Peptide Linker Affecting the Activity Retention Rate of VHH in Immunosorbents


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Expression of the Anti-β2MG VHHs
2.3. Purification of the Recombinant Anti-β2MG VHHs
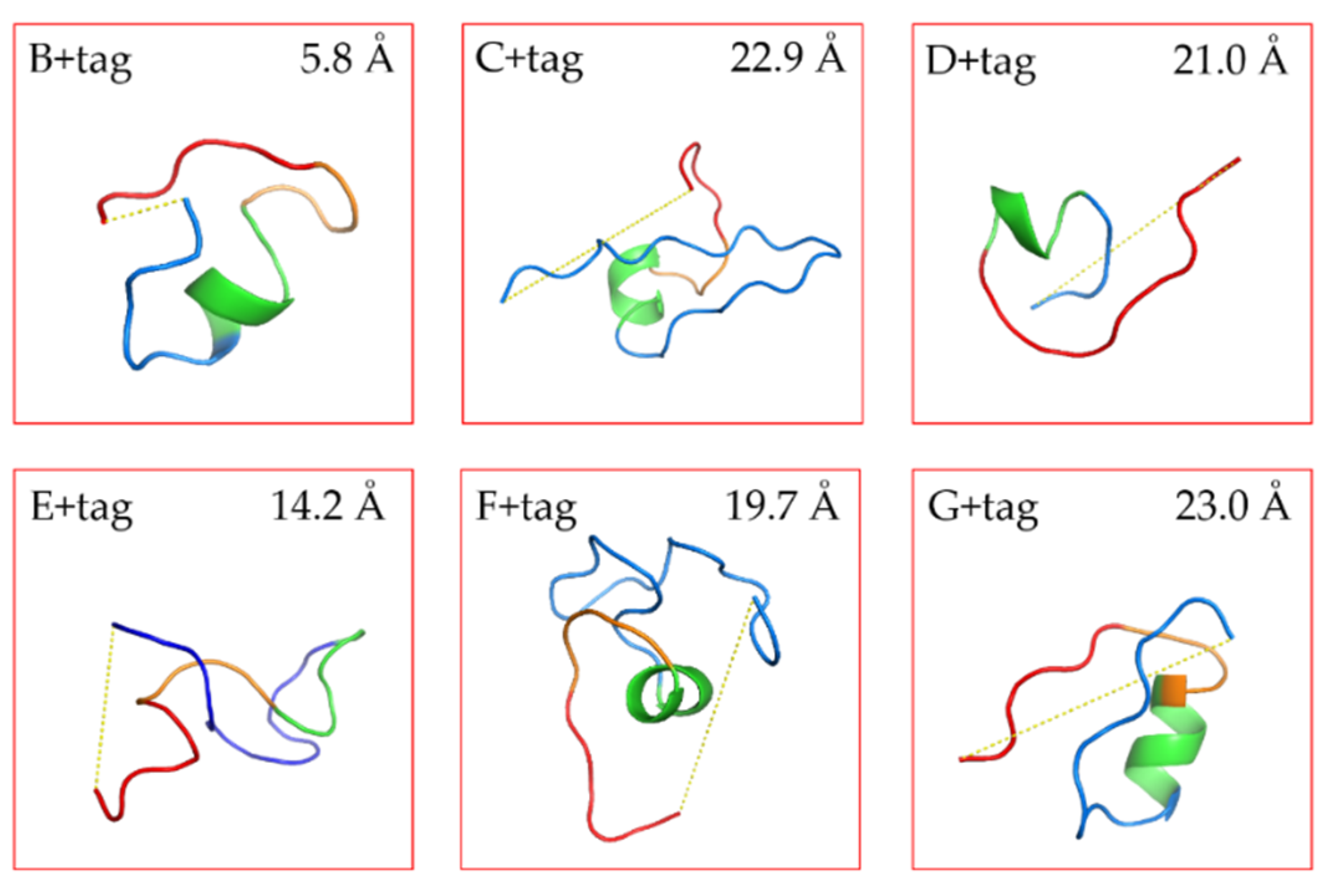
2.4. De Novo Structure Prediction for Linkers
2.5. Anti-β2MG VHHs Affinity Measurement
2.6. Generation of the Aldehyde Group
2.7. Preparation of VHH Based Immunosorbents
2.8. Adsorption Performance of the VHH-Based Immunosorbents
3. Results and Discussions
3.1. Expression and Purification of Anti-β2MG VHH with Different Linkers
3.2. De Novo Structure Prediction for Linkers
3.3. Anti-β2MG VHH Affinity Measurements
3.4. fGly Conversion
3.5. Immobilization of VHH-Aldehyde
3.6. Adsorption Performance of the VHH-Based Immunosorbents
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Subramanian, A. Immunoaffinity chromatography. Mol. Biotechnol. 2002, 20, 41–47. [Google Scholar] [CrossRef]
- Ingavle, G.C.; Baillie, L.; Zheng, Y.; Lis, E.K.; Savina, I.N.; Howel, C.A.; Mikhalovsky, S.; Sandeman, S. Affinity binding of antibodies to supermacroporous cryogel adsorbents with immobilized protein A for removal of anthrax toxin protective antigen. Biomaterials 2015, 50, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Weber, V.; Linsberger, I.; Ettenauer, M.; Loth, F.; Höyhtyä, M.; Falkenhagen, D. Development of specific adsorbents for human tumor necrosis factor-alpha: Influence of antibody immobilization on performance and biocompatibility. Biomacromolecules 2005, 6, 1864–1870. [Google Scholar] [CrossRef]
- Vallar, L.; Costa, P.M.; Teixeira, A.; Pfister, M.; Barrois, R.; Costa, P.P.; Rivat, C. Immunoadsorption Procedure as a Potential Method for the Specific Beta-2-Microglobulin Removal from Plasma of Patients with Chronic-Renal-Failure. J. Chromatogr. B Biomed. Appl. 1995, 664, 97–106. [Google Scholar] [CrossRef]
- Ameer, G.A.; Grovender, E.A.; Ploegh, H.; Ting, D.; Owen, W.F.; Rupnick, M.; Langer, R. A novel immunoadsorption device for removing beta(2)-microglobulin from whole blood. Kidney Int. 2001, 59, 1544–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grovender, E.A.; Kellogg, B.; Singh, J.; Blom, D.; Ploegh, H.; Wittrup, K.D.; Langer, R.S.; Ameer, G.A. Single-chain antibody fragment-based adsorbent for the extracorporeal removal of beta(2)-microglobulin. Kidney Int. 2004, 65, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zang, B.; Huang, C.; Ren, J.; Jia, L. One-step Preparation of a VHH-based Immunoadsorbent for the Extracorporeal Removal of beta2-microglobulin. Molecules 2019, 24, 2119. [Google Scholar] [CrossRef] [Green Version]
- Appel, M.J.; Bertozzi, C.R. Formylglycine, a Post-Translationally Generated Residue with Unique Catalytic Capabilities and Biotechnology Applications. ACS Chem. Biol. 2014, 10, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Krüger, T.; Dierks, T.; Sewald, N. Formylglycine-generating enzymes for site-specific bioconjugation. Biol. Chem. 2019, 400, 289–297. [Google Scholar] [CrossRef]
- Dierks, T.; Lecca, M.; Schlotterhose, P.; Schmidt, B.; Von Figura, K. Sequence determinants directing conversion of cysteine to formylglycine in eukaryotic sulfatases. EMBO J. 1999, 18, 2084–2091. [Google Scholar] [CrossRef] [Green Version]
- Drake, P.M.; Albers, A.E.; Baker, J.; Banas, S.; Barfield, R.M.; Bhat, A.S.; De Hart, G.W.; Garofalo, A.W.; Holder, P.; Jones, L.C.; et al. Aldehyde Tag Coupled with HIPS Chemistry Enables the Production of ADCs Conjugated Site-Specifically to Different Antibody Regions with Distinct in Vivo Efficacy and PK Outcomes. Bioconjug. Chem. 2014, 25, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zheng, J.; Xu, R.; Barany, A.G.; Cowburn, D. Flexibility of Interdomain Contacts Revealed by Topological Isomers of Bivalent Consolidated Ligands to the Dual Src Homology Domain SH(32) of Abelson. Biochemistry 1999, 38, 3491–3497. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, R.S. Role of linkers in communication between protein modules. Curr. Opin. Chem. Biol. 2000, 4, 22–27. [Google Scholar] [CrossRef]
- Silacci, M.; Baenziger-Tobler, N.; Lembke, W.; Zha, W.; Batey, S.; Bertschinger, J.; Grabulovski, D. Linker Length Matters, Fynomer-Fc Fusion with an Optimized Linker Displaying Picomolar IL-17A Inhibition Potency. J. Biol. Chem. 2014, 289, 14392–14398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Zaro, J.L.; Shen, W.-C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- George, R.A.; Heringa, J. An analysis of protein domain linkers: Their classification and role in protein folding. Protein Eng. Des. Sel. 2002, 15, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Evers, T.H.; Van Dongen, E.M.W.M.; Faesen, A.C.; Meijer, E.B.; Merkx, M. Quantitative Understanding of the Energy Transfer between Fluorescent Proteins Connected via Flexible Peptide Linkers. Biochemistry 2006, 45, 13183–13192. [Google Scholar] [CrossRef] [Green Version]
- Schuler, B.; Lipman, E.A.; Steinbach, P.J.; Kumke, M.; Eaton, W.A. Polyproline and the “spectroscopic ruler” revisited with single-molecule fluorescence. Proc. Natl. Acad. Sci. USA 2005, 102, 2754–2759. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Ji, F.; Huang, C.; Jia, L. High Expression Achievement of Active and Robust Anti-beta 2 microglobulin Nanobodies via E.coli Hosts Selection. Molecules 2019, 24, 2860. [Google Scholar] [CrossRef] [Green Version]
- Lamiable, A.; Thévenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tufféry, P. PEP-FOLD3: Faster denovo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Zang, B.; Zhao, W.; Li, D.; Ren, J.; Ji, F.; Jia, L. Efficient continuous-flow aldehyde tag conversion using immobilized formylglycine generating enzyme. Catal. Sci. Technol. 2020, 10, 484–492. [Google Scholar] [CrossRef]
- Knop, M.; Dang, T.Q.Q.; Jeschke, G.; Seebeck, F.P. Copper is a Cofactor of the Formylglycine-Generating Enzyme. ChemBioChem 2017, 18, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Holder, P.G.; Jones, L.C.; Drake, P.M.; Barfield, R.M.; Bañas, S.; De Hart, G.W.; Baker, J.; Rabuka, D. Reconstitution of Formylglycine-generating Enzyme with Copper(II) for Aldehyde Tag Conversion. J. Biol. Chem. 2015, 290, 15730–15745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knop, M.; Engi, P.; Lemnaru, R.; Seebeck, F.P. In Vitro Reconstitution of Formylglycine-Generating Enzymes Requires Copper(I). ChemBioChem 2015, 16, 2147–2150. [Google Scholar] [CrossRef] [Green Version]
- Shamriz, S.; Ofoghi, H.; Moazami, N. Effect of linker length and residues on the structure and stability of a fusion protein with malaria vaccine application. Comput. Biol. Med. 2016, 76, 24–29. [Google Scholar] [CrossRef]
- Sabourin, M.; Tuzon, C.T.; Fisher, T.S.; Zakian, V.A. A flexible protein linker improves the function of epitope-tagged proteins in Saccharomyces cerevisiae. Yeast 2007, 24, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Gray, N.K.; Hentze, M.W. Regulation of protein synthesis by mRNA structure. Mol. Biol. Rep. 1994, 19, 195–200. [Google Scholar] [CrossRef]
- Gutman, G.A.; Hatfield, G.W. Nonrandom utilization of codon pairs in Escherichia coli. Proc. Natl. Acad. Sci. USA 1989, 86, 3699–3703. [Google Scholar] [CrossRef] [Green Version]
- Muyldermans, S. Nanobodies: Natural Single-Domain Antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Huang, H. Expression of single-domain antibody in different systems. Appl. Microbiol. Biotechnol. 2018, 102, 539–551. [Google Scholar] [CrossRef]
- Klement, M.; Liu, C.; Loo, B.L.W.; Choo, A.B.-H.; Ow, D.S.-W.; Lee, D.-Y. Effect of linker flexibility and length on the functionality of a cytotoxic engineered antibody fragment. J. Biotechnol. 2015, 199, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Huang, Z.; Zhang, C.; Dong, B.-J.; Guo, R.-H.; Yue, H.-W.; Yan, L.-T.; Xing, X.-H. Construction of a linker library with widely controllable flexibility for fusion protein design. Appl. Microbiol. Biotechnol. 2015, 100, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Cetinkaya, M.; Gräter, F. How Sequence Determines Elasticity of Disordered Proteins. Biophys. J. 2010, 99, 3863–3869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lith, S.A.M.; Van Duijnhoven, S.M.J.; Navis, A.C.; Leenders, W.P.; Dolk, E.; Wennink, J.W.H.; Van Nostrum, C.F.; Van Hest, J.C.M. Legomedicine—A Versatile Chemo-Enzymatic Approach for the Preparation of Targeted Dual-Labeled Llama Antibody—Nanoparticle Conjugates. Bioconjug. Chem. 2017, 28, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Vosjan, M.J.; Vercammen, J.; Kolkman, J.A.; Walsum, M.S.-V.; Revets, H.; Van Dongen, G.A.M.S. Nanobodies Targeting the Hepatocyte Growth Factor: Potential New Drugs for Molecular Cancer Therapy. Mol. Cancer Ther. 2012, 11, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, R.; Korfmacher, W. The emergence of high-resolution MS as the premier analytical tool in the pharmaceutical bioanalysis arena. Bioanalysis 2012, 4, 467–469. [Google Scholar] [CrossRef]
- Sun, J.; Chen, P. Ultra High-Performance Liquid Chromatography with High-Resolution Mass Spectrometry Analysis of African Mango (Irvingia gabonensis) Seeds, Extract, and Related Dietary Supplements. J. Agric. Food Chem. 2012, 60, 8703–8709. [Google Scholar] [CrossRef] [Green Version]
- Kumada, Y.; Kawasaki, T.; Kikuchi, Y.; Katoh, S. Polypeptide linkers suitable for the efficient production of dimeric scFv in Escherichia coli. Biochem. Eng. J. 2007, 35, 158–165. [Google Scholar] [CrossRef]
- Huang, C.; Ren, J.; Ji, F.; Muyldermans, S.; Jia, L. Nanobody-Based high-performance immunosorbent for selective beta 2-microglobulin purification from blood. Acta Biomater. 2020, 107, 232–241. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VHH | Linker | Yield (mg/L) | Mass (Da) | Expression Level in Molar % Relative to No Linker (100%) |
|---|---|---|---|---|
| CNb1-A | - | 197 ± 4 | 15,975.03 | 100 |
| CNb1-B | AHHSEDP | 191 ± 6 | 16,784.98 | 92 |
| CNb1-C | EPKTPKPQPQPQPQPQPNPTTE | 157 ± 3 | 18,421.83 | 69 |
| CNb1-D | GGGGS | 161 ± 2 | 16,290.61 | 81 |
| CNb1-E | GGGGSGGGGSGGGGS | 95 ± 5 | 16,956.39 | 45 |
| CNb1-F | AHHSEDPGGGGSGGGGSGGGGS | 208 ± 4 | 17,730.84 | 95 |
| CNb1-G | STPPTPSPSTPP | 160 ± 2 | 17,121.40 | 76 |
| VHH | qm (mg/g Resin) | Kd (M) | Activity Retention (%) |
|---|---|---|---|
| CNb1-A | 5.68 ± 0.02 | (2.14 ± 0.19) × 10−5 | 49.87 |
| CNb1-B | 4.83 ± 0.03 | (7.51 ± 0.13) × 10−6 | 47.74 |
| CNb1-C | 2.89 ± 0.11 | (9.61 ± 1.22) × 10−6 | 71.39 |
| CNb1-D | 2.49 ± 0.02 | (9.18 ± 0.40) × 10−6 | 49.11 |
| CNb1-G | 2.48 ± 0.02 | (7.81 ± 0.32) × 10−6 | 68.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Ren, J.; Ji, F.; Peng, Q.; Teng, H.; Jia, L. Peptide Linker Affecting the Activity Retention Rate of VHH in Immunosorbents. Biomolecules 2020, 10, 1610. https://doi.org/10.3390/biom10121610
Li D, Ren J, Ji F, Peng Q, Teng H, Jia L. Peptide Linker Affecting the Activity Retention Rate of VHH in Immunosorbents. Biomolecules. 2020; 10(12):1610. https://doi.org/10.3390/biom10121610
Chicago/Turabian StyleLi, Da, Jun Ren, Fangling Ji, Qiang Peng, Hu Teng, and Lingyun Jia. 2020. "Peptide Linker Affecting the Activity Retention Rate of VHH in Immunosorbents" Biomolecules 10, no. 12: 1610. https://doi.org/10.3390/biom10121610
APA StyleLi, D., Ren, J., Ji, F., Peng, Q., Teng, H., & Jia, L. (2020). Peptide Linker Affecting the Activity Retention Rate of VHH in Immunosorbents. Biomolecules, 10(12), 1610. https://doi.org/10.3390/biom10121610