Structural Insight into the Mechanism of N-Linked Glycosylation by Oligosaccharyltransferase




Abstract
:1. Introduction
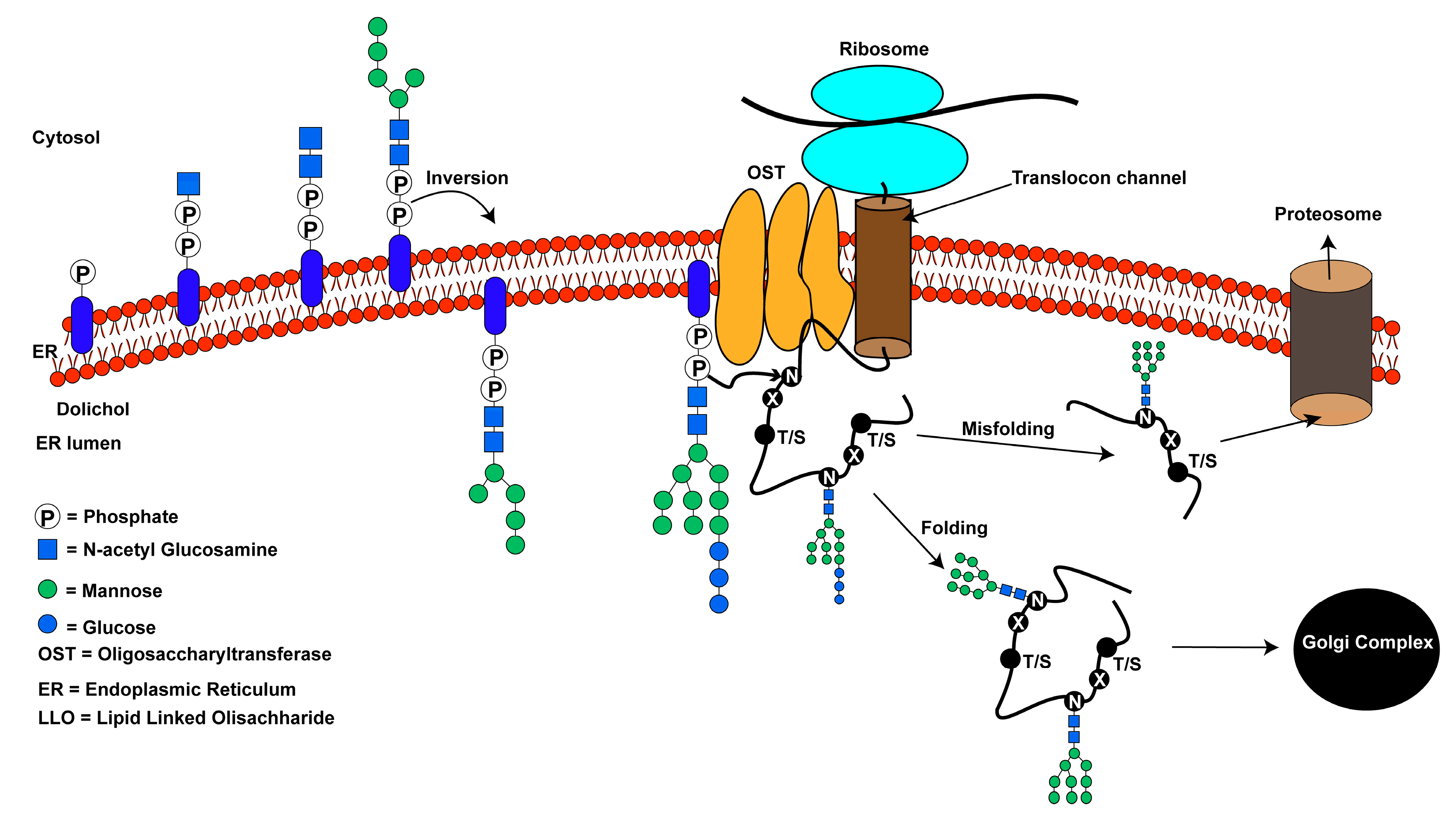
2. N-Linked Glycosylation: An Overview
2.1. Donor Substrates in Prokaryotes and Archaea
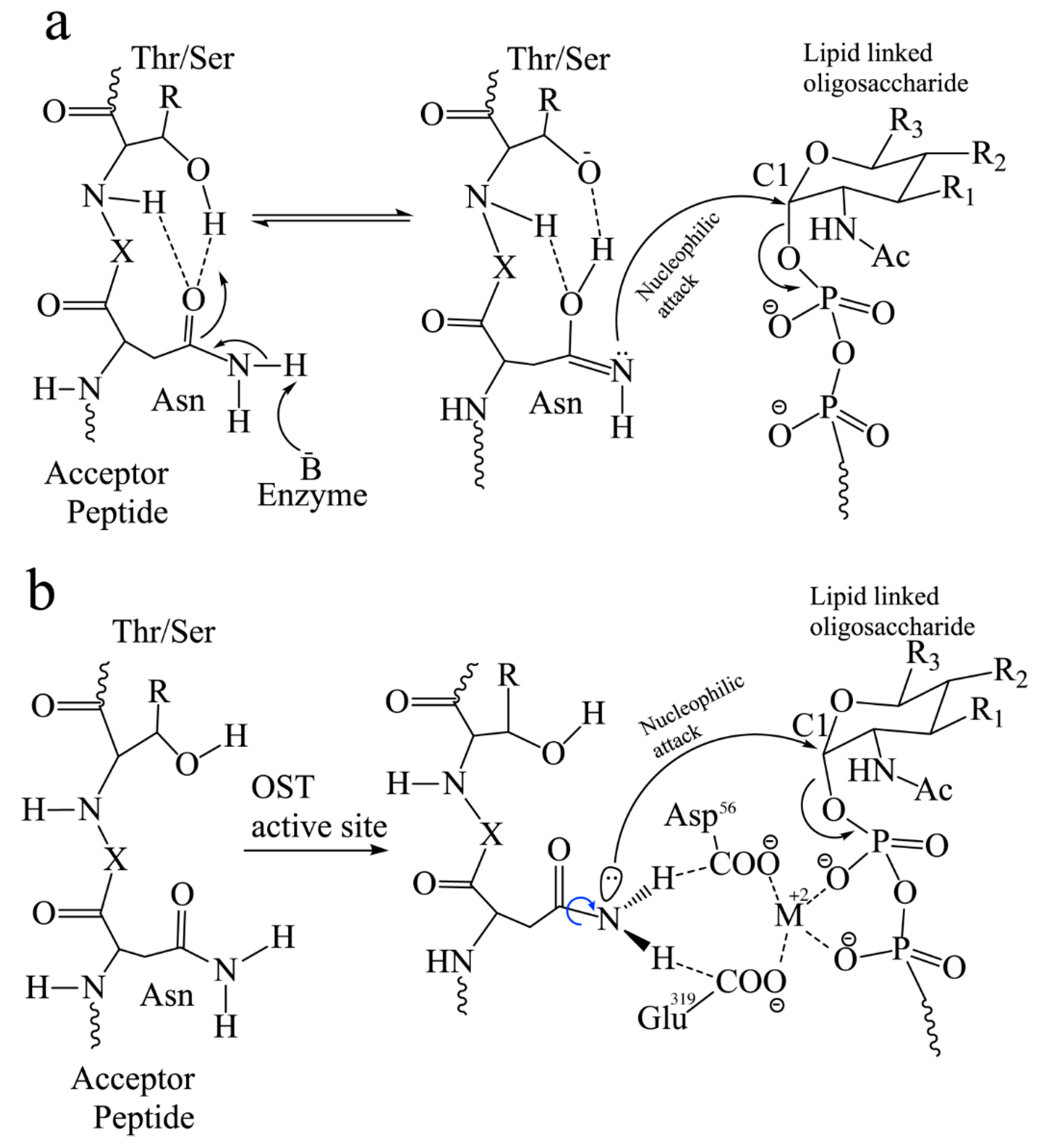
2.2. Donor Substrate in Eukaryotes and Possible Mechanism of Sugar Transfer
3. Structural Mechanisms of N-Linked Glycosylation by OST
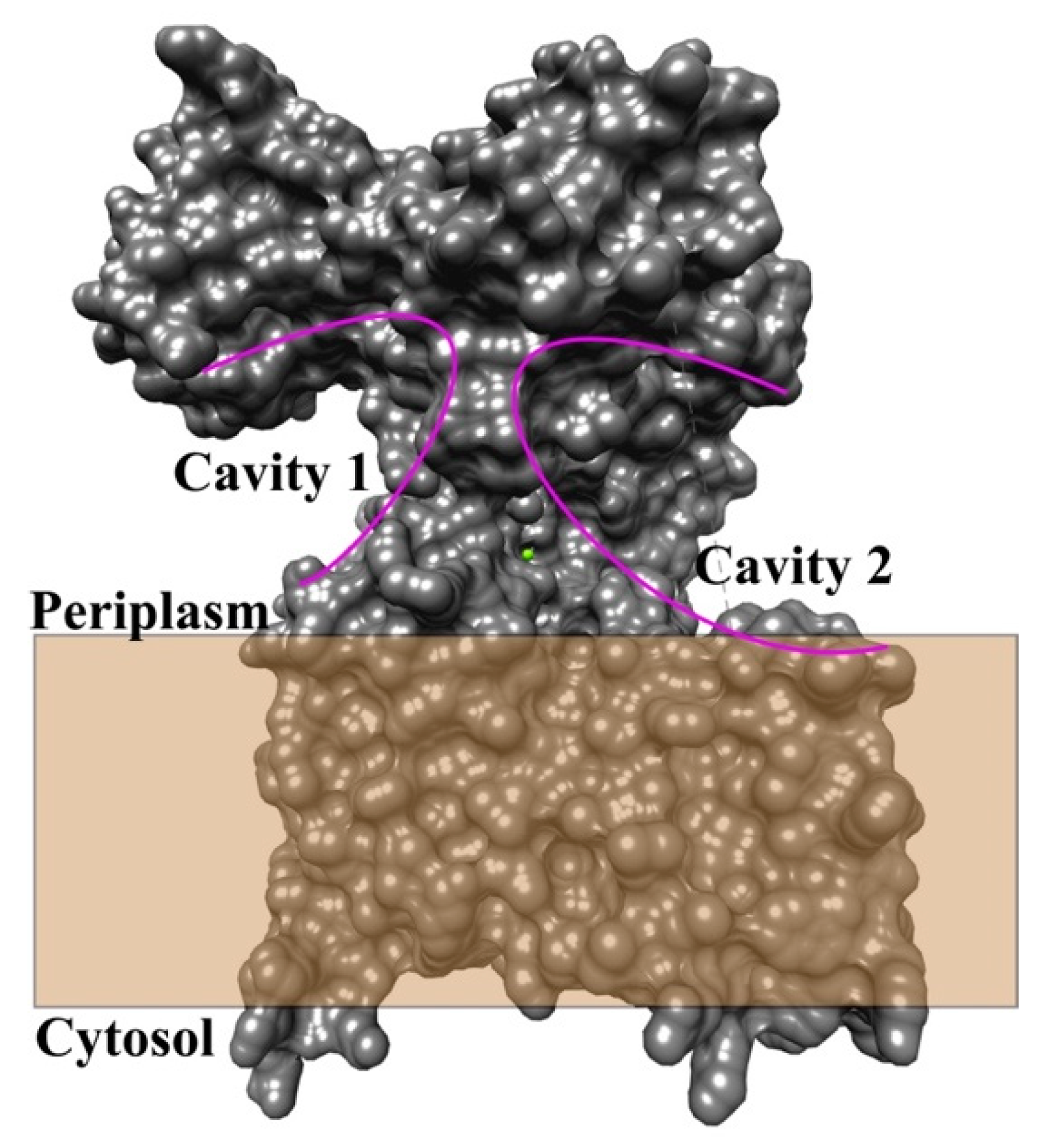
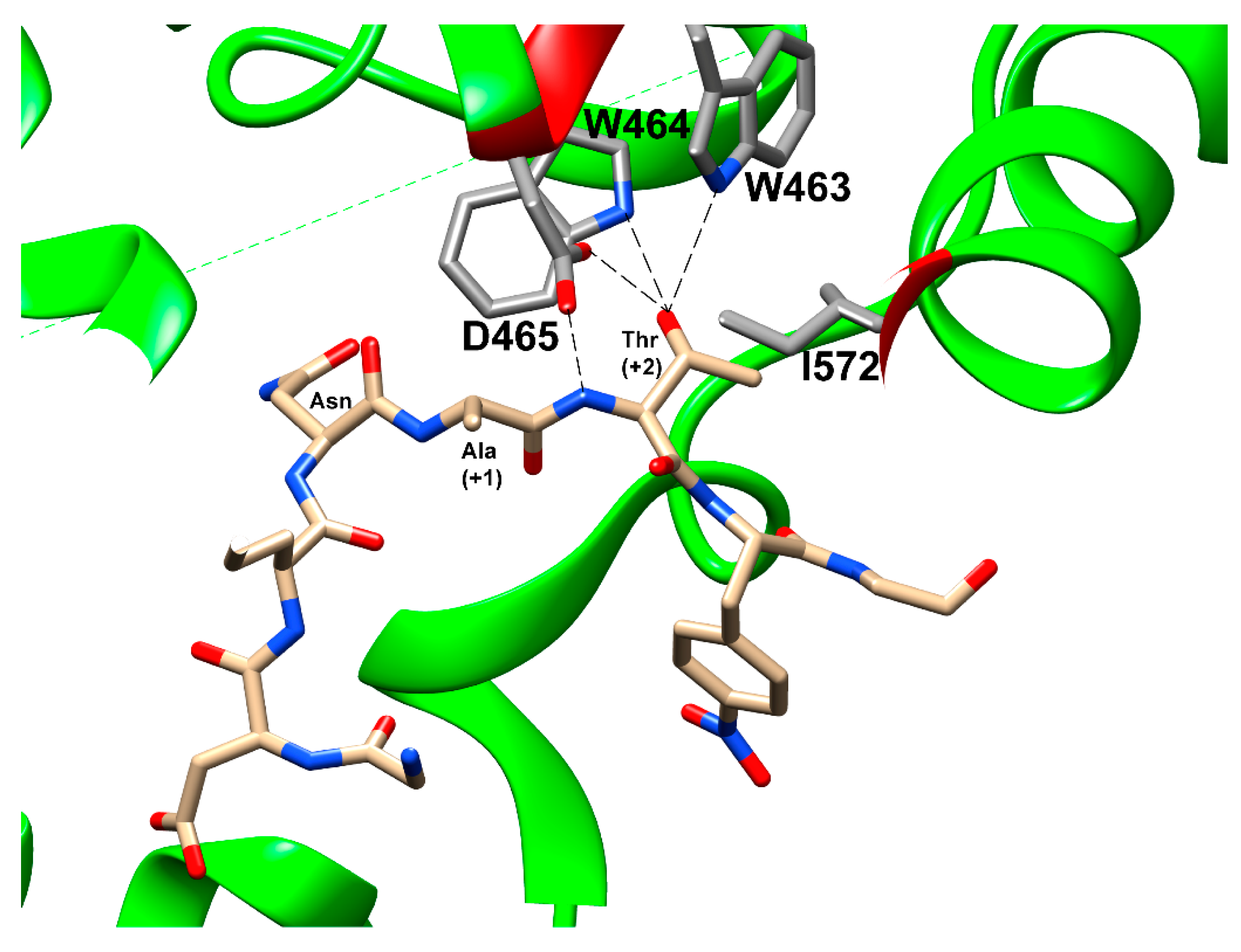
3.1. Bacteria and Archaea
3.2. Yeast
3.2.1. Catalytic Subunit Stt3
3.2.2. Non-Catalytic Subunits
3.2.3. Pathway for LLO Entry in Yeast OST
3.2.4. OST-Translocon Interaction
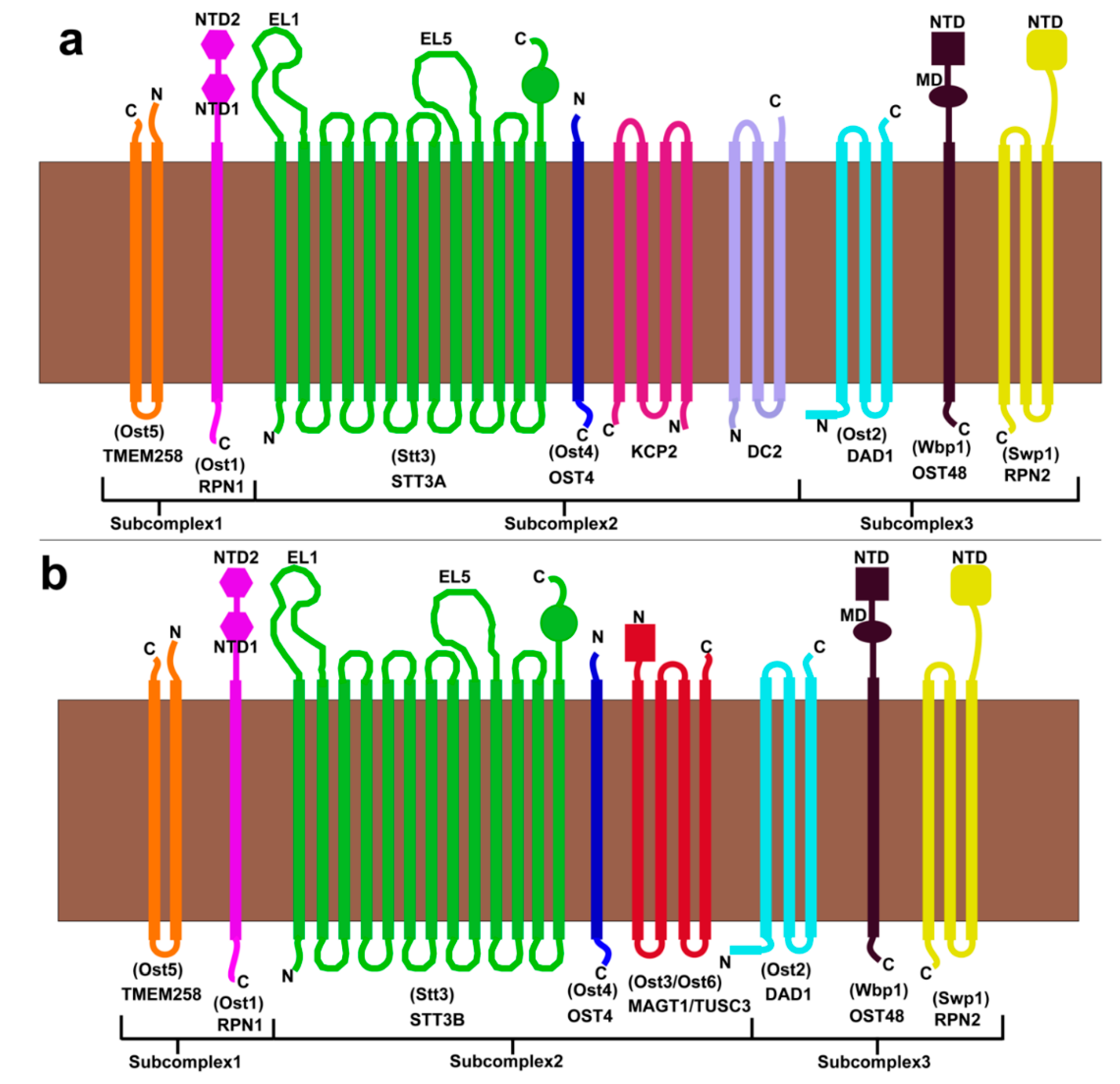
3.2.5. Assembly of Subcomplexes in the OST Complex
3.3. Human
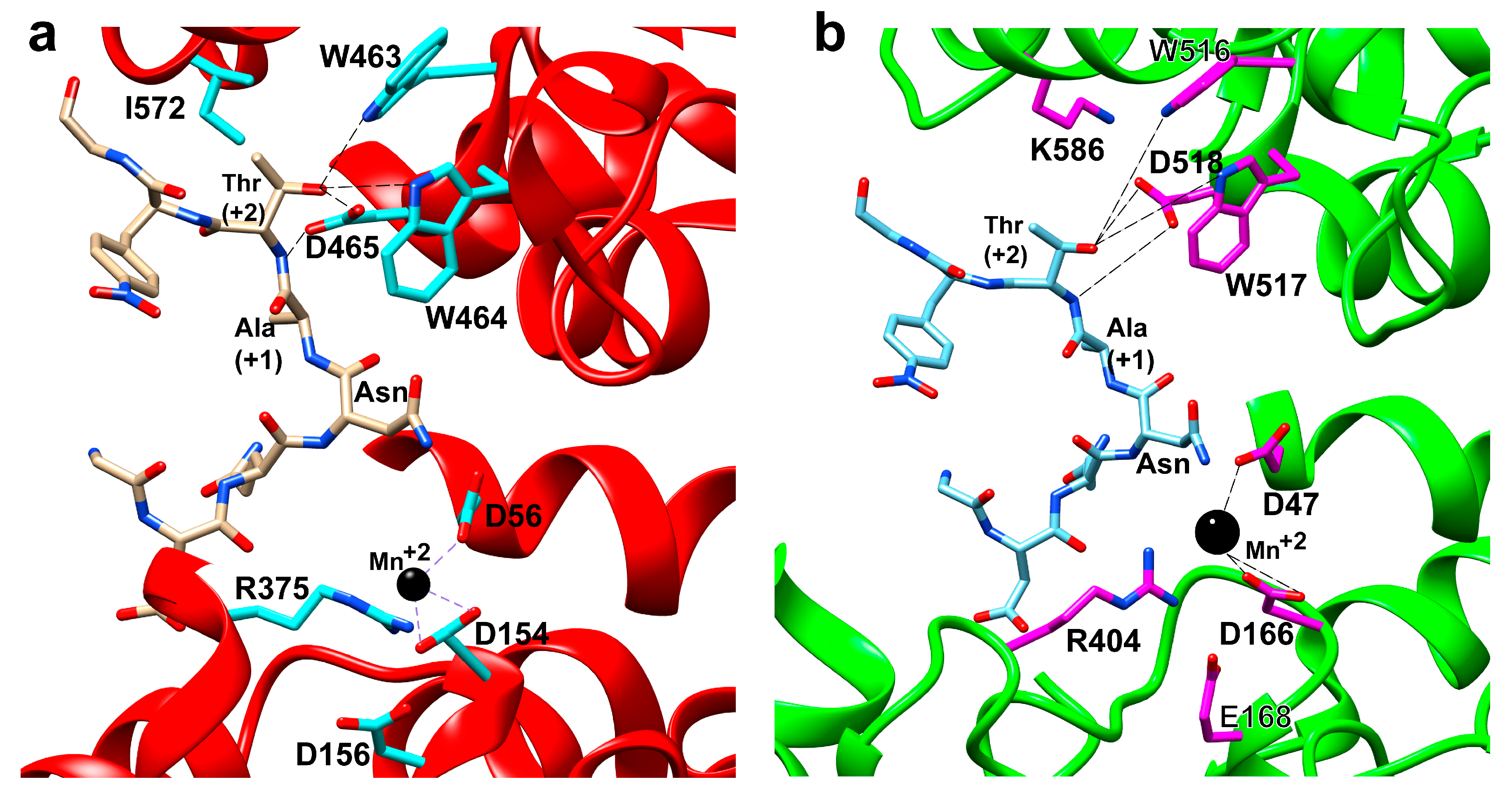
3.3.1. Glycosylation by the OST-A Isoform
3.3.2. Proofreading by the OST-B Complex
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Dempski, R.E., Jr.; Imperiali, B. Oligosaccharyl transferase: Gatekeeper to the secretory pathway. Curr. Opin. Chem. Biol. 2002, 6, 844–850. [Google Scholar] [CrossRef]
- Kornfeld, R.; Kornfeld, S. Assembly of asparagine-linked oligosaccharides. Annu. Rev. Biochem. 1985, 54, 631–664. [Google Scholar] [CrossRef] [PubMed]
- Larkin, A.; Imperiali, B. The expanding horizons of asparagine-linked glycosylation. Biochemistry 2011, 50, 4411–4426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacker, M.; Linton, D.; Hitchen, P.G.; Nita-Lazar, M.; Haslam, S.M.; North, S.J.; Panico, M.; Morris, H.R.; Dell, A.; Wren, B.W.; et al. N-linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science 2002, 298, 1790–1793. [Google Scholar] [CrossRef]
- Mohorko, E.; Glockshuber, R.; Aebi, M. Oligosaccharyltransferase: The central enzyme of N-linked protein glycosylation. J. Inherit. Metab. Dis. 2011, 34, 869–878. [Google Scholar] [CrossRef]
- Knauer, R.; Lehle, L. The oligosaccharyltransferase complex from yeast. Biochim. Biophys. Acta 1999, 1426, 259–273. [Google Scholar] [CrossRef]
- Welply, J.K.; Shenbagamurthi, P.; Lennarz, W.J.; Naider, F. Substrate recognition by oligosaccharyltransferase. Studies on glycosylation of modified Asn-X-Thr/Ser tripeptides. J. Biol. Chem. 1983, 258, 11856–11863. [Google Scholar]
- Gahmberg, C.G.; Tolvanen, M. Why mammalian cell surface proteins are glycoproteins. Trends Biochem. Sci. 1996, 21, 308–311. [Google Scholar] [CrossRef]
- Helenius, A.; Aebi, M. Roles of N-linked glycans in the endoplasmic reticulum. Annu. Rev. Biochem. 2004, 73, 1019–1049. [Google Scholar] [CrossRef]
- Freeze, H.H. Understanding human glycosylation disorders: Biochemistry leads the charge. J. Biol. Chem. 2013, 288, 6936–6945. [Google Scholar] [CrossRef] [Green Version]
- Hennet, T.; Cabalzar, J. Congenital disorders of glycosylation: A concise chart of glycocalyx dysfunction. Trends Biochem. Sci. 2015, 40, 377–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helenius, A. How N-linked oligosaccharides affect glycoprotein folding in the endoplasmic reticulum. Mol. Biol. Cell 1994, 5, 253–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulson, J.C. Glycoproteins: What are the sugar chains for? Trends Biochem. Sci. 1989, 14, 272–276. [Google Scholar] [CrossRef]
- Wormald, M.R.; Dwek, R.A. Glycoproteins: Glycan presentation and protein-fold stability. Structure 1999, 7, R155–R160. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, I.; Kelleher, D.J.; Miao, Y.; Shao, Y.; Kreibich, G.; Gilmore, R.; von Heijne, G.; Johnson, A.E. Photocross-linking of nascent chains to the STT3 subunit of the oligosaccharyltransferase complex. J. Cell Biol. 2003, 161, 715–725. [Google Scholar] [CrossRef]
- Spirig, U.; Bodmer, D.; Wacker, M.; Burda, P.; Aebi, M. The 3.4-kDa Ost4 protein is required for the assembly of two distinct oligosaccharyltransferase complexes in yeast. Glycobiology 2005, 15, 1396–1406. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, D.J.; Gilmore, R. An evolving view of the eukaryotic oligosaccharyltransferase. Glycobiology 2006, 16, 47R–62R. [Google Scholar] [CrossRef]
- Mueller, S.; Wahlander, A.; Selevsek, N.; Otto, C.; Ngwa, E.M.; Poljak, K.; Frey, A.D.; Aebi, M.; Gauss, R. Protein degradation corrects for imbalanced subunit stoichiometry in OST complex assembly. Mol. Biol. Cell 2015, 26, 2596–2608. [Google Scholar] [CrossRef] [Green Version]
- Zufferey, R.; Knauer, R.; Burda, P.; Stagljar, I.; te Heesen, S.; Lehle, L.; Aebi, M. STT3, a highly conserved protein required for yeast oligosaccharyl transferase activity in vivo. EMBO J. 1995, 14, 4949–4960. [Google Scholar] [CrossRef]
- Cherepanova, N.; Shrimal, S.; Gilmore, R. N-linked glycosylation and homeostasis of the endoplasmic reticulum. Curr. Opin. Cell Biol. 2016, 41, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Shrimal, S.; Cherepanova, N.A.; Gilmore, R. Cotranslational and posttranslocational N-glycosylation of proteins in the endoplasmic reticulum. Semin. Cell Dev. Biol. 2015, 41, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelleher, D.J.; Karaoglu, D.; Mandon, E.C.; Gilmore, R. Oligosaccharyltransferase isoforms that contain different catalytic STT3 subunits have distinct enzymatic properties. Mol. Cell 2003, 12, 101–111. [Google Scholar] [CrossRef]
- Mohorko, E.; Owen, R.L.; Malojcic, G.; Brozzo, M.S.; Aebi, M.; Glockshuber, R. Structural basis of substrate specificity of human oligosaccharyl transferase subunit N33/Tusc3 and its role in regulating protein N-glycosylation. Structure 2014, 22, 590–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, B.L.; Stirnimann, C.U.; Grimshaw, J.P.; Brozzo, M.S.; Fritsch, F.; Mohorko, E.; Capitani, G.; Glockshuber, R.; Grutter, M.G.; Aebi, M. Oxidoreductase activity of oligosaccharyltransferase subunits Ost3p and Ost6p defines site-specific glycosylation efficiency. Proc. Natl. Acad. Sci. USA 2009, 106, 11061–11066. [Google Scholar] [CrossRef] [Green Version]
- Gayen, S.; Kang, C. Solution structure of a human minimembrane protein Ost4, a subunit of the oligosaccharyltransferase complex. Biochem. Biophys. Res. Commun. 2011, 409, 572–576. [Google Scholar] [CrossRef]
- Zubkov, S.; Lennarz, W.J.; Mohanty, S. Structural basis for the function of a minimembrane protein subunit of yeast oligosaccharyltransferase. Proc. Natl. Acad. Sci. USA 2004, 101, 3821–3826. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Bhaskaran, R.; Mohanty, S. Eukaryotic N-glycosylation occurs via the membrane-anchored C-terminal domain of the Stt3p subunit of oligosaccharyltransferase. J. Biol. Chem. 2012, 287, 32450–32458. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Chavan, M.; Schindelin, H.; Lennarz, W.J.; Li, H. Structure of the oligosaccharyl transferase complex at 12 A resolution. Structure 2008, 16, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, S.; Dudek, J.; Gogala, M.; Schorr, S.; Linxweiler, J.; Lang, S.; Becker, T.; Beckmann, R.; Zimmermann, R.; Forster, F. Structure of the mammalian oligosaccharyl-transferase complex in the native ER protein translocon. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Wang, T.; Zhao, G.; Kovach, A.; Li, H. The atomic structure of a eukaryotic oligosaccharyltransferase complex. Nature 2018, 555, 328–333. [Google Scholar] [CrossRef]
- Wild, R.; Kowal, J.; Eyring, J.; Ngwa, E.M.; Aebi, M.; Locher, K.P. Structure of the yeast oligosaccharyltransferase complex gives insight into eukaryotic N-glycosylation. Science 2018, 359, 545–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, A.S.; Kowal, J.; Locher, K.P. Cryo-electron microscopy structures of human oligosaccharyltransferase complexes OST-A and OST-B. Science 2019, 366, 1372–1375. [Google Scholar] [CrossRef] [PubMed]
- Lizak, C.; Gerber, S.; Numao, S.; Aebi, M.; Locher, K.P. X-ray structure of a bacterial oligosaccharyltransferase. Nature 2011, 474, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Napiórkowska, M.; Boilevin, J.; Sovdat, T.; Darbre, T.; Reymond, J.-L.; Aebi, M.; Locher, K.P. Molecular basis of lipid-linked oligosaccharide recognition and processing by bacterial oligosaccharyltransferase. Nat. Struct. Mol. Biol. 2017, 24, 1100–1106. [Google Scholar] [CrossRef] [Green Version]
- Napiórkowska, M.; Boilevin, J.; Darbre, T.; Reymond, J.L.; Locher, K.P. Structure of bacterial oligosaccharyltransferase PglB bound to a reactive LLO and an inhibitory peptide. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maita, N.; Nyirenda, J.; Igura, M.; Kamishikiryo, J.; Kohda, D. Comparative structural biology of eubacterial and archaeal oligosaccharyltransferases. J. Biol. Chem. 2010, 285, 4941–4950. [Google Scholar] [CrossRef] [Green Version]
- Nyirenda, J.; Matsumoto, S.; Saitoh, T.; Maita, N.; Noda, N.N.; Inagaki, F.; Kohda, D. Crystallographic and NMR evidence for flexibility in oligosaccharyltransferases and its catalytic significance. Structure 2013, 21, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Shimada, A.; Kohda, D. Crystal structure of the C-terminal globular domain of the third paralog of the Archaeoglobus fulgidus oligosaccharyltransferases. BMC Struct. Biol. 2013, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Shimada, A.; Nyirenda, J.; Igura, M.; Kawano, Y.; Kohda, D. Crystal structures of an archaeal oligosaccharyltransferase provide insights into the catalytic cycle of N-linked protein glycosylation. Proc. Natl. Acad. Sci. USA 2013, 110, 17868–17873. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Taguchi, Y.; Shimada, A.; Igura, M.; Kohda, D. Tethering an N-glycosylation sequon-containing peptide creates a catalytically competent oligosaccharyltransferase complex. Biochemistry 2017, 56, 602–611. [Google Scholar] [CrossRef]
- Lechner, J.; Wieland, F. Structure and biosynthesis of prokaryotic glycoproteins. Annu. Rev. Biochem. 1989, 58, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, C.; Sonnenbichler, J.; Sonnenbichler, I.; Sumper, M.; Zeitler, R. Isolation and characterization of dolichol-linked oligosaccharides from Haloferax volcanii. Glycobiology 1997, 7, 897–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanski, C.M.; Yao, R.; Ewing, C.P.; Trust, T.J.; Guerry, P. Evidence for a system of general protein glycosylation in Campylobacter jejuni. Mol. Microbiol. 1999, 32, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Voisin, S.; Houliston, R.S.; Kelly, J.; Brisson, J.-R.; Watson, D.; Bardy, S.L.; Jarrell, K.F.; Logan, S.M. Identification and characterization of the unique N-linked glycan common to the flagellins and S-layer glycoprotein of Methanococcus voltae. J. Biol. Chem. 2005, 280, 16586–16593. [Google Scholar] [CrossRef] [Green Version]
- Wieland, F.; Paul, G.; Sumper, M. Halobacterial flagellins are sulfated glycoproteins. J. Biol. Chem. 1985, 260, 15180–15185. [Google Scholar]
- Chang, M.M.; Imperiali, B.; Eichler, J.; Guan, Z. N-linked glycans are assembled on highly reduced dolichol phosphate carriers in the hyperthermophilic archaea pyrococcus furiosus. PLoS ONE 2015, 10, e0130482. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, Y.; Fujinami, D.; Kohda, D. Comparative analysis of archaeal lipid-linked oligosaccharides that serve as oligosaccharide donors for asn glycosylation. J. Biol. Chem. 2016, 291, 11042–11054. [Google Scholar] [CrossRef] [Green Version]
- Fujinami, D.; Taguchi, Y.; Kohda, D. Asn-linked oligosaccharide chain of a crenarchaeon, Pyrobaculum calidifontis, is reminiscent of the eukaryotic high-mannose-type glycan. Glycobiology 2017, 27, 701–712. [Google Scholar] [CrossRef]
- Fujinami, D.; Nyirenda, J.; Matsumoto, S.; Kohda, D. Structural elucidation of an asparagine-linked oligosaccharide from the hyperthermophilic archaeon, Archaeoglobus fulgidus. Carbohydr. Res. 2015, 413, 55–62. [Google Scholar] [CrossRef]
- Young, N.M.; Brisson, J.R.; Kelly, J.; Watson, D.C.; Tessier, L.; Lanthier, P.H.; Jarrell, H.C.; Cadotte, N.; St Michael, F.; Aberg, E.; et al. Structure of the N-linked glycan present on multiple glycoproteins in the Gram-negative bacterium, Campylobacter jejuni. J. Biol. Chem. 2002, 277, 42530–42539. [Google Scholar] [CrossRef] [Green Version]
- Chapman, A.; Li, E.; Kornfeld, S. The biosynthesis of the major lipid-linked oligosaccharide of Chinese hamster ovary cells occurs by the ordered addition of mannose residues. J. Biol. Chem. 1979, 254, 10243–10249. [Google Scholar] [PubMed]
- Liu, T.; Stetson, B.; Turco, S.J.; Hubbard, S.C.; Robbins, P.W. Arrangement of glucose residues in the lipid-linked oligosaccharide precursor of asparaginyl oligosaccharides. J. Biol. Chem. 1979, 254, 4554–45549. [Google Scholar] [PubMed]
- Samuelson, J.; Banerjee, S.; Magnelli, P.; Cui, J.; Kelleher, D.J.; Gilmore, R.; Robbins, P.W. The diversity of dolichol-linked precursors to asn-linked glycans likely results from secondary loss of sets of glycosyltransferases. Proc. Natl. Acad. Sci. USA 2005, 102, 1548–1553. [Google Scholar] [CrossRef] [Green Version]
- Bause, E.; Legler, G. The role of the hydroxy amino acid in the triplet sequence Asn-Xaa-Thr(Ser) for the N-glycosylation step during glycoprotein biosynthesis. Biochem. J. 1981, 195, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Imperiali, B.; Shannon, K.L.; Unno, M.; Rickert, K.W. Mechanistic proposal for asparagine-linked glycosylation. J. Am. Chem. Soc. 1992, 114, 7944–7945. [Google Scholar] [CrossRef]
- Marshall, R.D. Glycoproteins. Annu. Rev. Biochem. 1972, 41, 673–702. [Google Scholar] [CrossRef]
- Kohda, D. Structural basis of protein asn-glycosylation by oligosaccharyltransferases. Adv. Exp. Med. Biol. 2018, 1104, 171–199. [Google Scholar]
- Lizak, C.; Gerber, S.; Michaud, G.; Schubert, M.; Fan, Y.-Y.; Bucher, M.; Darbre, T.; Aebi, M.; Reymond, J.-L.; Locher, K.P. Unexpected reactivity and mechanism of carboxamide activation in bacterial N-linked protein glycosylation. Nat. Commun. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Igura, M.; Maita, N.; Kamishikiryo, J.; Yamada, M.; Obita, T.; Maenaka, K.; Kohda, D. Structure-guided identification of a new catalytic motif of oligosaccharyltransferase. EMBO J. 2008, 27, 234–243. [Google Scholar] [CrossRef]
- Imperiali, B.; Moats, R.A.; Fisher, S.L.; Prins, T.J. A conformational study of peptides with the general structure Ac-L-Xaa-Pro-D-Xaa-L-Xaa-NH2: Spectroscopic evidence for a peptide with significant.beta.-turn character in water and in dimethyl sulfoxide. J. Am. Chem. Soc. 1992, 114, 3182–3188. [Google Scholar] [CrossRef]
- Breuer, W.; Klein, R.A.; Hardt, B.; Bartoschek, A.; Bause, E. Oligosaccharyltransferase is highly specific for the hydroxy amino acid in Asn-Xaa-Thr/Ser. FEBS Lett. 2001, 501, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.M.; Glover, K.J.; Imperiali, B. From peptide to protein: Comparative analysis of the substrate specificity of N-linked glycosylation in C. jejuni. Biochemistry 2007, 46, 5579–5585. [Google Scholar] [CrossRef] [PubMed]
- Gerber, S.; Lizak, C.; Michaud, G.; Bucher, M.; Darbre, T.; Aebi, M.; Reymond, J.L.; Locher, K.P. Mechanism of bacterial oligosaccharyltransferase: In vitro quantification of sequon binding and catalysis. J. Biol. Chem. 2013, 288, 8849–8861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaoglu, D.; Kelleher, D.J.; Gilmore, R. The highly conserved Stt3 protein is a subunit of the yeast oligosaccharyltransferase and forms a subcomplex with Ost3p and Ost4p. J. Biol. Chem. 1997, 272, 32513–32520. [Google Scholar] [CrossRef] [Green Version]
- Spirig, U.; Glavas, M.; Bodmer, D.; Reiss, G.; Burda, P.; Lippuner, V.; te Heesen, S.; Aebi, M. The STT3 protein is a component of the yeast oligosaccharyltransferase complex. Mol. Genet. Genom. MGG 1997, 256, 628–637. [Google Scholar] [CrossRef]
- Te Heesen, S.; Janetzky, B.; Lehle, L.; Aebi, M. The yeast WBP1 is essential for oligosaccharyl transferase activity in vivo and in vitro. EMBO J. 1992, 11, 2071–2075. [Google Scholar] [CrossRef]
- te Heesen, S.; Knauer, R.; Lehle, L.; Aebi, M. Yeast Wbp1p and Swp1p form a protein complex essential for oligosaccharyl transferase activity. EMBO J. 1993, 12, 279–284. [Google Scholar] [CrossRef]
- Kowarik, M.; Young, N.M.; Numao, S.; Schulz, B.L.; Hug, I.; Callewaert, N.; Mills, D.C.; Watson, D.C.; Hernandez, M.; Kelly, J.F.; et al. Definition of the bacterial N-glycosylation site consensus sequence. EMBO J. 2006, 25, 1957–1966. [Google Scholar] [CrossRef] [Green Version]
- Murray, A.N.; Chen, W.; Antonopoulos, A.; Hanson, S.R.; Wiseman, R.L.; Dell, A.; Haslam, S.M.; Powers, D.L.; Powers, E.T.; Kelly, J.W. Enhanced aromatic sequons increase oligosaccharyltransferase glycosylation efficiency and glycan homogeneity. Chem. Biol. 2015, 22, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Prestwich, G.D.; Lennarz, W.J. The Ost1p subunit of yeast oligosaccharyl transferase recognizes the peptide glycosylation site sequence, -Asn-X-Ser/Thr. J. Biol. Chem. 1999, 274, 5021–5025. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Park, H.; Montalvo, L.; Lennarz, W.J. Studies on the role of the hydrophobic domain of Ost4p in interactions with other subunits of yeast oligosaccharyl transferase. Proc. Natl. Acad. Sci. USA 2000, 97, 1516–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Yan, Q.; Von Heijne, G.; Caputo, G.A.; Lennarz, W.J. Determination of the membrane topology of Ost4p and its subunit interactions in the oligosaccharyltransferase complex in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2003, 100, 7460–7464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, B.; Mazumder, S.; Mohanty, S. Production and biophysical characterization of a mini-membrane protein, Ost4V23D: A functionally important mutant of yeast oligosaccharyltransferase subunit Ost4p. Protein Expr. Purif. 2017, 139, 43–48. [Google Scholar] [CrossRef]
- Kumar, A.; Ward, P.; Katre, U.V.; Mohanty, S. A novel and simple method of production and biophysical characterization of a mini-membrane protein, Ost4p: A subunit of yeast oligosaccharyl transferase. Biopolymers 2012, 97, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, R.; Hendrickson, T.L.; Imperiali, B. Sulfhydryl modification of the yeast Wbp1p inhibits oligosaccharyl transferase activity. Biochemistry 1995, 34, 4179–4185. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Lennarz, W.J. Studies on the function of oligosaccharyl transferase subunits. Stt3p is directly involved in the glycosylation process. J. Biol. Chem. 2002, 277, 47692–47700. [Google Scholar] [CrossRef] [Green Version]
- Shrimal, S.; Gilmore, R. Oligosaccharyltransferase structures provide novel insight into the mechanism of asparagine-linked glycosylation in prokaryotic and eukaryotic cells. Glycobiology 2019, 29, 288–297. [Google Scholar] [CrossRef]
- Chavan, M.; Yan, A.; Lennarz, W.J. Subunits of the translocon interact with components of the oligosaccharyl transferase complex. J. Biol. Chem. 2005, 280, 22917–22924. [Google Scholar] [CrossRef] [Green Version]
- Beatson, S.; Ponting, C.P. GIFT domains: Linking eukaryotic intraflagellar transport and glycosylation to bacterial gliding. Trends Biochem. Sci. 2004, 29, 396–399. [Google Scholar] [CrossRef]
- Harada, Y.; Li, H.; Li, H.; Lennarz, W.J. Oligosaccharyltransferase directly binds to ribosome at a location near the translocon-binding site. Proc. Natl. Acad. Sci. USA 2009, 106, 6945–6949. [Google Scholar] [CrossRef] [Green Version]
- Yan, A.; Lennarz, W.J. Two oligosaccharyl transferase complexes exist in yeast and associate with two different translocons. Glycobiology 2005, 15, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Ahmed, E.; Yan, Q.; Lennarz, W.J. New findings on interactions among the yeast oligosaccharyl transferase subunits using a chemical cross-linker. J. Biol. Chem. 2003, 278, 33078–33087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunger, K.; Pfeffer, S.; Shrimal, S.; Gilmore, R.; Berninghausen, O.; Mandon, E.C.; Becker, T.; Forster, F.; Beckmann, R. Structural basis for coupling protein transport and N-glycosylation at the mammalian endoplasmic reticulum. Science 2018, 360, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrimal, S.; Cherepanova, N.A.; Gilmore, R. DC2 and KCP2 mediate the interaction between the oligosaccharyltransferase and the ER translocon. J. Cell Biol. 2017, 216, 3625–3638. [Google Scholar] [CrossRef]
- Cherepanova, N.A.; Venev, S.V.; Leszyk, J.D.; Shaffer, S.A.; Gilmore, R. Quantitative glycoproteomics reveals new classes of STT3A- and STT3B-dependent N-glycosylation sites. J. Cell Biol. 2019, 218, 2782–2796. [Google Scholar] [CrossRef] [Green Version]
- Galli, C.; Bernasconi, R.; Soldà, T.; Calanca, V.; Molinari, M. Malectin participates in a backup glycoprotein quality control pathway in the mammalian ER. PLoS ONE 2011, 6, e16304. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K.; Qin, S.-Y.; Matsumoto, N.; Yamamoto, K. Association of malectin with ribophorin I is crucial for attenuation of misfolded glycoprotein secretion. Biochem. Biophys. Res. Commun. 2014, 454, 436–440. [Google Scholar] [CrossRef]
- Qin, S.-Y.; Hu, D.; Matsumoto, K.; Takeda, K.; Matsumoto, N.; Yamaguchi, Y.; Yamamoto, K. Malectin forms a complex with ribophorin I for enhanced association with misfolded glycoproteins. J. Biol. Chem. 2012, 287, 38080–38089. [Google Scholar] [CrossRef] [Green Version]
- Goytain, A.; Quamme, G.A. Identification and characterization of a novel mammalian Mg2+ transporter with channel-like properties. BMC Genom. 2005, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Canada, C.; Kelleher, D.J.; Gilmore, R. Cotranslational and posttranslational N-glycosylation of polypeptides by distinct mammalian OST isoforms. Cell 2009, 136, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Shibatani, T.; David, L.L.; McCormack, A.L.; Frueh, K.; Skach, W.R. Proteomic analysis of mammalian oligosaccharyltransferase reveals multiple subcomplexes that contain Sec61, TRAP, and two potential new subunits. Biochemistry 2005, 44, 5982–5992. [Google Scholar] [CrossRef] [PubMed]
- Conti, B.J.; Devaraneni, P.K.; Yang, Z.; David, L.L.; Skach, W.R. Cotranslational stabilization of Sec62/63 within the ER Sec61 translocon is controlled by distinct substrate-driven translocation events. Mol. Cell 2015, 58, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Shrimal, S.; Cherepanova, N.A.; Mandon, E.C.; Venev, S.V.; Gilmore, R. Asparagine-linked glycosylation is not directly coupled to protein translocation across the endoplasmic reticulum in Saccharomyces cerevisiae. Mol. Biol. Cell 2019, 30, 2626–2638. [Google Scholar] [CrossRef]
- Shrimal, S.; Gilmore, R. Glycosylation of closely spaced acceptor sites in human glycoproteins. J. Cell Sci. 2013, 126, 5513–5523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherepanova, N.A.; Shrimal, S.; Gilmore, R. Oxidoreductase activity is necessary for N-glycosylation of cysteine-proximal acceptor sites in glycoproteins. J. Cell Biol. 2014, 206, 525–539. [Google Scholar] [CrossRef] [Green Version]
- Shrimal, S.; Trueman, S.F.; Gilmore, R. Extreme C-terminal sites are posttranslocationally glycosylated by the STT3B isoform of the OST. J. Cell Biol. 2013, 201, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Malaby, H.L.H.; Kobertz, W.R. The middle X residue influences cotranslational N-glycosylation consensus site skipping. Biochemistry 2014, 53, 4884–4893. [Google Scholar] [CrossRef]
- Bas, T.; Gao, G.Y.; Lvov, A.; Chandrasekhar, K.D.; Gilmore, R.; Kobertz, W.R. Post-translational N-glycosylation of type I transmembrane KCNE1 peptides: Implications for membrane protein biogenesis and disease. J. Biol. Chem. 2011, 286, 28150–28159. [Google Scholar] [CrossRef] [Green Version]
- Malaby, H.L.H.; Kobertz, W.R. Molecular determinants of co- and post-translational N-glycosylation of type I transmembrane peptides. Biochem. J. 2013, 453, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Dell, A.; Galadari, A.; Sastre, F.; Hitchen, P. Similarities and differences in the glycosylation mechanisms in prokaryotes and eukaryotes. Int. J. Microbiol. 2010, 2010, 148178. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Archaea | Bacteria | Yeast | Human | Function | |
|---|---|---|---|---|---|
| ssOST | ssOST | OST complex | OST-A complex | OST-B complex | |
| AglB | PglB | Stt3 | STT3A | STT3B | Catalytic activity |
| Ost4 | OST4 | OST4 | Maintains stability of catalytic sub-complex | ||
| Ost3/Ost6 | MAGT1|N33/TUSC3 | Oxidoreductase activity | |||
| Ost5 | TMEM258 | TMEM258 | Not clear | ||
| Ost1 | RPN1 | RPN1 | Restrains glycosylated peptide from sliding back to the catalytic site | ||
| Ost2 | DAD1 | DAD1 | Not clear | ||
| Wbp1 | OST48 | OST48 | Possibly LLO recruitment | ||
| Swp1 | RPN2 | RPN2 | Possibly LLO recruitment | ||
| KCP2 | Mediates interaction with translocon channel | ||||
| DC2 | Mediates interaction with translocon channel | ||||
| Prokaryotic Oligosaccharyltransferase | References | ||
|---|---|---|---|
| Bacterial Oligosaccharyltransferase | |||
| Protein | Method | PDB ID | |
| Compylobactor lari PglB with acceptor peptide | X-ray | 3RCE | [33] |
| Compylobactor lari PglB with acceptor peptide and LLO analog | X-ray | 5OGL | [34] |
| Compylobactor lari PglB with inhibitory peptide and reactive LLO | X-ray | 6GXC | [35] |
| C-terminal domain of Compylobactor jejuni PglB | X-ray | 3AAG | [36] |
| Archaeal Oligosaccharyltransferase | |||
| C-terminal soluble domain from Archaeoglobus fulgidus | X-ray | 3VU0 | [37] |
| C-terminal soluble domain from Pyrococcus horikoshii | X-ray | 3VU1 | [37] |
| C-term globular domain as MBP fusion from Archaeoglobus fulgidus | X-ray | 3WAI | [38] |
| Archaeoglobus fulgidus AglB | X-ray | 3WAK | [39] |
| Archaeoglobus fulgidus AglB with Zn and sulfate | X-ray | 3WAJ | [39] |
| Archaeoglobus fulgidus AglB with acceptor peptide | X-ray | 5GMY | [40] |
| Eukaryotic Oligosaccharyltransferase | |||
| (1) Yeast Oligosaccharyltransferase (OST) | |||
| Yeast OST subunit Ost4p | Solution NMR | 1RKL | [26] |
| Oxidized Ost6L | X-ray | 3G7Y | [24] |
| Reduced Ost6L | X-ray | 3G9B | [24] |
| Photo-reduced Ost6L | X-ray | 3GA4 | [24] |
| C-terminal domain of Stt3p subunit | Solution NMR | 2LGZ | [27] |
| OST complex | Cryo-EM | 6EZN | [31] |
| OST complex | Cryo-EM | 6C26 | [30] |
| (2) Human Oligosaccharyltransferase | |||
| Soluble N-terminal domain of N33/Tusc3 subunit | X-ray | 4M90, 4M91, 4M92, and 4M8G | [23] |
| Ost4 subunit | Solution NMR | 2LAT | [25] |
| OST-A complex | Cryo-EM | 6S7O | [32] |
| OST-B complex | Cryo-EM | 6S7T | [32] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohanty, S.; P Chaudhary, B.; Zoetewey, D. Structural Insight into the Mechanism of N-Linked Glycosylation by Oligosaccharyltransferase. Biomolecules 2020, 10, 624. https://doi.org/10.3390/biom10040624
Mohanty S, P Chaudhary B, Zoetewey D. Structural Insight into the Mechanism of N-Linked Glycosylation by Oligosaccharyltransferase. Biomolecules. 2020; 10(4):624. https://doi.org/10.3390/biom10040624
Chicago/Turabian StyleMohanty, Smita, Bharat P Chaudhary, and David Zoetewey. 2020. "Structural Insight into the Mechanism of N-Linked Glycosylation by Oligosaccharyltransferase" Biomolecules 10, no. 4: 624. https://doi.org/10.3390/biom10040624
APA StyleMohanty, S., P Chaudhary, B., & Zoetewey, D. (2020). Structural Insight into the Mechanism of N-Linked Glycosylation by Oligosaccharyltransferase. Biomolecules, 10(4), 624. https://doi.org/10.3390/biom10040624