Naik, R.R.; Stringer, S.J.; Agarwal, G.; Jones, S.E.; Stone, M.O. Biomimetic synthesis and patterning of silver nanoparticles. Nat. Mater. 2002, 1, 169–172. [Google Scholar] [CrossRef]
- Nel, A.E.; Madler, L.; Velegol, D.; Xia, T.; Hoek, E.M.V.; Somasundaran, P. Understanding biophysicochemical interactions at the nano-bio interface. Nat. Mater. 2009, 8, 543–557. [Google Scholar] [CrossRef]
- Seker, U.O.S.; Wilson, B.; Dincer, S.; Kim, I.W.; Oren, E.E.; Evans, J.S.; Tamerler, C.; Sarikaya, M. Adsorption behaviour of linear and cyclic genetically engineered platinum binding peptides. Langmuir 2007, 23, 7895–7900. [Google Scholar] [CrossRef]
- Sultan, A.M.; Westcott, Z.C.; Hughes, Z.E.; Palafox-Hernandez, J.P.; Giesa, T.; Puddu, V. Aqueous peptide–TiO2 interfaces: Isoenergetic binding via either entropically or enthalpically driven mechanisms. ACS Appl. Mater. Interface 2016, 8, 18620–18630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, T.; Okeya, Y.; Hashizume, M.; Serizawa, T. Screening of peptides recognizing simple polycyclic aromatic hydrocarbons. Chem. Commun. (Camb. U.K.) 2013, 49, 5088–5090. [Google Scholar] [CrossRef] [PubMed]
- Sunna, A.; Chi, F.; Bergquist, P.L. A linker peptide with high affinity towards silica-containing materials. N. Biotechnol. 2013, 30, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Care, A.; Zhang, R.; Lu, Y.; Packer, N.H.; Sunna, A.; Qian, Y.; Zvyagin, A.V. Facile assembly of functional upconversion nanoparticles for targeted cancer imaging and photodynamic therapy. ACS Appl. Mater. Interfaces 2016, 8, 11945–11953. [Google Scholar] [CrossRef] [PubMed]
- Care, A.; Petroll, K.; Gibson, E.S.Y.; Bergquist, P.L.; Sunna, A. Solid-binding peptides for immobilisation of thermostable enzymes to hydrolyse biomass polysaccharides. Biotechnol. Biofuels 2017, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayyadi, N.; Care, A.; Connally, R.E.; Try, A.C.; Bergquist, P.L.; Sunna, A. A novel universal detection agent for time-gated luminescence bioimaging. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shipunova, V.O.; Zelepukin, I.V.; Stremovskiy, O.A.; Nikitin, M.P.; Care, A.; Sunna, A.; Zvyagin, A.V.; Deyev, S.M. Versatile platform for nanoparticle surface bioengineering based on SiO2-binding peptide and proteinaceous Barnase*Barstar interface. ACS Appl. Mater. Interfaces 2018, 10, 17437–17447. [Google Scholar] [CrossRef] [PubMed]
- Voinova, M.B.; Jonson, M.; Kasemo, B. ‘Missing mass’ effect in biosensor’s QCM applications. Biosens. Bioelectron. 2002, 17, 835–841. [Google Scholar] [CrossRef]
- Lord, M.S.; Whitelock, J.M.; Simmons, A.; Williams, R.L.; Milthorpe, B.K. Fibrinogen adsorption and platelet adhesion to silica surfaces with stochastic nanotopography. Biointerphases 2014, 9, 041002. [Google Scholar] [CrossRef]
- Liu, S.X.; Kim, J.T. Application of Kevin-Voigt model in quantifying whey protein adsorption on Polyethersulfone using QCM-D. JALA 2009, 14, 213–220. [Google Scholar] [CrossRef]
- Hovgaard, M.B.; Dong, M.; Otzen, D.E.; Besenbacher, F. Quartz Crystal Microbalance studies of multilayer glucagon fibrillation at the solid-liquid interface. Biophys. J. 2007, 93, 2162–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmore, L.; Wallace, B.A. DICHROWEB: An online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, W668–W673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreerema, N.; Venyaminov, S.Y.; Woody, R.W. Estimation of the number of helical and strand segments in proteins using CD spectroscopy. Protein Sci. 1999, 8, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from CD spectra: Comparison of CONTIN, SELCON and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Venyaminov, S.Y.; Woody, R.W. Estimation of protein secondary structure from CD spectra: Inclusion of denatured proteins with native protein in the analysis. Anal. Biochem. 2000, 287, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Goward, C.R.; Irons, L.I.; Murphy, J.P.; Atkinson, T. The secondary structure of Protein G’, a robust molecule. Biochem. J. 1991, 274, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Khrapunov, S. Circular dichroism spectroscopy has intrinsic limitations for protein secondary structure analysis. Anal. Biochem. 2009, 389, 174–176. [Google Scholar] [CrossRef] [Green Version]
- Atkins, P.W. Physikalische Chemie; Wiley-VCH: Weinheim, Germany, 2001. [Google Scholar]
- Seker, U.O.S.; Wilson, B.; Sahin, D.; Tamerler, C.; Sarikaya, M. Quantitative affinity of genetically engineered repeating polypeptides to inorganic surfaces. Biomacromolecules 2009, 10, 250–257. [Google Scholar] [CrossRef]
- Hnilova, M.; So, C.R.; Oren, E.E.; Wilson, B.R.; Kacar, T.; Tamerler, C.; Sarikaya, M. Peptide-directed co-assembly of nanoprobes on multimaterial patterned solid surfaces. Soft Matter 2012, 8, 4327–4334. [Google Scholar] [CrossRef] [Green Version]
- Seker, U.O.S.; Sharma, V.K.; Akhavan, S.; Demir, H.V. Engineered peptides for nanohybrid assemblies. Langmuir 2014, 30, 2137–2143. [Google Scholar] [CrossRef] [Green Version]
- Marx, K.A. Quartz crystal microbalance: A useful tool for studying thin polymer films and comolex biomolecular systems at the solution-surface interface. Biomacromolecules 2003, 4, 1099–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Marx, K.A.; Warren, M.; Schulze, H.; Braunhut, S.J. The quartz crystal microbalance as a continuous monitoring tool for the study of endothelial cell surface attachment and growth. Biotechnol. Prog. 2000, 16, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.C.; Green, R.A.; Boscher, F.; Poole-Warren, L.A.; Carter, P.M.; Enke, Y.L.; Lovell, N.H.; Lord, M.S. Development and performance of a biomimetic artificial perilymph for in vitro testing of medical devices. J. Neural Eng. 2019, 16, 026006. [Google Scholar] [CrossRef] [PubMed]
- Puddu, V.; Perry, C.C. Peptide adsorption on silica nanoparticles: Evidence of hydrophobic interactions. ACS Nano 2012, 6, 6356–6363. [Google Scholar] [CrossRef]
- Höök, F.; Vörös, J.; Rodahl, M.; Kurrat, R.; Böni, P.; Ramsden, J.J.; Textor, M.; Spencer, N.D.; Tengvall, P.; Gold, J.; et al. A comparative study of protein adsorption on titanium oxide surfaces using in situ ellipsometry, optical waveguide lightmode spectroscopy, and quartz crystal microbalance/dissipation. Colloids Surf. B Biointerfaces 2002, 24, 155–170. [Google Scholar] [CrossRef]
- Heger, J.I.; Froehlich, K.; Pastuschek, J.; Schmidt, A.; Baer, C.; Mrowka, R.; Backsh, C.; Schleußner, E.; Markert, U.R.; Schmidt, A. Human serum alters cell culture behavior and improves spheroid formation in comparison to fetal bovine serum. Exp. Cell Res. 2018, 365, 57–65. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein-Antibody Complex | Rate of Association ka (M−1s−1) | Rate of Dissociation kd (s−1) | Equilibrium Dissociation Constant KD (M) | Half-Life t1/2 (s) |
|---|---|---|---|---|
| PG + trastuzumab | 1.79 ± 0.10 × 106 | 8.24 ± 0.92 × 10−3 | 4.62 ± 0.79 × 10−9 | 83.7 |
| LPG + trastuzumab | 3.88 ± 1.20 × 106 | 7.48 ± 0.38 × 10−3 | 2.07 ± 0.71 × 10−9 | 92.2 |
| PG + G203 (IgG1) | 0.84 ± 0.02 × 106 | 5.32 ± 0.29 × 10−3 | 6.30 ± 0.32 × 10−9 | 129.7 |
| LPG + G203 (IgG1) | 1.23 ± 0.06 × 106 | 9.50 ± 0.30 × 10−3 | 7.77 ± 0.58 × 10−9 | 72.6 |
| Thickness (@Req)a nm | Thickness (@kd)b nm | Mass Deposited (@Req) ng/cm2 | Mass Deposited (@kd ) ng/cm2 | Viscosity (@Req) × 10−4 kg/ms | Viscosity (@kd) × 10−4 kg/ms | |
|---|---|---|---|---|---|---|
| LPG/SiO2 | 7.84 ± 0.22 | 5.59 ± 0.22 | 862.95 ± 42.43 | 615.13 ± 58.06 | 24.70 ± 0.50 | 20.57 ± 0.45 |
| Immobilization | Thickness of Bound Protein (nm) | Mass of Protein Deposited (ng/cm2) | Thickness of Bound Antibody (nm) | Mass Deposited for Bound Antibody (ng/cm2) | Thickness of Bound Antigen (nm) | Mass Deposited for Bound Antigen (ng/cm2) |
|---|---|---|---|---|---|---|
| LPG physisorbed | 7.84 ± 0.22 | 862.95 | 20 ± 0.05 | 1200.14 | 27.84 ± 0.74 | 1600.47 |
| PG chemically immobilized | 2.89 ± 0.18 | 256.76 | 7.43 ± 0.15 | 450.79 | 12.76 ± 0.22 | 578.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bansal, R.; Elgundi, Z.; Care, A.; C. Goodchild, S.; S. Lord, M.; Rodger, A.; Sunna, A. Elucidating the Binding Mechanism of a Novel Silica-Binding Peptide. Biomolecules 2020, 10, 4. https://doi.org/10.3390/biom10010004
Bansal R, Elgundi Z, Care A, C. Goodchild S, S. Lord M, Rodger A, Sunna A. Elucidating the Binding Mechanism of a Novel Silica-Binding Peptide. Biomolecules. 2020; 10(1):4. https://doi.org/10.3390/biom10010004
Chicago/Turabian StyleBansal, Rachit, Zehra Elgundi, Andrew Care, Sophia C. Goodchild, Megan S. Lord, Alison Rodger, and Anwar Sunna. 2020. "Elucidating the Binding Mechanism of a Novel Silica-Binding Peptide" Biomolecules 10, no. 1: 4. https://doi.org/10.3390/biom10010004
APA StyleBansal, R., Elgundi, Z., Care, A., C. Goodchild, S., S. Lord, M., Rodger, A., & Sunna, A. (2020). Elucidating the Binding Mechanism of a Novel Silica-Binding Peptide. Biomolecules, 10(1), 4. https://doi.org/10.3390/biom10010004