PIRT the TRP Channel Regulating Protein Binds Calmodulin and Cholesterol-Like Ligands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rosetta Flexible Peptide and High-Resolution Docking
2.2. Protein Expression and Purification
2.3. Microscale Thermophoresis
2.4. PIRT and Calmodulin Pull-Down Assay
2.5. Nuclear Magnetic Resonance-Detected Binding Assay
3. Results
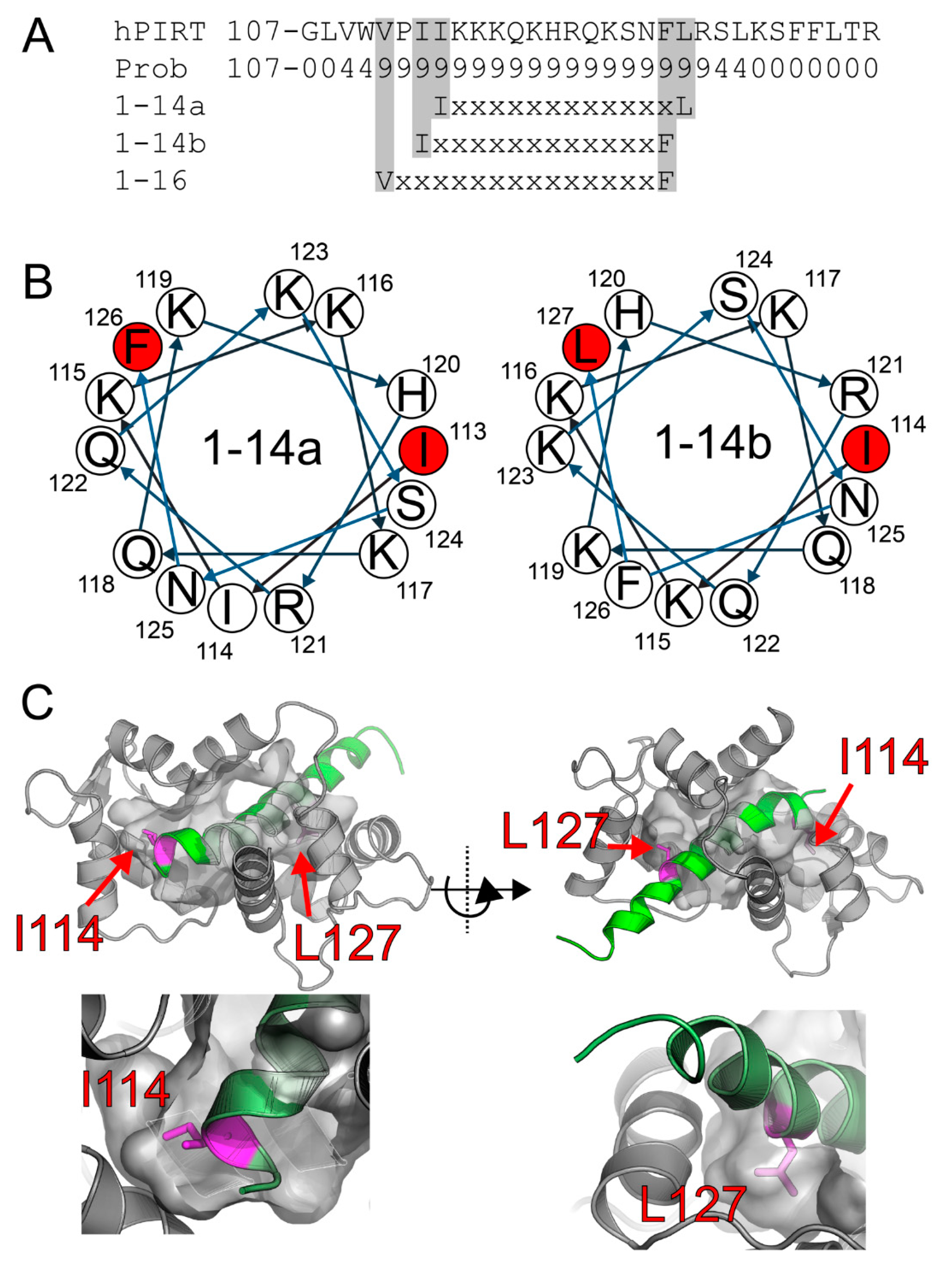
3.1. Bioinformatic Analysis of PIRT Predicts Calmodulin-Binding Motifs
3.2. Flex Pep Dock
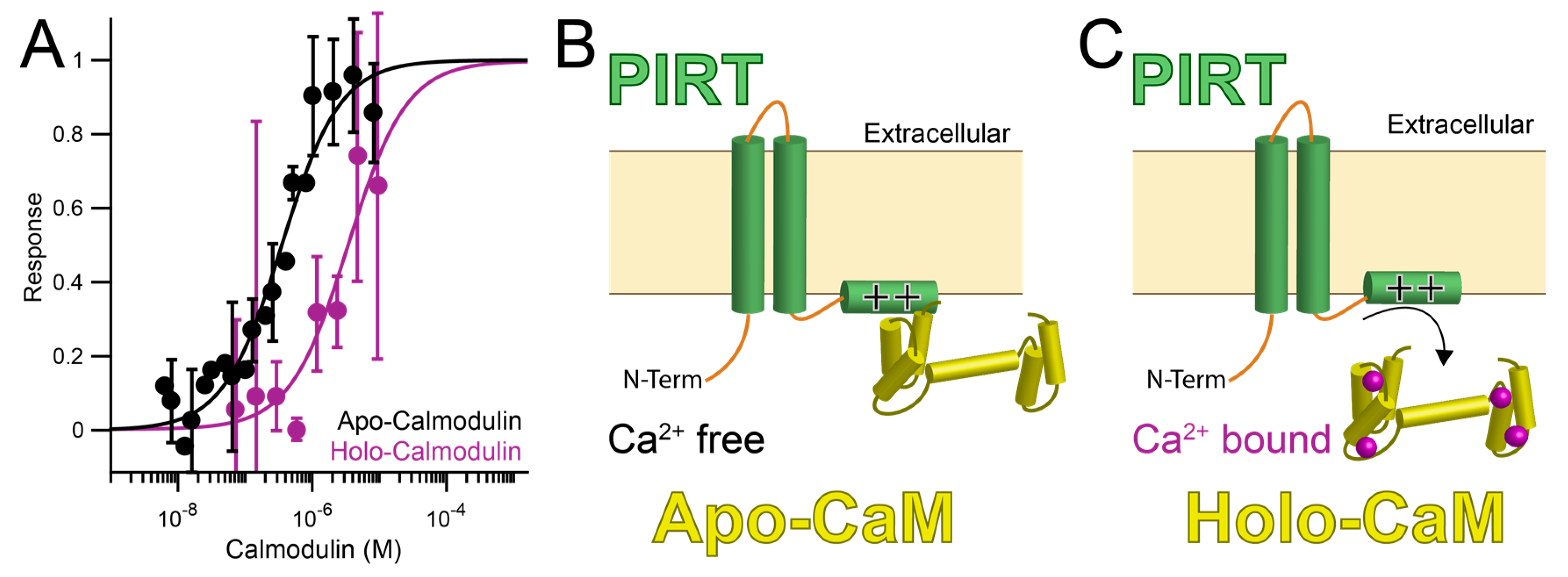
3.3. PIRT Specifically Interacts with Calmodulin
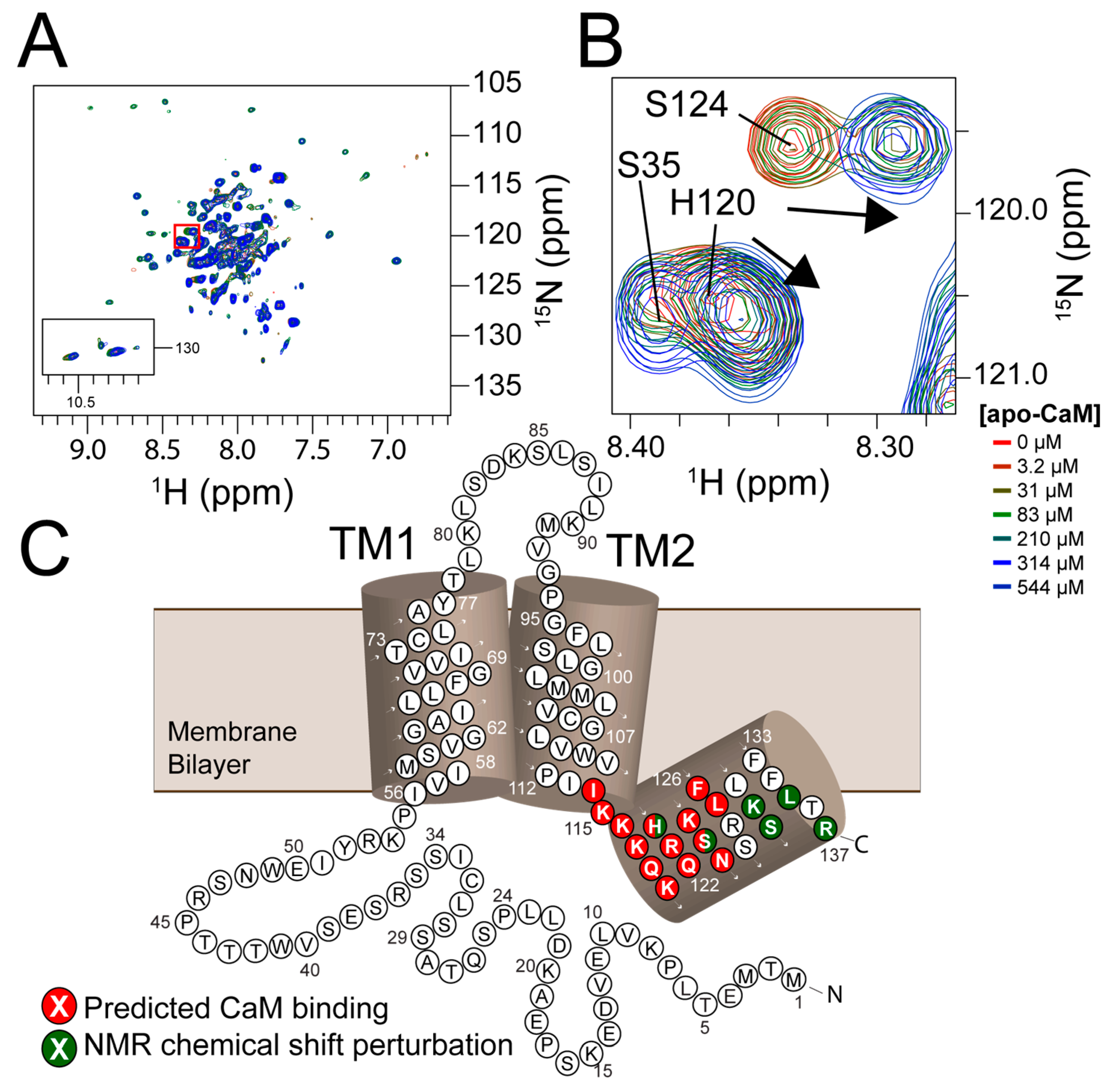
3.4. NMR Titration Shows Residues with Ligand-Dependent Perturbation Including the Predicted Calmodulin Binding Site
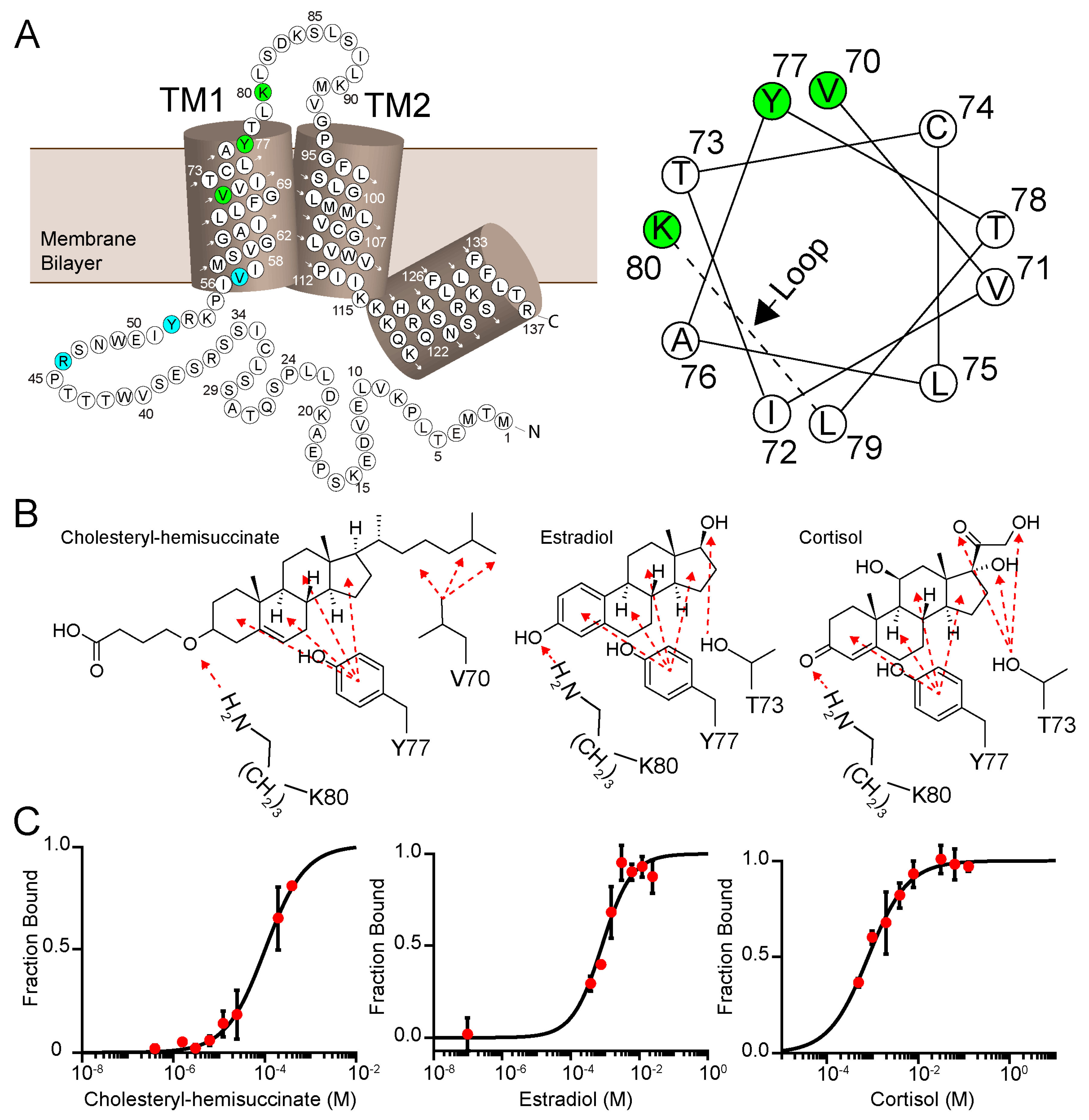
3.5. Two Cholesterol-Binding Motifs Found on PIRT with Bioinformatic Analysis
3.6. PIRT and Cholesterol Ligands Screening with Microscale Thermophoresis
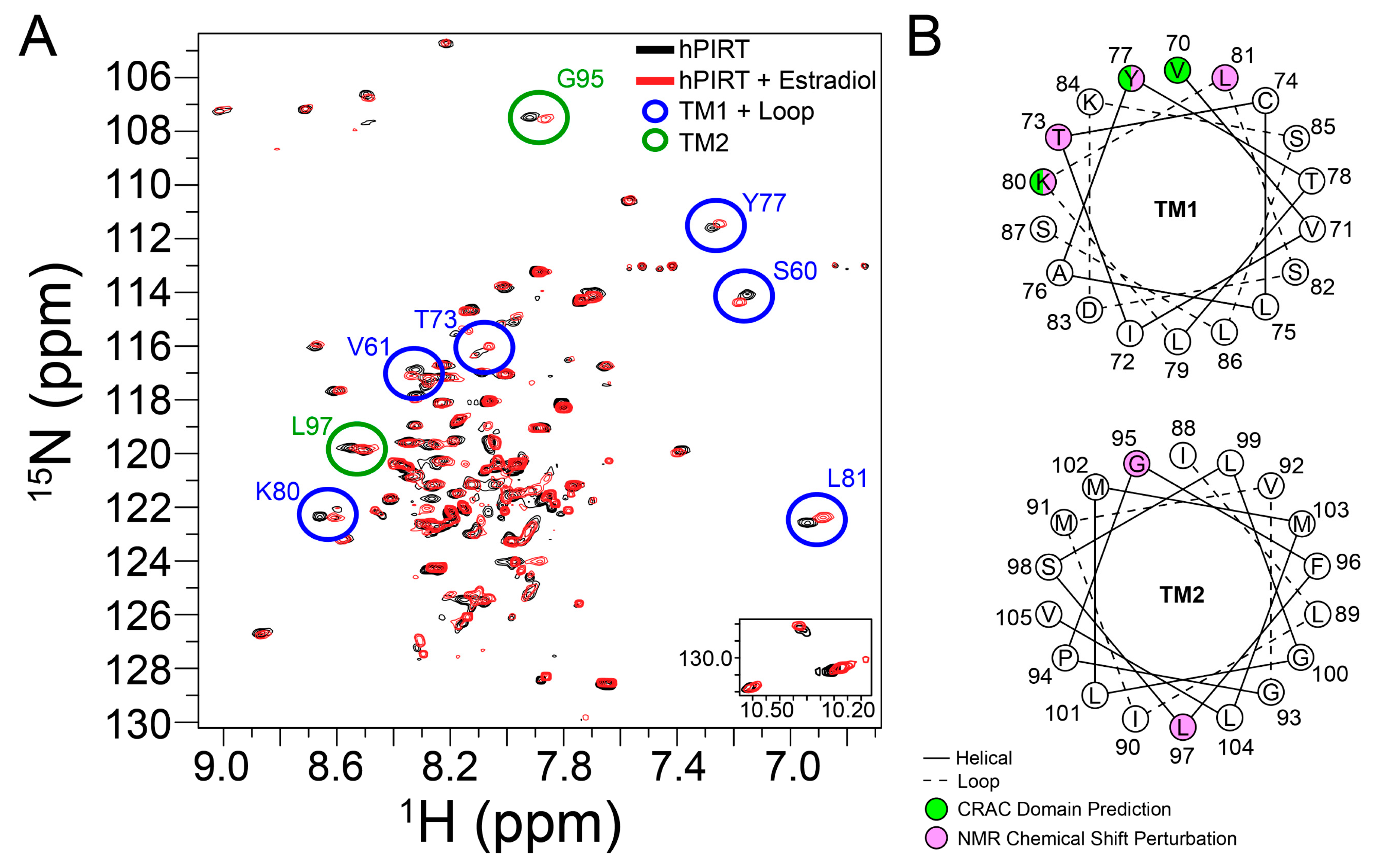
3.7. β-Estradiol Induced PIRT Chemical Shift Perturbation
3.8. Microscale Thermophoresis Shows PIRT Binds to Other Trp Channel Modulators
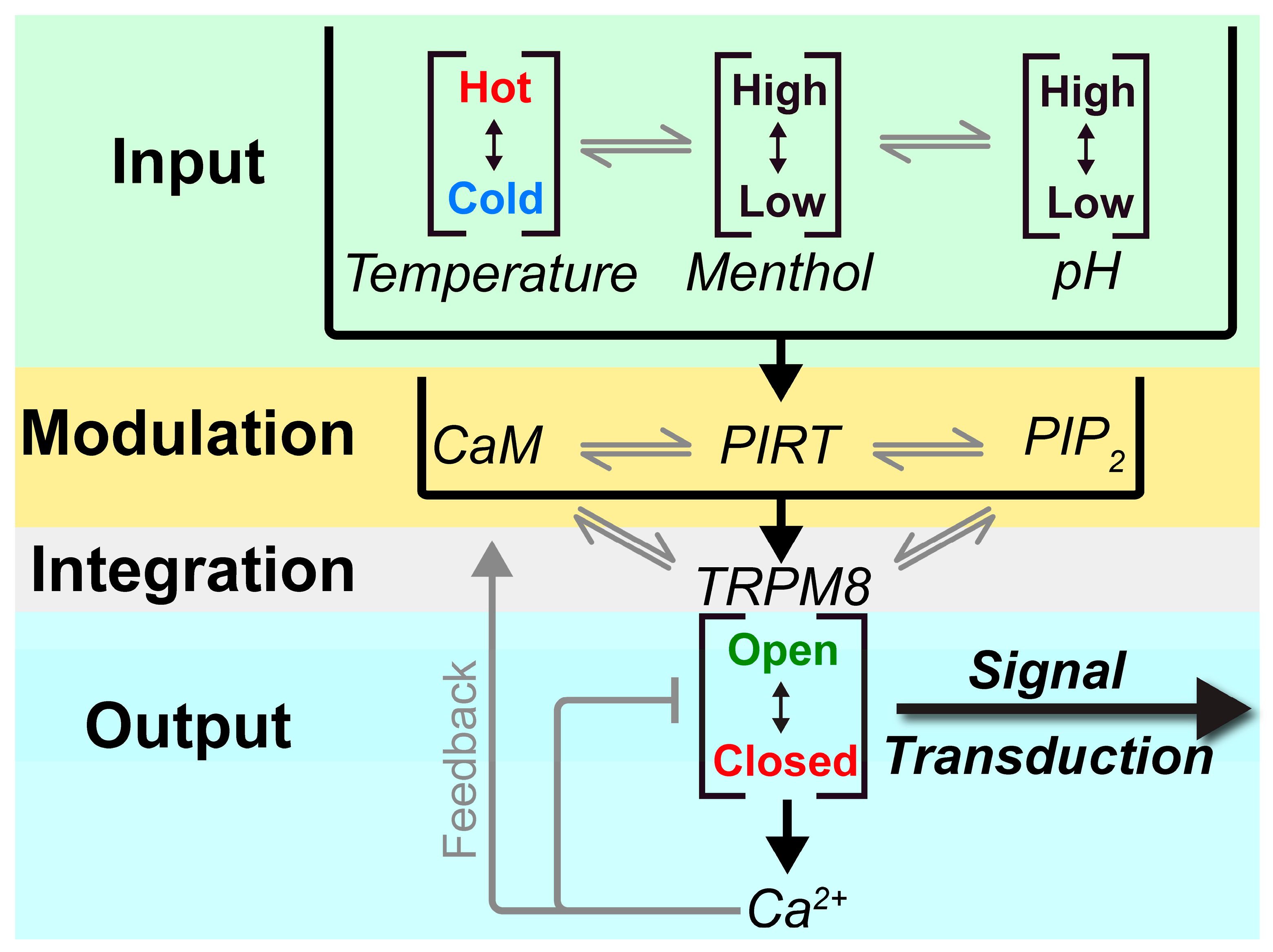
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Veldhuis, N.A.; Poole, D.P.; Grace, M.; McIntyre, P.; Bunnett, N.W. The G protein-coupled receptor-transient receptor potential channel axis: Molecular insights for targeting disorders of sensation and inflammation. Pharmacol. Rev. 2015, 67, 36–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proudfoot, C.J.; Garry, E.M.; Cottrell, D.F.; Rosie, R.; Anderson, H.; Robertson, D.C.; Fleetwood-Walker, S.M.; Mitchell, R. Analgesia mediated by the TRPM8 cold receptor in chronic neuropathic pain. Curr. Biol. 2006, 16, 1591–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsavaler, L.; Shapero, M.H.; Morkowski, S.; Laus, R. Trp-p8, a novel prostate-specific gene, is up-regulated in prostate cancer and other malignancies and shares high homology with transient receptor potential calcium channel proteins. Cancer Res. 2001, 61, 3760–3769. [Google Scholar] [PubMed]
- Zhang, L.; Barritt, G.J. Evidence that TRPM8 is an androgen-dependent Ca2+ channel required for the survival of prostate cancer cells. Cancer Res. 2004, 64, 8365–8373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Yu, H.; Zhao, Z.; Luo, Z.; Chen, J.; Ni, Y.; Jin, R.; Ma, L.; Wang, P.; Zhu, Z.; et al. Activation of the cold-sensing TRPM8 channel triggers UCP1-dependent thermogenesis and prevents obesity. J. Mol. Cell Biol. 2012, 4, 88–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, H.L.; Jenkins, A.C.; Kaufman, J.; Bhattacharyya, I.; Caudle, R.M.; Neubert, J.K. Characterization of bilateral trigeminal constriction injury using an operant facial pain assay. Neuroscience 2012, 224, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Garami, A.; Shimansky, Y.P.; Rumbus, Z.; Vizin, R.C.L.; Farkas, N.; Hegyi, J.; Szakacs, Z.; Solymar, M.; Csenkey, A.; Chiche, D.A.; et al. Hyperthermia induced by transient receptor potential vanilloid-1 (TRPV1) antagonists in human clinical trials: Insights from mathematical modeling and meta-analysis. Pharmacol. Tarmacol. 2020, 1, 107474. [Google Scholar] [CrossRef]
- Koplas, P.A.; Rosenberg, R.L.; Oxford, G.S. The role of calcium in the desensitization of capsaicin responses in rat dorsal root ganglion neurons. J. Neurosci. Res. 1997, 17, 3525–3537. [Google Scholar] [CrossRef]
- Hetenyi, A.; Nemeth, L.; Weber, E.; Szakonyi, G.; Winter, Z.; Josvay, K.; Bartus, E.; Olah, Z.; Martinek, T.A. Competitive inhibition of TRPV1-calmodulin interaction by vanilloids. FEBS Lett. 2016, 590, 2768–2775. [Google Scholar] [CrossRef] [Green Version]
- Grycova, L.; Holendova, B.; Lansky, Z.; Bumba, L.; Jirku, M.; Bousova, K.; Teisinger, J. Ca(2+) binding protein S100A1 competes with calmodulin and PIP2 for binding site on the C-terminus of the TPRV1 receptor. ACS Chem. Neurosci. 2015, 6, 386–392. [Google Scholar] [CrossRef]
- Kim, A.Y.; Tang, Z.; Liu, Q.; Patel, K.N.; Maag, D.; Geng, Y.; Dong, X. Pirt, a phosphoinositide-binding protein, functions as a regulatory subunit of TRPV1. Cell 2008, 133, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.N.; Liu, Q.; Meeker, S.; Undem, B.J.; Dong, X. Pirt, a TRPV1 modulator, is required for histamine-dependent and -independent itch. PLoS ONE 2011, 6, e20559. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Yang, Y.; Zhu, C.; Wu, G.; Yu, G.; Jian, T.; Yang, N.; Shi, H.; Tang, M.; et al. Pirt contributes to uterine contraction-induced pain in mice. Mol. Pain 2015, 11, 57. [Google Scholar] [CrossRef] [Green Version]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Sarria, I.; Ling, J.; Zhu, M.X.; Gu, J.G. TRPM8 acute desensitization is mediated by calmodulin and requires PIP2: Distinction from tachyphylaxis. J. Neurophysiol. 2011, 106, 3056–3066. [Google Scholar] [CrossRef] [Green Version]
- Halling, D.B.; Liebeskind, B.J.; Hall, A.W.; Aldrich, R.W. Conserved properties of individual Ca2+-binding sites in calmodulin. Proc. Natl. Acad. Sci. USA 2016, 113, E1216–E1225. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; McGoldrick, L.L.; Twomey, E.C.; Sobolevsky, A.I. Mechanism of calmodulin inactivation of the calcium-selective TRP channel TRPV6. Sci. Adv. 2018, 4, eaau6088. [Google Scholar] [CrossRef] [Green Version]
- Kovalevskaya, N.V.; van de Waterbeemd, M.; Bokhovchuk, F.M.; Bate, N.; Bindels, R.J.; Hoenderop, J.G.; Vuister, G.W. Structural analysis of calmodulin binding to ion channels demonstrates the role of its plasticity in regulation. Pflugers Arch. 2013, 465, 1507–1519. [Google Scholar] [CrossRef]
- Han, B.; He, K.; Cai, C.; Tang, Y.; Yang, L.; Heinemann, S.H.; Hoshi, T.; Hou, S. Human EAG channels are directly modulated by PIP2 as revealed by electrophysiological and optical interference investigations. Sci. Rep. 2016, 6, 23417. [Google Scholar] [CrossRef]
- Sun, J.; MacKinnon, R. Cryo-EM structure of a KCNQ1/CaM complex reveals insights into congenital long QT syndrome. Cell 2017, 169, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Rohacs, T.; Lopes, C.M.; Michailidis, I.; Logothetis, D.E. PI(4,5)P2 regulates the activation and desensitization of TRPM8 channels through the TRP domain. Nat. Neurosci. 2005, 8, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Hilton, J.K.; Salehpour, T.; Sisco, N.J.; Rath, P.; Van Horn, W.D. Phosphoinositide-interacting regulator of TRP (PIRT) has opposing effects on human and mouse TRPM8 ion channels. J. Biol. Chem. 2018, 293, 9423–9434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisco, N.J.; Helsell, C.V.M.; Van Horn, W.D. Competitive interactions between PIRT, the cold sensing ion channel TRPM8, and PIP2 suggest a mechanism for regulation. Sci. Rep. 2019, 9, 14128. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gu, L.; Ruan, Y.; Gegen, T.; Yu, L.; Zhu, C.; Yang, Y.; Zhou, Y.; Yu, G.; Tang, Z. Pirt together with TRPV1 is involved in the regulation of neuropathic pain. Neural Plast. 2018, 2018, 4861491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.F.; Feng, J.F.; Wang, W.; Xiang, Z.H.; Liu, X.J.; Zhu, C.; Tang, Z.X.; Dong, X.Z.; He, C. Pirt reduces bladder overactivity by inhibiting purinergic receptor P2 × 3. Nat. Commun. 2015, 6, 7650. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Sui, Q.Q.; Gao, X.F.; Feng, J.F.; Zhu, J.; He, C.; Knight, G.E.; Burnstock, G.; Xiang, Z. Co-localization of Pirt protein and P2 × 2 receptors in the mouse enteric nervous system. Purinergic Signal. 2016, 12, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; MacKinnon, R. Structural basis of human KCNQ1 modulation and gating. Cell 2020, 180, 340–347. [Google Scholar] [CrossRef]
- Cao, C.; Zakharian, E.; Borbiro, I.; Rohacs, T. Interplay between calmodulin and phosphatidylinositol 4,5-bisphosphate in Ca2+-induced inactivation of transient receptor potential vanilloid 6 channels. J. Biol. Chem. 2013, 288, 5278–5290. [Google Scholar] [CrossRef] [Green Version]
- Ziemba, B.P.; Swisher, G.H.; Masson, G.; Burke, J.E.; Williams, R.L.; Falke, J.J. Regulation of a coupled MARCKS-PI3K lipid kinase circuit by calmodulin: Single-molecule analysis of a membrane-bound signaling module. Biochemistry 2016, 55, 6395–6405. [Google Scholar] [CrossRef] [Green Version]
- Brewer, G.J. Calmodulin, zinc, and calcium in cellular and membrane regulation: An interpretive review. Am. J. Hematol. 1980, 8, 231–248. [Google Scholar] [CrossRef]
- Means, A.R.; Dedman, J.R. Calmodulin—An intracellular calcium receptor. Nature 1980, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W.E. Calcium-Binding Proteins of the EF-Hand Superfamily from Basics to Medical Applications; Heizmann, C.W., Ed.; Springer: New York, NY, USA, 2019. [Google Scholar] [CrossRef]
- Yin, Y.; Le, S.C.; Hsu, A.L.; Borgnia, M.J.; Yang, H.; Lee, S.-Y. Structural basis of cooling agent and lipid sensing by the cold-activated TRPM8 channel. Science 2019, 9334, eaav9334. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Cordero-Morales, J.F.; Liu, B.; Qin, F.; Julius, D. TRPV1 channels are intrinsically heat sensitive and negatively regulated by phosphoinositide lipids. Neuron 2013, 77, 667–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, M.D.; Gordon, S.E. Short-chain phosphoinositide partitioning into plasma membrane models. Biophys. J. 2013, 105, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauchi, S.; Orta, G.; Mascayano, C.; Salazar, M.; Raddatz, N.; Urbina, H.; Rosenmann, E.; Gonzalez-Nilo, F.; Latorre, R. Dissection of the components for PIP2 activation and thermosensation in TRP channels. Proc. Natl. Acad. Sci. USA 2007, 104, 10246–10251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Qin, F. Functional control of cold- and menthol-sensitive TRPM8 ion channels by phosphatidylinositol 4,5-bisphosphate. J. Neurosci. 2005, 25, 1674–1681. [Google Scholar] [CrossRef]
- Nelson, M.R.; Chazin, W.J. Structures of EF-hand Ca2+-binding proteins: Diversity in the organization, packing and response to Ca2+ binding. BioMetals 1998, 11, 297–318. [Google Scholar] [CrossRef]
- Fantini, J.; Barrantes, F.J. How cholesterol interacts with membrane proteins: An exploration of cholesterol-binding sites including CRAC, CARC, and tilted domains. Front. Physiol. 2013, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Raveh, B.; London, N.; Zimmerman, L.; Schueler-Furman, O. Rosetta FlexPepDock ab-initio: Simultaneous folding, docking and refinement of peptides onto their receptors. PLoS ONE 2011, 6, e18934. [Google Scholar] [CrossRef] [Green Version]
- Vernon, R.; Shen, Y.; Baker, D.; Lange, O.F. Improved chemical shift based fragment selection for CS-Rosetta using Rosetta3 fragment picker. J. Biomol. NMR 2013, 57, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Conway, P.; Tyka, M.D.; DiMaio, F.; Konerding, D.E.; Baker, D. Relaxation of backbone bond geometry improves protein energy landscape modeling. Protein Sci. 2014, 23, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Rath, P.; Hilton, J.K.; Sisco, N.J.; Van Horn, W.D. Implications of human transient receptor potential melastatin 8 (TRPM8) channel gating from menthol binding studies of the sensing domain. Biochemistry 2016, 55, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Wienken, C.J.; Baaske, P.; Rothbauer, U.; Braun, D.; Duhr, S. Protein-binding assays in biological liquids using microscale thermophoresis. Nat. Commun. 2010, 1, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Sisco, N.J.; Hilton, J.K.; Montano, C.M.; Castro, M.A.; Cherry, B.R.; Levitus, M.; Van Horn, W.D. Evidence that the TRPV1 S1–S4 membrane domain contributes to thermosensing. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Delaglio, F.; Grzesiek, S.; Vuister, G.W.; Zhu, G.; Pfeifer, J.; Bax, A. NMRPipe: A multidimensional spectral processing system based on UNIX pipes. J. Biomol. NMR 1995, 6, 277–293. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Yap, K.L.; Kim, J.; Truong, K.; Sherman, M.; Yuan, T.; Ikura, M. Calmodulin target database. J. Struct. Funct. Genom. 2000, 1, 8–14. [Google Scholar] [CrossRef]
- White, R.R.; Kwon, Y.-G.; Taing, M.; Lawrence, D.S.; Edelman, A.M. Definition of optimal substrate recognition motifs of Ca2+-calmodulin-dependent protein kinases IV and II reveals shared and distinctive features. J. Biol. Chem. 1998, 273, 3166–3172. [Google Scholar] [CrossRef] [Green Version]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The european molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Walters, J.R.; Balesaria, S.; Chavele, K.M.; Taylor, V.; Berry, J.L.; Khair, U.; Barley, N.F.; van Heel, D.A.; Field, J.; Hayat, J.O.; et al. Calcium channel TRPV6 expression in human duodenum: Different relationships to the vitamin D system and aging in men and women. J. Bone Miner. Res. 2006, 21, 1770–1777. [Google Scholar] [CrossRef] [PubMed]
- Rohacs, T. Phosphoinositide Regulation of TRP Channels. Handb. Exp. Pharmacol. 2014, 223, 1143–1176. [Google Scholar] [PubMed] [Green Version]
- Nilius, B.; Owsianik, G.; Voets, T. Transient receptor potential channels meet phosphoinositides. EMBO J. 2008, 27, 2809–2816. [Google Scholar] [CrossRef]
- Gordon-Shaag, A.; Zagotta, W.N.; Gordon, S.E. Mechanism of Ca2+-dependent desensitization in TRP channels. Channels 2008, 2, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, R.; Zhang, X. Ca2+ regulation of TRP ion channels. Int. J. Mol. Sci. 2018, 19, 1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.X. Multiple roles of calmodulin and other Ca2+-binding proteins in the functional regulation of TRP channels. Pflugers Arch. 2005, 451, 105–115. [Google Scholar] [CrossRef]
- Hughes, T.E.T.; Pumroy, R.A.; Yazici, A.T.; Kasimova, M.A.; Fluck, E.C.; Huynh, K.W.; Samanta, A.; Molugu, S.K.; Zhou, Z.H.; Carnevale, V.; et al. Structural insights on TRPV5 gating by endogenous modulators. Nat. Commun. 2018, 9, 4198. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.Y.; Procko, E.; Gaudet, R. Distinct properties of Ca2+-calmodulin binding to N- and C-terminal regulatory regions of the TRPV1 channel. J. Gen. Physiol. 2012, 140, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Flores, C.M. Polypeptide Complex of TRPM8 and Calmodulin and Its Uses Thereof. U.S. Patent 2007/0105155 A1, 10 May 2007. [Google Scholar]
- Hasan, R.; Leeson-Payne, A.T.S.; Jaggar, J.H.; Zhang, X. Calmodulin is responsible for Ca2+-dependent regulation of TRPA1 Channels. Sci. Rep. 2017, 7, 45098. [Google Scholar] [CrossRef] [Green Version]
- Diver, M.M.; Cheng, Y.; Julius, D. Structural insights into TRPM8 inhibition and desensitization. Science 2019, 6672, eaax6672. [Google Scholar] [CrossRef]
- Autzen, H.E.; Myasnikov, A.G.; Campbell, M.G.; Asarnow, D.; Julius, D.; Cheng, Y. Structure of the human TRPM4 ion channel in a lipid nanodisc. Science 2018, 359, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faouzi, M.; Penner, R. TRPM2. Handb. Exp. Pharmacol. 2014, 222, 403–426. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Chubanov, V.; Gudermann, T.; Montell, C. TRPM5 is a voltage-modulated and Ca2+-activated monovalent selective cation channel. Curr. Biol. 2003, 13, 1153–1158. [Google Scholar] [CrossRef] [Green Version]
- Morales-Lázaro, S.L.; Lemus, L.; Rosenbaum, T. Regulation of thermoTRPs by lipids. Temperature 2017, 4, 24–40. [Google Scholar] [CrossRef]
- van de Graaf, S.F.; Boullart, I.; Hoenderop, J.G.; Bindels, R.J. Regulation of the epithelial Ca2+ channels TRPV5 and TRPV6 by 1alpha,25-dihydroxy Vitamin D3 and dietary Ca2+. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 303–308. [Google Scholar] [CrossRef]
- Morenilla-Palao, C.; Pertusa, M.; Meseguer, V.; Cabedo, H.; Viana, F. Lipid raft segregation modulates TRPM8 channel activity. J. Biol. Chem. 2009, 284, 9215–9224. [Google Scholar] [CrossRef] [Green Version]
- Startek, J.B.; Boonen, B.; López-Requena, A.; Talavera, A.; Alpizar, Y.A.; Ghosh, D.; Ranst, N.V.; Nilius, B.; Voets, T.; Talavera, K. Mouse TRPA1 function and membrane localization are modulated by direct interactions with cholesterol. eLife 2019, 8, e46084. [Google Scholar] [CrossRef]
- Huynh, K.W.; Cohen, M.R.; Jiang, J.; Samanta, A.; Lodowski, D.T.; Zhou, Z.H.; Moiseenkova-Bell, V.Y. Structure of the full-length TRPV2 channel by cryo-EM. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Li, J.; Zeng, B.; Chen, G.L.; Peng, X.; Zhang, Y.; Wang, J.; Clapham, D.E.; Li, Z.; Zhang, J. Structure of the mouse TRPC4 ion channel. Nat. Commun. 2018, 9, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Prevarskaya, N.; Zhang, L.; Barritt, G. TRP channels in cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 937–946. [Google Scholar] [CrossRef] [Green Version]
- So, C.L.; Milevskiy, M.J.G.; Monteith, G.R. Transient receptor potential cation channel subfamily V and breast cancer. Lab. Investig. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chodon, D.; Guilbert, A.; Dhennin-Duthille, I.; Gautier, M.; Telliez, M.-s.; Sevestre, H.; Ouadid-Ahidouch, H. Estrogen regulation of TRPM8 expression in breast cancer cells. BMC Cancer 2010, 10, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jall, S.; Finan, B.; Collden, G.; Fischer, K.; Dong, X.; Tschöp, M.H.; Müller, T.D.; Clemmensen, C. Pirt deficiency has subtle female-specific effects on energy and glucose metabolism in mice. Mol. Metab. 2019, 23, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Hille, B.; Dickson, E.J.; Kruse, M.; Vivas, O.; Suh, B.C. Phosphoinositides regulate ion channels. Biochim. Biophys. Acta 2015, 1851, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Kim, A.; Masuch, T.; Park, K.; Weng, H.; Wetzel, C.; Dong, X. Pirt functions as an endogenous regulator of TRPM8. Nat. Commun. 2013, 4, 2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Wu, G.Y.; Dong, X.Z.; Tang, Z.X. Phosphoinositide interacting regulator of TRP (Pirt) enhances TRPM8 channel activity in vitro via increasing channel conductance. Acta Pharmacol. Sin. 2016, 37, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Joo, K.W.; Jeon, U.S.; Kim, G.H.; Park, J.; Oh, Y.K.; Kim, Y.S.; Ahn, C.; Kim, S.; Kim, S.Y.; Lee, J.S.; et al. Antidiuretic action of oxytocin is associated with increased urinary excretion of aquaporin-2. Nephrol. Dial. Transplant 2004, 19, 2480–2486. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S. Is oxytocin a player in antidiuresis? J. Am. Soc. Nephrol. 2008, 19, 189–190. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Kd (M) | RMSE (M) | Ligand Type |
|---|---|---|---|
| Calmodulin (apo, free) | 350 × 10−9 | 40 × 10−9 | CaM, Intracellular Protein |
| Calmodulin (holo, bound) | 60 × 10−6 | 30 × 10−6 | Calcium Bound CaM |
| Cholesteryl HS | 103 × 10−6 | 6 × 10−6 | Steroid Hormone Precursor |
| Cortisol | 790 × 10−6 | 50 × 10−6 | Cortical Steroid Hormone |
| β-Estradiol | 800 × 10−6 | 100 × 10−6 | Sex Steroid Hormone |
| Cholecalciferol | 2.1 × 10−3 | 0.4 × 10−3 | Secosteroid Hormone |
| Oxytocin | 7 × 10−6 | 1 × 10−6 | Peptide Hormone |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sisco, N.J.; Luu, D.D.; Kim, M.; Van Horn, W.D. PIRT the TRP Channel Regulating Protein Binds Calmodulin and Cholesterol-Like Ligands. Biomolecules 2020, 10, 478. https://doi.org/10.3390/biom10030478
Sisco NJ, Luu DD, Kim M, Van Horn WD. PIRT the TRP Channel Regulating Protein Binds Calmodulin and Cholesterol-Like Ligands. Biomolecules. 2020; 10(3):478. https://doi.org/10.3390/biom10030478
Chicago/Turabian StyleSisco, Nicholas J., Dustin D. Luu, Minjoo Kim, and Wade D. Van Horn. 2020. "PIRT the TRP Channel Regulating Protein Binds Calmodulin and Cholesterol-Like Ligands" Biomolecules 10, no. 3: 478. https://doi.org/10.3390/biom10030478
APA StyleSisco, N. J., Luu, D. D., Kim, M., & Van Horn, W. D. (2020). PIRT the TRP Channel Regulating Protein Binds Calmodulin and Cholesterol-Like Ligands. Biomolecules, 10(3), 478. https://doi.org/10.3390/biom10030478