Selenium and Selenoproteins in Adipose Tissue Physiology and Obesity

 , ,
, , 
 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. A Brief Introduction into Selenoprotein Function
3. Se Status in Obesity
3.1. Se Blood Levels and Intake Patterns
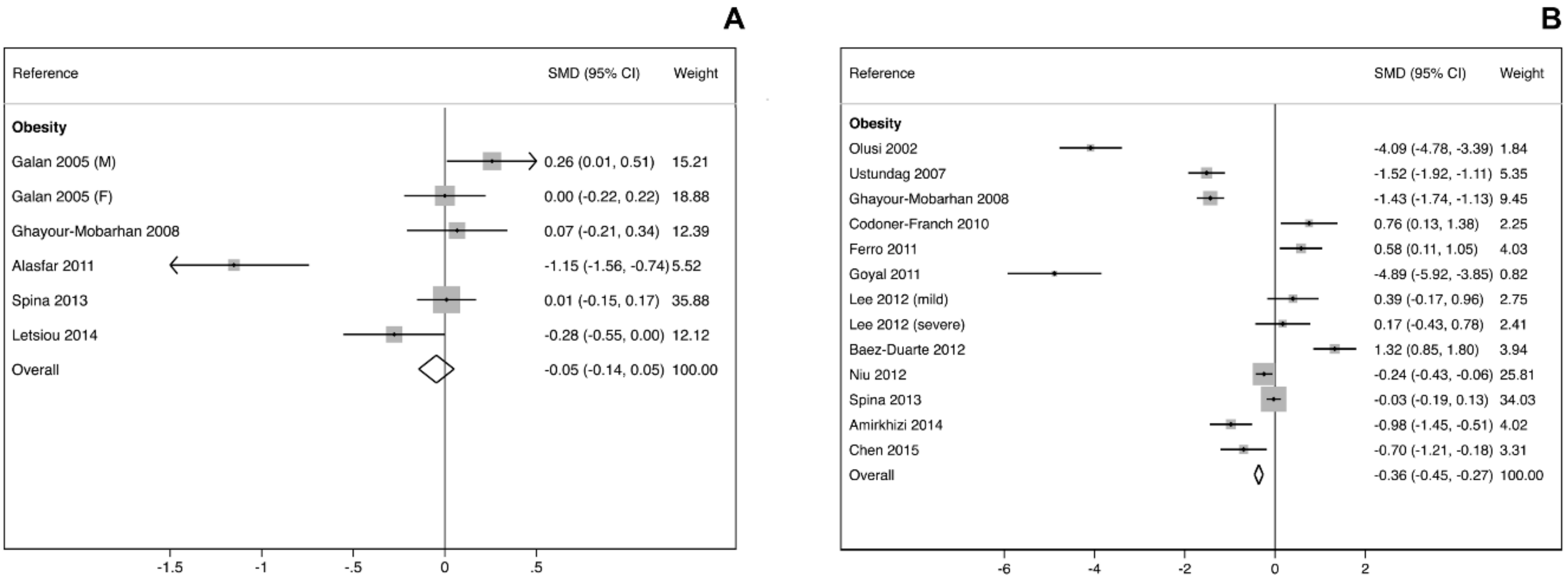
3.2. GPX Activity and Levels
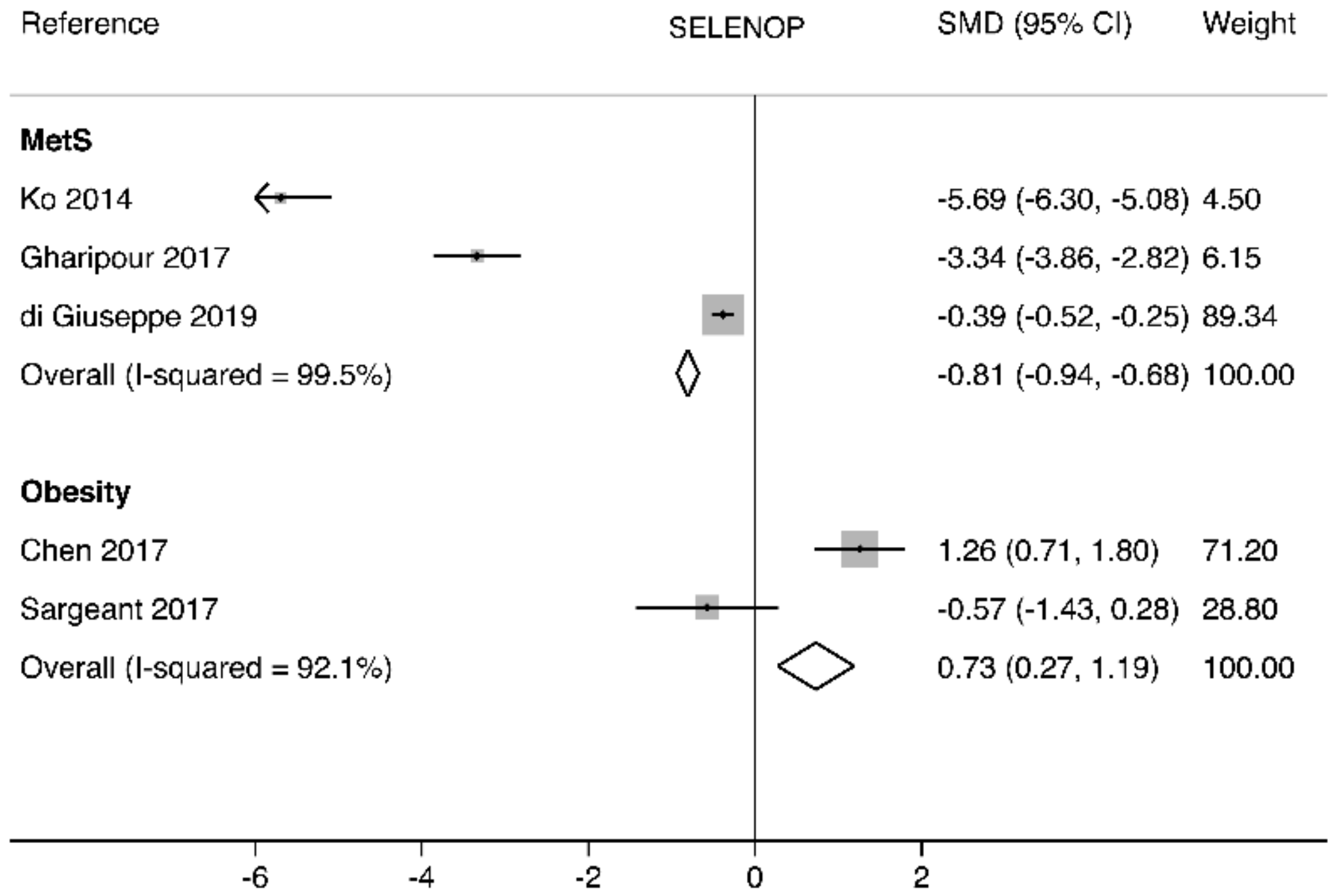
3.3. Circulating SELENOP Levels
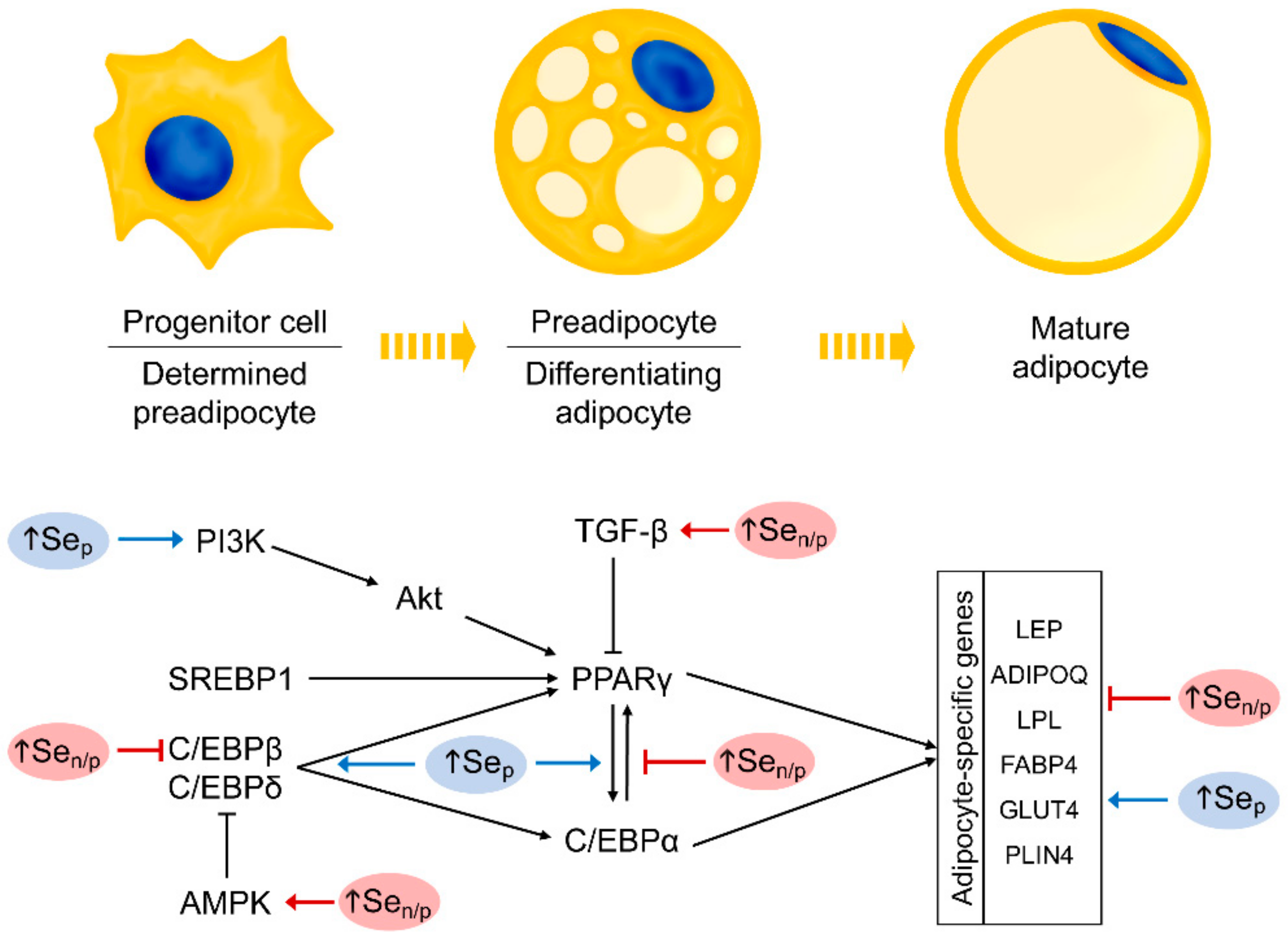
4. Se in Adipogenesis and Adipocyte Signaling Pathways
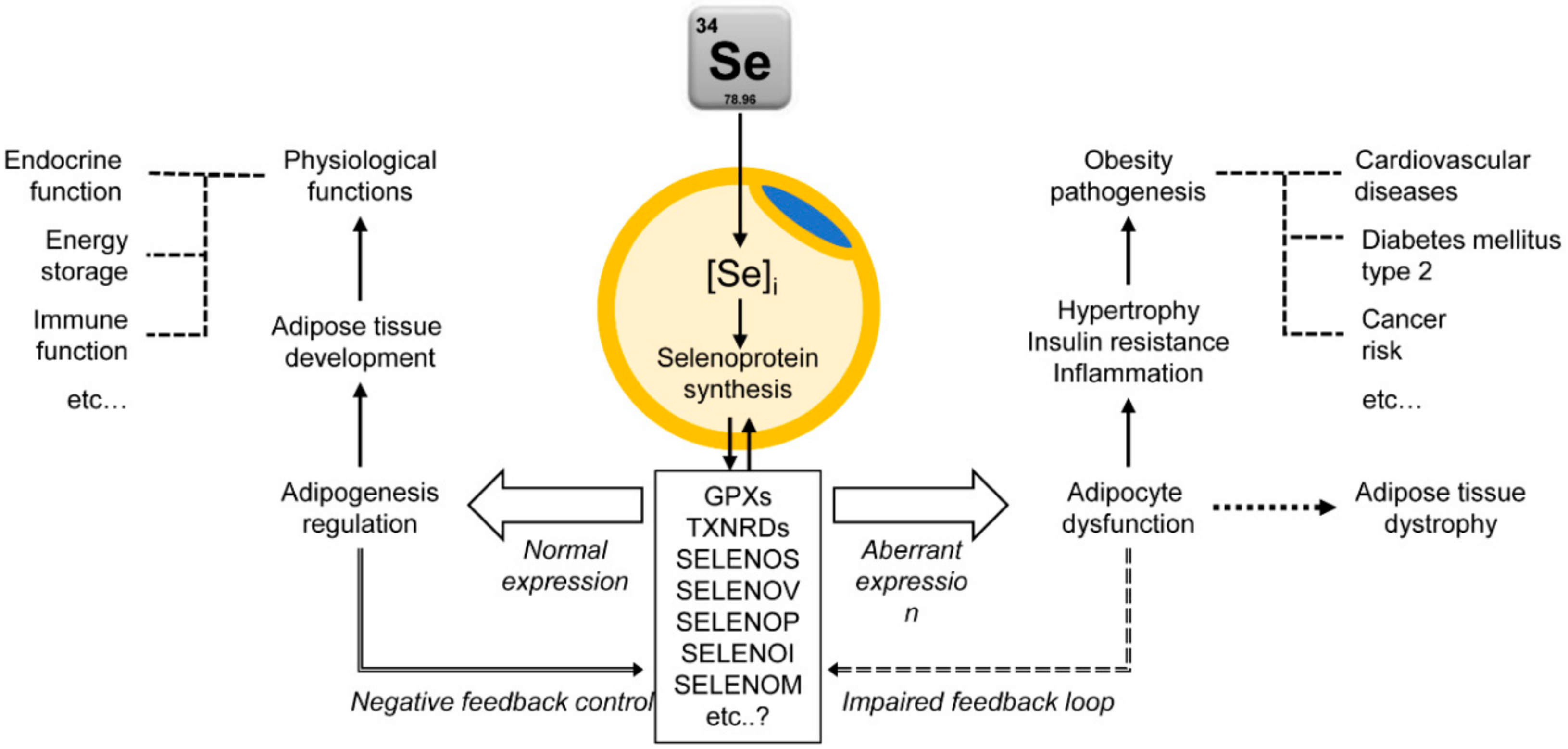
5. Adipose Tissue as a Target for Se Activity: Adipocyte Selenoproteins
5.1. Selenoprotein P (SELENOP)
5.2. Selenoprotein S (SELENOS)
5.3. Glutathione Peroxidases (GPXs)
5.4. Thioredoxin Reductases (TXNRDs)
5.5. Deiodinases (DIOs)
5.6. Other Selenoproteins
5.7. Proteins Involved in Selenoprotein Synthesis and Degradation
6. Selenium and Brown Adipose Tissue (BAT)
7. Se in Central Control of Food Intake
8. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Brigelius-Flohé, R.; Arnér, E. Selenium and selenoproteins in (redox) signaling, diseases, and animal models-200 year anniversary issue. Free Radic Biol. Med. 2018, 127, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Papp, L.V.; Holmgren, A.; Khanna, K.K. Selenium and selenoproteins in health and disease. Antioxid. Redox Signal. 2010, 12, 793–795. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, B. Redox-active selenium in health and disease: A conceptual review. Mini Rev. Med. Chem. 2019, 19, 720–726. [Google Scholar] [CrossRef]
- Kasaikina, M.V.; Hatfield, D.L.; Gladyshev, V.N. Understanding selenoprotein function and regulation through the use of rodent models. BBA Mol. Cell Res. 2012, 1823, 1633–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, J.; Hoffmann, P. Selenium, selenoproteins, and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
- Köhrle, J. Selenium and endocrine tissues. In Selenium; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: Cham, Switzerland, 2016; pp. 389–400. [Google Scholar]
- Sneddon, A.A. Selenium and vascular health. Pure Appl. Chem. 2011, 84, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Mistry, H.D.; Pipkin, F.B.; Redman, C.W.; Poston, L. Selenium in reproductive health. Am. J. Obstet. Gynecol. 2012, 206, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Steinbrenner, H.; Sies, H. Selenium homeostasis and antioxidant selenoproteins in brain: Implications for disorders in the central nervous system. Arch. Biochem. Biophys. 2013, 536, 152–157. [Google Scholar] [CrossRef]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Sun, S.; Feng, H.; Hou, J. Keshan disease. In Endemic Disease in China; Sun, D., Ed.; Springer: Singapore, 2019; pp. 175–221. [Google Scholar]
- Wang, K.; Yu, J.; Liu, H.; Liu, Y.; Liu, N.; Cao, Y.; Sun, D. Endemic Kashin–Beck disease: A food-sourced osteoarthropathy. In Seminars in Arthritis and Rheumatism; WB Saunders: Philadelphia, PA, USA, 2019. [Google Scholar]
- Alehagen, U.; Aaseth, J. Selenium and coenzyme Q10 interrelationship in cardiovascular diseases–a clinician’s point of view. J. Trace Elem. Med. Biol. 2015, 31, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Peters, K.M.; Galinn, S.E.; Tsuji, P.A. Selenium: Dietary sources, human nutritional requirements and intake across populations. In Selenium; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: Cham, Switzerland, 2016; pp. 295–305. [Google Scholar]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. FASEB J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayman, M.P. Selenium intake, status, and health: A complex relationship. Hormones 2019, 19, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.C.; Zhou, J.; Su, L.; Huang, K.; Lei, X.G. Selenium and diabetes. In Selenium; Michalke, B., Ed.; Springer: Cham, Switzerland, 2018; pp. 317–344. [Google Scholar]
- Zhang, X.; Li, X.; Zhang, W.; Song, Y. Selenium and cardiovascular disease: Epidemiological evidence of a possible U-shaped relationship. In Selenium; Michalke, B., Ed.; Springer: Cham, Switzerland, 2018; pp. 303–316. [Google Scholar]
- Vinceti, M.; Mandrioli, J.; Borella, P.; Michalke, B.; Tsatsakis, A.; Finkelstein, Y. Selenium neurotoxicity in humans: Bridging laboratory and epidemiologic studies. Toxicol. Lett. 2014, 230, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Vinceti, M.; Filippini, T.; Rothman, K.J. Selenium exposure and the risk of type 2 diabetes: A systematic review and meta-analysis. Eur. J. Epidemiol. 2018, 33, 789–810. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H. Interference of selenium and selenoproteins with the insulin-regulated carbohydrate and lipid metabolism. Free Radic. Biol. Med. 2013, 65, 1538–1547. [Google Scholar] [CrossRef]
- Zeng, M.-S.; Li, X.; Liu, Y.; Zhao, H.; Zhou, J.-C.; Li, K.; Huang, J.-Q.; Sun, L.-H.; Tang, J.-Y.; Xia, X.-J.; et al. A high-selenium diet induced insulin resistance in gestating rats and their offspring. Free Radic. Biol. Med. 2012, 52, 1335–1342. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, H.; Zhang, J.Q.-S.; Tang, Y.; Li, K.; Xia, X.-J.; Wang, K.-N.; Li, K.; Lei, X.G. Prolonged dietary Se deficiency or excess does not globally affect selenoprotein gene expression and(or) protein production in various tissues of pigs. J. Nutr. 2012, 142, 1410–1416. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.P.; Barcus, M.; Kim, J.G.; Lum, K.; Mills, C.; Lei, X.G. High dietary selenium intake alters lipid metabolism and protein synthesis in liver and muscle of pigs. J. Nutr. 2016, 146, 1625–1633. [Google Scholar] [CrossRef]
- Mita, Y.; Nakayama, K.; Inari, S.; Nishito, Y.; Yoshioka, Y.; Sakai, N.; Iwasaki, M. Selenoprotein P-neutralizing antibodies improve insulin secretion and glucose sensitivity in type 2 diabetes mouse models. Nat. Commun. 2017, 8, 1–17. [Google Scholar] [CrossRef]
- Saito, Y. Selenoprotein P as an in vivo redox regulator: Disorders related to its deficiency and excess. J. Clin. Biochem. Nutr. 2019, 66, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, B.; Saedisomeolia, A.; Allman-Farinelli, M. Association between antioxidant intake/status and obesity: A systematic review of observational studies. Biol. Trace Elem. Res. 2017, 175, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Tajaddini, M.H.; Keikha, M.; Razzazzadeh, A.; Kelishadi, R. A systematic review on the association of serum selenium and metabolic syndrome. J. Res. Med. Sci. 2015, 20, 782. [Google Scholar] [PubMed]
- Tang, X.; Li, J.; Zhao, W.G.; Sun, H.; Guo, Z.; Jing, L.; Fu, Y. Comprehensive map and functional annotation of the mouse white adipose tissue proteome. Peer J. 2019, 7, e7352. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, K.; Tang, J.Y.; Zhou, J.C.; Wang, K.N.; Xia, X.J.; Lei, X.G. Expression of selenoprotein genes is affected by obesity of pigs fed a high-fat diet. Nutr. J. 2015, 145, 1394–1401. [Google Scholar] [CrossRef] [Green Version]
- Adani, G.; Filippini, T.; Michalke, B.; Vinceti, M. Selenium and Other Trace Elements in the Etiology of Parkinson’s Disease: A Systematic Review and Meta-Analysis of Case-Control Studies. Neuroepidemiology 2020, 54, 1–23. [Google Scholar] [CrossRef]
- Gladyshev, V.N.; Hatfield, D.L. Selenocysteine biosynthesis, selenoproteins, and selenoproteomes. In Recoding: Expansion of Decoding Rules Enriches Gene Expression; Atkins, J.F., Gesteland, R.F., Eds.; Springer: New York, NY, USA, 2010; pp. 3–27. [Google Scholar]
- Pitts, M.W.; Hoffmann, P.R. Endoplasmic reticulum-resident selenoproteins as regulators of calcium signaling and homeostasis. Cell Calcium 2018, 70, 76–86. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Speckmann, B.; Klotz, L.O. Selenoproteins: Antioxidant selenoenzymes and beyond. Arch. Biochem. Biophys. 2016, 595, 113–119. [Google Scholar] [CrossRef]
- Flohé, L.; Brigelius-Flohé, R. Basics and news on glutathione peroxidases. In Selenium; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: Cham, Switzerland, 2016; pp. 211–222. [Google Scholar]
- Brigelius-Flohé, R.; Flohé, L. Regulatory phenomena in the glutathione peroxidase superfamily. Antioxid. Redox Signal. 2019, in press. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Kim, H.Y. The methionine sulfoxide reduction system: Selenium utilization and methionine sulfoxide reductase enzymes and their functions. Antioxid. Redox Signal. 2013, 19, 958–969. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L. Selenium, selenoproteins and the thyroid gland: Interactions in health and disease. Nat. Rev. Endocrinol. 2012, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Tobe, R.; Mihara, H. Delivery of selenium to selenophosphate synthetase for selenoprotein biosynthesis. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2433–2440. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E.; Motley, A.K. Selenoprotein metabolism and function: Evidence for more than one function for selenoprotein P. J. Nutr. 2003, 133, 1517S–1520S. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Takahashi, K. Selenoprotein P. In Selenoproteins and Mimics; Liu, J., Luo, G., Mu, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 77–88. [Google Scholar]
- Almanza, A.; Carlesso, A.; Chintha, C.; Creedican, S.; Doultsinos, D.; Leuzzi, B.; Papaioannou, A. Endoplasmic reticulum stress signalling–from basic mechanisms to clinical applications. FEBS J. 2019, 286, 241–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, J.H.; Zhang, X.; Cheng, W.H. The thioredoxin-like family of selenoproteins: Implications in aging and age-related degeneration. Biol. Trace Elem. Res. 2019, 188, 189–195. [Google Scholar] [CrossRef]
- Han, S.J.; Lee, B.C.; Yim, S.H.; Gladyshev, V.N.; Lee, S.R. Characterization of mammalian selenoprotein o: A redox-active mitochondrial protein. PLoS ONE 2014, 9, e95518. [Google Scholar] [CrossRef] [Green Version]
- Horibata, Y.; Elpeleg, O.; Eran, A.; Hirabayashi, Y.; Savitzki, D.; Tal, G.; Sugimoto, H. EPT1 (selenoprotein I) is critical for the neural development and maintenance of plasmalogen in humans. J. Lipid Res. 2018, 59, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Ying, H.; Zhang, Y. Systems biology of selenium and complex disease. Biol. Trace Elem. Res. 2019, 192, 38–50. [Google Scholar] [CrossRef]
- Combs, F., Jr. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Q.; Lin, R.; Nong, Q. Adiposity and Serum Selenium in US Adults. Nutrients 2018, 10, 727. [Google Scholar] [CrossRef] [Green Version]
- Alasfar, F.; Ben-Nakhi, M.; Khoursheed, M.; Kehinde, E.O.; Alsaleh, M. Selenium is significantly depleted among morbidly obese female patients seeking bariatric surgery. Obes. Surg. 2011, 21, 1710–1713. [Google Scholar] [CrossRef] [PubMed]
- Ortega, R.M.; Rodríguez-Rodríguez, E.; Aparicio, A.; Jiménez-Ortega, A.I.; Palermos, C.; Perea, J.M.; López-Sobaler, A.M. Young children with excess of weight show an impaired selenium status. Int. J. Vitam. Nutr. Res. 2012, 82, 121. [Google Scholar] [CrossRef] [PubMed]
- Błażewicz, A.; Klatka, M.; Astel, A.; Korona-Glowniak, I.; Dolliver, W.; Szwerc, W.; Kocjan, R. Serum and urinary selenium levels in obese children: A cross-sectional study. J. Trace Elem. Med. Biol. 2015, 29, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Papamargaritis, D.; Aasheim, E.T.; Sampson, B.; le Roux, C.W. Copper, selenium and zinc levels after bariatric surgery in patients recommended to take multivitamin-mineral supplementation. J. Trace Elem. Med. Biol. 2015, 31, 167–172. [Google Scholar] [CrossRef]
- Lefebvre, P.; Letois, F.; Sultan, A.; Nocca, D.; Mura, T.; Galtier, F. Nutrient deficiencies in patients with obesity considering bariatric surgery: A cross-sectional study. Surg. Obes. Relat. Dis. 2014, 10, 540–546. [Google Scholar] [CrossRef]
- Galan, P.; Viteri, F.E.; Bertrais, S.; Czernichow, S.; Faure, H.; Arnaud, J.; Roussel, A.M. Serum concentrations of β-carotene, vitamins C and E, zinc and selenium are influenced by sex, age, diet, smoking status, alcohol consumption and corpulence in a general French adult population. Eur. J. Clin. Nutr. 2005, 59, 1181. [Google Scholar] [CrossRef] [Green Version]
- Letsiou, S.; Nomikos, T.; Panagiotakos, D.B.; Pergantis, S.A.; Fragopoulou, E.; Pitsavos, C.; Antonopoulou, S. Gender-specific distribution of selenium to serum selenoproteins: Associations with total selenium levels, age, smoking, body mass index, and physical activity. Biofactors 2014, 40, 524–535. [Google Scholar] [CrossRef]
- Fatani, S.H.; Saleh, S.A.; Adly, H.M.; Abdulkhaliq, A.A. Trace element alterations in the hair of diabetic and obese women. Biol. Trace Elem. Res. 2016, 174, 32–39. [Google Scholar] [CrossRef]
- Liu, A.; Xu, P.; Gong, C.; Zhu, Y.; Zhang, H.; Nie, W.; Zhou, X.; Liang, X.; Xu, Y.; Huang, C.; et al. High serum concentration of selenium, but not calcium, cobalt, copper, iron, and magnesium, increased the risk of both hyperglycemia and dyslipidemia in adults: A health examination center based cross-sectional study. J. Trace Elem. Med. Biol. 2020, 59, 126470. [Google Scholar] [CrossRef]
- Lu, C.W.; Chang, H.H.; Yang, K.C.; Kuo, C.S.; Lee, L.T.; Huang, K.C. High serum selenium levels are associated with increased risk for diabetes mellitus independent of central obesity and insulin resistance. BMJ Open Diabetes Res. Care 2016, 4, e000253. [Google Scholar] [CrossRef] [Green Version]
- Binti Othman, F.; bin Jan Mohamed, H.J.; Sirajudeen, K.N.S.; Rajab, N.F. The influence of selenium status on body composition, oxidative DNA damage and total antioxidant capacity in newly diagnosed type 2 diabetes mellitus: A case-control study. J. Trace Elem. Med. Biol. 2017, 43, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Oo, S.M.; Misu, H.; Saito, Y.; Tanaka, M.; Kato, S.; Kita, Y.; Nakagen, M. Serum selenoprotein P, but not selenium, predicts future hyperglycemia in a general Japanese population. Sci. Rep. 2018, 8, 16727. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, J.; De Lorgeril, M.; Akbaraly, T.; Salen, P.; Arnout, J.; Cappuccio, F.P.; Iacoviello, L. Gender differences in copper, zinc and selenium status in diabetic-free metabolic syndrome European population–The IMMIDIET study. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.W.; Chang, H.H.; Yang, K.C.; Chiang, C.H.; Yao, C.A.; Huang, K.C. Gender Differences with Dose–Response Relationship between Serum Selenium Levels and Metabolic Syndrome—A Case-Control Study. Nutrients 2019, 11, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutakin Meiliana, A.; Wijaya, A.; Kobayashi, K.; Yamazaki, C.; Kameo, S.; Nakazawa, M.; Koyama, H. Association between selenium nutritional status and metabolic risk factors in men with visceral obesity. J. Trace Elem. Med. Biol. 2013, 27, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, X.; Pedram, P.; Shahidi, M.; Du, J.; Yi, Y.; Sun, G. Significant beneficial association of high dietary selenium intake with reduced body fat in the CODING Study. Nutrients 2016, 8, 24. [Google Scholar] [CrossRef]
- Hermsdorff, H.H.M.; Zulet, M.Á.; Puchau, B.; Bressan, J.; Martínez, J.A. Association of retinol-binding protein-4 with dietary selenium intake and other lifestyle features in young healthy women. Nutrition 2009, 25, 392–399. [Google Scholar] [CrossRef]
- Duarte, G.B.S.; Reis, B.Z.; Rogero, M.M.; Vargas-Mendez, E.; Júnior, F.B.; Cercato, C.; Cozzolino, S.M.F. Consumption of Brazil nuts with high selenium levels increased inflammation biomarkers in obese women: A randomized controlled trial. Nutrition 2019, 63, 162–168. [Google Scholar] [CrossRef]
- Karaouzene, N.; Merzouk, H.; Aribi, M.; Merzouk, S.A.; Berrouiguet, A.Y.; Tessier, C.; Narce, M. Effects of the association of aging and obesity on lipids, lipoproteins and oxidative stress biomarkers: A comparison of older with young men. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 792–799. [Google Scholar] [CrossRef]
- Olusi, S.O. Obesity is an independent risk factor for plasma lipid peroxidation and depletion of erythrocyte cytoprotectic enzymes in humans. Int. J. Obes. 2002, 26, 1159. [Google Scholar] [CrossRef] [Green Version]
- Amirkhizi, F.; Siassi, F.; Djalali, M.; Shahraki, S.H. Impaired enzymatic antioxidant defense in erythrocytes of women with general and abdominal obesity. Obes. Res. Clin. Pract. 2014, 8, e26–e34. [Google Scholar] [CrossRef] [PubMed]
- Ghayour-Mobarhan, M.; Taylor, A.; Lanham-New, S.; Lamb, D.J.; Nezhad, M.A.; Kazemi-Bajestani, S.M.R.; Ferns, G.A. Serum selenium and glutathione peroxidase in patients with obesity and metabolic syndrome. Pak. J. Nutr. 2008, 7, 112–117. [Google Scholar] [CrossRef]
- Lee, S.D.; Ju, G.; Choi, J.A.; Kim, J.W.; Yoon, I.Y. The association of oxidative stress with central obesity in obstructive sleep apnea. Sleep Breath. 2012, 16, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Spina, A.; Guallar, E.; Rayman, M.P.; Tigbe, W.; Kandala, N.B.; Stranges, S. Anthropometric indices and selenium status in British adults: The UK National Diet and Nutrition Survey. Free Radic. Biol. Med. 2013, 65, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Ferro, F.E.D.; de Sousa Lima, V.B.; Soares, N.R.M.; de Sousa Almondes, K.G.; Pires, L.V.; Cozzolino, S.M.F.; do Nascimento Marreiro, D. Parameters of metabolic syndrome and its relationship with zincemia and activities of superoxide dismutase and glutathione peroxidase in obese women. Biol. Trace Elem. Res. 2011, 143, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Gunawardena, H.P.; Silva, K.D.R.R.; Sivakanesan, R.; Katulanda, P. Increased lipid peroxidation and erythrocyte glutathione peroxidase activity of patients with type 2 diabetes mellitus: Implications for obesity and central obesity. Obes. Med. 2019, 15, 100118. [Google Scholar] [CrossRef]
- Baez-Duarte, B.G.; Zamora-Ginez, I.; Mendoza-Carrera, F.; Ruiz-Vivanco, G.; Torres-Rasgado, E.; Gonzalez-Mejia, M.E.; Perez-Fuentes, R. Serum levels of glutathione peroxidase 3 in overweight and obese subjects from central Mexico. Arch. Med. Res. 2012, 43, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Tinahones, F.J.; Murri-Pierri, M.; Garrido-Sánchez, L.; García-Almeida, J.M.; García-Serrano, S.; García-Arnés, J.; García-Fuentes, E. Oxidative stress in severely obese persons is greater in those with insulin resistance. Obesity 2009, 17, 240–246. [Google Scholar] [CrossRef]
- Ustundag, B.; Gungor, S.; Aygün, A.D.; Turgut, M.; Yilmaz, E. Oxidative status and serum leptin levels in obese prepubertal children. Cell Biochem. Funct. Cell. Biochem. Modul. Act. Agents Dis. 2007, 25, 479–483. [Google Scholar] [CrossRef]
- Chen, S.; Sun, L.; Gao, H.; Ren, L.; Liu, N.; Song, G. Visfatin and oxidative stress influence endothelial progenitor cells in obese populations. Endocr. Res. 2015, 40, 83–87. [Google Scholar] [CrossRef]
- Codoñer-Franch, P.; Boix-García, L.; Simó-Jordá, R.; del Castillo-Villaescusa, C.; Maset-Maldonado, J.; Valls-Bellés, V. Is obesity associated with oxidative stress in children? Int. J. Pediatr. Obes. 2010, 5, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Sfar, S.; Boussoffara, R.; Sfar, M.T.; Kerkeni, A. Antioxidant enzymes activities in obese Tunisian children. Nutr. J. 2013, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Albuali, W.H. Evaluation of oxidant-antioxidant status in overweight and morbidly obese Saudi children. World J. Clin. Pediatr. 2014, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Ozgen, I.T.; Tascilar, M.E.; Bilir, P.; Boyraz, M.; Guncikan, M.N.; Akay, C.; Dundaroz, R. Oxidative stress in obese children and its relation with insulin resistance. J. Pediatr. Endocrinol. Meta 2012, 25, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Bougoulia, M.; Triantos, A.; Koliakos, G. Plasma interleukin-6 levels, glutathione peroxidase and isoprostane in obese women before and after weight loss. Association with cardiovascular risk factors. Horm. Athens 2006, 5, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bougoulia, M.; Triantos, A.; Koliakos, G. Effect of weight loss with or without orlistat treatment on adipocytokines, inflammation, and oxidative markers in obese women. Horm. Athens 2006, 5, 259. [Google Scholar] [CrossRef] [Green Version]
- Goyal, R.; Singhai, M.; Faizy, A.F. Glutathione peroxidase activity in obese and nonobese diabetic patients and role of hyperglycemia in oxidative stress. J. Mid Life Health 2011, 2, 72. [Google Scholar]
- López-Domènech, S.; Abad-Jiménez, Z.; Iannantuoni, F.; de Marañón, A.M.; Rovira-Llopis, S.; Morillas, C.; Rocha, M. Moderate weight loss attenuates chronic endoplasmic reticulum stress and mitochondrial dysfunction in human obesity. Mol. Metab. 2019, 19, 24–33. [Google Scholar] [CrossRef]
- Feng, R.N.; Niu, Y.C.; Sun, X.W.; Li, Q.; Zhao, C.; Wang, C.; Li, Y. Histidine supplementation improves insulin resistance through suppressed inflammation in obese women with the metabolic syndrome: A randomised controlled trial. Diabetologia 2013, 56, 985–994. [Google Scholar] [CrossRef]
- Niu, Y.C.; Feng, R.N.; Hou, Y.; Li, K.; Kang, Z.; Wang, J.; Li, Y. Histidine and arginine are associated with inflammation and oxidative stress in obese women. Br. J. Nutr. 2012, 108, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Kuzuya, M.; Ando, F.; Iguchi, A.; Shimokata, H. Glutathione peroxidase 1 Pro198Leu variant contributes to the metabolic syndrome in men in a large Japanese cohort. Am. J. Clin. Nutr. 2008, 87, 1939–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, C.H.; Chávez, P.H.; Castro, N.M.; Carriedo, A.P.; Del Río, S.G.; Lizaur, A.P. Glutathione peroxidase-1 Pro200Leu polymorphism (rs1050450) is associated with morbid obesity independently of the presence of prediabetes or diabetes in women from Central Mexico. Nutr. Hosp. 2015, 32, 1516–1525. [Google Scholar]
- Rupérez, A.I.; Olza, J.; Gil-Campos, M.; Leis, R.; Mesa, M.D.; Tojo, R.; Aguilera, C.M. Association of genetic polymorphisms for glutathione peroxidase genes with obesity in Spanish children. Lifestyle Genom. 2014, 7, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Cominetti, C.; de Bortoli, M.C.; Purgatto, E.; Ong, T.P.; Moreno, F.S.; Garrido, A.B., Jr.; Cozzolino, S.M.F. Associations between glutathione peroxidase-1 Pro198Leu polymorphism, selenium status, and DNA damage levels in obese women after consumption of Brazil nuts. Nutrition 2011, 27, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E. Selenoprotein P—Expression, functions, and roles in mammals. BBA Gen. Subj. 2009, 1790, 1441–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D Giuseppe, R.; Koch, M.; Schlesinger, S.; Borggrefe, J.; Both, M.; Müller, H.P.; Lieb, W. Circulating selenoprotein P levels in relation to MRI-derived body fat volumes, liver fat content, and metabolic disorders. Obesity 2017, 25, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Gharipour, M.; Sadeghi, M.; Salehi, M.; Behmanesh, M.; Khosravi, E.; Dianatkhah, M.; Gharipour, A. Association of expression of selenoprotein P in mRNA and protein levels with metabolic syndrome in subjects with cardiovascular disease: Results of the Selenegene study. J. Gene Med. 2017, 19, e2945. [Google Scholar] [CrossRef]
- Ko, B.J.; Kim, S.M.; Park, K.H.; Park, H.S.; Mantzoros, C.S. Levels of circulating selenoprotein P, fibroblast growth factor (FGF) 21 and FGF23 in relation to the metabolic syndrome in young children. Int. J. Obes. 2014, 38, 1497. [Google Scholar] [CrossRef]
- Lim, J.; Park, H.S.; Lee, S.K.; Jang, Y.J.; Lee, Y.J.; Heo, Y. Longitudinal changes in serum levels of angiopoietin-like protein 6 and selenoprotein P after gastric bypass surgery. Obes. Surg. 2016, 26, 825–832. [Google Scholar] [CrossRef]
- di Giuseppe, R.; Plachta-Danielzik, S.; Koch, M.; Nöthlings, U.; Schlesinger, S.; Borggrefe, J.; Lieb, W. Dietary pattern associated with selenoprotein P and MRI-derived body fat volumes, liver signal intensity, and metabolic disorders. Eur. J. Nutr. 2019, 58, 1067–1079. [Google Scholar] [CrossRef]
- Choi, H.Y.; Hwang, S.Y.; Lee, C.H.; Hong, H.C.; Yang, S.J.; Yoo, H.J.; Choi, D.S. Increased selenoprotein p levels in subjects with visceral obesity and nonalcoholic Fatty liver disease. Diabetes Metab. 2013, 37, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Liu, B.; Wilkinson, D.; Hutchison, A.T.; Thompson, C.H.; Wittert, G.A.; Heilbronn, L.K. Selenoprotein P is elevated in individuals with obesity, but is not independently associated with insulin resistance. Obes. Res. Clin. Pract. 2017, 11, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, J.A.; Aithal, G.P.; Takamura, T.; Misu, H.; Takayama, H.; Douglas, J.A.; Yates, T. The influence of adiposity and acute exercise on circulating hepatokines in normal-weight and overweight/obese men. Appl. Physiol. Nutr. Metab. 2017, 43, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired adipogenesis and dysfunctional adipose tissue in human hypertrophic obesity. Physiol. Rev. 2018, 98, 1911–1941. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, J.H.; Nam, S.W.; Kim, B.W.; Kim, G.Y.; Kim, W.J.; Choi, Y.H. Selenium improves stem cell potency by stimulating the proliferation and active migration of 3T3-L1 preadipocytes. Int. J. Oncol. 2014, 44, 336–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, A.; Ahn, J.; Suh, Y.; Choi, Y.M.; Chen, P.; Lee, K. Selenium promotes adipogenic determination and differentiation of chicken embryonic fibroblasts with regulation of genes involved in fatty acid uptake, triacylglycerol synthesis and lipolysis. J. Nutr. Biochem. 2014, 25, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.O.; Kim, M.M.; Park, S.J.; Kim, D.; Chung, J.; Chung, A.S. Selenite suppresses hydrogen peroxide-induced cell apoptosis through inhibition of ASK1/JNK and activation of PI3-K/Akt pathways. FASEB J. 2002, 16, 111–113. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Hwang, J.T.; Lee, Y.K.; Kim, Y.M.; Park, O.J. AMP-activated kinase regulates adipocyte differentiation process in 3T3-L1 adipocytes treated with selenium. J. Life Sci. 2009, 19, 423–428. [Google Scholar]
- Shon, M.S.; Song, J.H.; Kim, G.N. Anti-obese Function of Selenate, An Essential Micronutrient, by Regulation of Adipogenesis in C3H10T1/2 Cells. Korean J. Aesthet. Cosmetol. 2013, 11, 447–452. [Google Scholar]
- Kim, C.; Kim, K.H. Selenate Prevents Adipogenesis through Induction of Selenoprotein S and Attenuation of Endoplasmic Reticulum Stress. Molecules 2018, 23, 2882. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.Y.; Kim, G.N.; Wiacek, J.L.; Chen, C.Y.; Kim, K.H. Selenate inhibits adipogenesis through induction of transforming growth factor-β1 (TGF-β1) signaling. Biochem. Biophys. Res. Commun. 2012, 426, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Wiacek, J.L.; Kim, K.H. Sodium selenate inhibits adipogenesis in vitro. FASEB J. 2010, 24, 547–548. [Google Scholar]
- Suh, N.; Lee, E.B. Antioxidant effects of selenocysteine on replicative senescence in human adipose-derived mesenchymal stem cells. BMB Rep. 2017, 50, 572. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Zhu, Y.; Buhman, K.K.; Kim, K.H. Dietary selenate attenuates adiposity and improves insulin sensitivity in high-fat diet-induced obese mice. J. Funct. Foods 2015, 17, 33–42. [Google Scholar] [CrossRef]
- Nido, S.A.; Shituleni, S.A.; Mengistu, B.M.; Liu, Y.; Khan, A.Z.; Gan, F.; Huang, K. Effects of selenium-enriched probiotics on lipid metabolism, antioxidative status, histopathological lesions, and related gene expression in mice fed a high-fat diet. Biol. Trace Elem. Res. 2016, 171, 399–409. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, F.; Yang, H.; Chen, J.; Ren, Y.; Yuan, Z.; Wang, Y. Selenium-enriched exopolysaccharides produced by Enterobacter cloacae Z0206 alleviate adipose inflammation in diabetic KKAy mice through the AMPK/SirT1 pathway. Mol. Med. Rep. 2014, 9, 683–688. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Choi, S.I.; Lee, H.R.; Hwang, I.S.; Lee, Y.J.; An, B.S.; Hwang, D.Y. Selenium significantly inhibits adipocyte hypertrophy and abdominal fat accumulation in OLETF rats via induction of fatty acid β-oxidation. Biol. Trace Elem. Res. 2012, 150, 360–370. [Google Scholar] [CrossRef]
- Wang, X.; Wu, H.; Long, Z.; Sun, Q.; Liu, J.; Liu, Y.; Hai, C. Differential effect of Se on insulin resistance: Regulation of adipogenesis and lipolysis. Mol. Cell. Biochem. 2016, 415, 89–102. [Google Scholar] [CrossRef]
- Zeng, H. Selenium as an essential micronutrient: Roles in cell cycle and apoptosis. Molecules 2009, 14, 1263–1278. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Navarrete, J.M.; Fernández-Real, J.M. Adipocyte differentiation. In Adipose Tissue Biology; Symonds, M.E., Ed.; Springer: Cham, Switzerland, 2017; pp. 69–90. [Google Scholar]
- Donma, M.M.; Donma, O. Promising link between selenium and peroxisome proliferator activated receptor gamma in the treatment protocols of obesity as well as depression. Med. Hypotheses 2016, 89, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, O. The insulin-like effects of selenate in rat adipocytes. J. Biol. Chem. 1990, 265, 1124–1128. [Google Scholar] [PubMed]
- Stapleton, S.R.; Garlock, G.L.; Foellmi-Adams, L.; Kletzien, R.F. Selenium: Potent stimulator of tyrosyl phosphorylation and activator of MAP kinase. BBA Mol. Cell Res. 1997, 1355, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Heart, E.; Sung, C.K. Insulin-like and non-insulin-like selenium actions in 3T3-L1 adipocytes. J. Cell. Biochem. 2003, 88, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Hei, Y.J.; Farahbakhshian, S.; Chen, X.; Battell, M.L.; McNeill, J.H. Stimulation of MAP kinase and S6 kinase by vanadium and selenium in rat adipocytes. Mol. Cell. Biochem. 1998, 178, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Ueki, H.; Ohkura, Y.; Motoyashiki, T.; Tominaga, N.; Mority, T. Increase in lipoprotein lipase activity in isolated rat adipose tissue by selenate. Biol. Pharm. Bull. 1993, 16, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Pinto, A.; Juniper, D.T.; Sanil, M.; Morgan, L.; Clark, L.; Sies, H.; Steinbrenner, H. Supranutritional selenium induces alterations in molecular targets related to energy metabolism in skeletal muscle and visceral adipose tissue of pigs. J. Inorg. Biochem. 2012, 114, 47–54. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, W.; Chen, H.; Liao, N.; Wang, Z.; Zhang, X.; Hai, C. High selenium impairs hepatic insulin sensitivity through opposite regulation of ROS. Toxicol. Lett. 2014, 224, 16–23. [Google Scholar] [CrossRef]
- Ullrey, D.E. Biochemical and physiological indicators of selenium status in animals. J. Lipid Res. 1987, 65, 1712–1726. [Google Scholar] [CrossRef]
- Tinkov, A.A.; Gatiatulina, E.R.; Popova, E.V.; Polyakova, V.S.; Skalnaya, A.A.; Agletdinov, E.F.; Skalny, A.V. Early high-fat feeding induces alteration of trace element content in tissues of juvenile male Wistar rats. Biol. Trace Elem. Res. 2017, 175, 367–374. [Google Scholar] [CrossRef]
- Prigol, M.; Brüning, C.A.; Martini, F.; Nogueira, C.W. Comparative excretion and tissue distribution of selenium in mice and rats following treatment with diphenyl diselenide. Biol. Trace Elem. Res. 2012, 150, 272–277. [Google Scholar] [CrossRef]
- Liang, Y.; Lin, S.L.; Wang, C.W.; Yao, H.D.; Zhang, Z.W.; Xu, S.W. Effect of selenium on selenoprotein expression in the adipose tissue of chickens. Biol. Trace Elem. Res. 2014, 160, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.; Marsicano, J.R.; Copeland, P.R. Uptake and utilization of selenium from selenoprotein P. Biol. Trace Elem. Res. 2018, 181, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, S.; Berry, M.J. Selenium. Role of the essential metalloid in health. In Interrelations between Essential Metal Ions and Human Diseases; Sigel, A., Sigel, H., Sigel, R.K.O., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 499–534. [Google Scholar]
- Yiannikouris, F.; Wang, Y.; Shoemaker, R.; Larian, N.; Thompson, J.; English, V.L.; Cassis, L.A. Deficiency of angiotensinogen in hepatocytes markedly decreases blood pressure in lean and obese male mice. Hypertension 2015, 66, 836–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezas, F.; Lagos, J.; Céspedes, C.; Vio, C.P.; Bronfman, M.; Marzolo, M.P. Megalin/LRP2 expression is induced by peroxisome proliferator-activated receptor-alpha and-gamma: Implications for PPARs’ roles in renal function. PLoS ONE 2011, 6, e16794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, B.; Farach-Carson, M.C.; Kenaley, E.; Akanbi, K.A. High extracellular calcium attenuates adipogenesis in 3T3-L1 preadipocytes. Exp. Cell Res. 2004, 301, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Abboud, M.; Gordon-Thomson, C.; Hoy, A.J.; Balaban, S.; Rybchyn, M.S.; Cole, L.; Mason, R.S. Uptake of 25-hydroxyvitamin D by muscle and fat cells. J. Steroid Biochem. Mol. Biol. 2014, 144, 232–236. [Google Scholar] [CrossRef]
- Urs, S.; Smith, C.; Campbell, B.; Saxton, A.M.; Taylor, J.; Zhang, B.; Moustaid-Moussa, N. Gene expression profiling in human preadipocytes and adipocytes by microarray analysis. J. Nutr. 2004, 134, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Liu, H.; Pan, Z.; Ding, F.; Kou, J.; Li, L.; Wang, J. The cloning, characterization, and expression profiling of the LRP8 gene in duck (Anas platyrhynchos). Mol. Cell. Biochem. 2013, 375, 139–149. [Google Scholar] [CrossRef]
- Reddy, S.S.; Connor, T.E.; Weeber, E.J.; Rebeck, W. Similarities and differences in structure, expression, and functions of VLDLR and ApoER2. Mol. Neurodegener. 2011, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Steinbrenner, H.; Micoogullari, M.; Hoang, N.A.; Bergheim, I.; Klotz, L.O.; Sies, H. Selenium-binding protein 1 (SELENBP1) is a marker of mature adipocytes. Redox Biol. 2019, 20, 489–495. [Google Scholar] [CrossRef]
- Peinado, J.R.; Pardo, M.; de la Rosa, O.; Malagón, M.M. Proteomic characterization of adipose tissue constituents, a necessary step for understanding adipose tissue complexity. Proteomics 2012, 12, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Abuelreich, S.; Alkeraishan, N.; Shwish, N.B.; Hamam, R.; Kassem, M.; Alajez, N.M. Multiple intracellular signaling pathways orchestrate adipocytic differentiation of human bone marrow stromal stem cells. Biosci. Rep. 2018, 38, BSR20171252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hida, K.; Wada, J.; Zhang, H.; Hiragushi, K.; Tsuchiyama, Y.; Shikata, K.; Makino, H. Identification of genes specifically expressed in the accumulated visceral adipose tissue of OLETF rats. J. Lipid Res. 2000, 41, 1615–1622. [Google Scholar] [PubMed]
- Zhang, Y.; Chen, X. Reducing selenoprotein P expression suppresses adipocyte differentiation as a result of increased preadipocyte inflammation. Am. J. Physiol. Endocrinol. Metab. 2010, 300, E77–E85. [Google Scholar] [CrossRef] [Green Version]
- Misu, H.; Takamura, T.; Takayama, H.; Hayashi, H.; Matsuzawa-Nagata, N.; Kurita, S.; Sakurai, M. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab. 2010, 12, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Kuroiwa, N.; Iwase, K.; Morii, S.; Takiguchi, M.; Hiwasa, T. Up-regulation of growth-inhibitory or tumor suppressive genes by overexpression of C/EBPα or C/EBPβ in ras-transformed NIH3T3 cells. Integr. Mol. Med. 2015, 2, 150–157. [Google Scholar]
- Park, S.H.; Moon, Y.S. Effects of Selenate on Adipocyte Differentiation and the Expression of Selenoproteins in 3T3-L1 Cells. J. Life Sci. 2014, 24, 1085–1091. [Google Scholar] [CrossRef] [Green Version]
- Niersmann, C.; Hauck, S.M.; Kannenberg, J.M.; Röhrig, K.; von Toerne, C.; Roden, M.; Carstensen-Kirberg, M. Omentin-regulated proteins combine a pro-inflammatory phenotype with an anti-inflammatory counterregulation in human adipocytes: A proteomics analysis. Diabetes Metab. Res. 2019, 35, e3074. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X. Adipose Expression and Regulation of Selenoprotein P in Obesity and Insulin Resistance. FASEB J. 2009, 23, 990.13. [Google Scholar]
- Yin, L.; Cai, W.; Sheng, J.; Sun, Y. Hypoxia induced changes of SePP1 expression in rat preadipocytes and its impact on vascular fibroblasts. Int. J. Clin. Exp. Med. 2014, 7, 41. [Google Scholar]
- Liang, C.P.; Tall, A.R. Transcriptional profiling reveals global defects in energy metabolism, lipoprotein, and bile acid synthesis and transport with reversal by leptin treatment in ob/ob mouse liver. J. Biol. Chem. 2001, 276, 49066–49076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, J.P.; Grune, T.; Speckmann, B. The two faces of reactive oxygen species (ROS) in adipocyte function and dysfunction. Biol. Chem. 2016, 397, 709–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.Y.; Kim, K.H. Dexamethasone-induced selenoprotein S degradation is required for adipogenesis. J. Lipid Res. 2013, 54, 2069–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Men, L.; Yu, S.; Yao, J.; Li, Y.; Ren, D.; Du, J. Selenoprotein S protects against adipocyte death through mediation of the IRE1α-sXBP1 pathway. Biochem. Biophys. Res. Commun. 2018, 503, 2866–2871. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Wiacek, J.L.; Kim, G.N.; Kim, K.H. Selenium inhibits adipogenesis through suppression of ER-stress and induction of selenoprotein S. FASEB J. 2011, 23, 2882. [Google Scholar]
- Ha, H.Y.; Alfulaij, N.; Berry, M.J.; Seale, L.A. From Selenium Absorption to Selenoprotein Degradation. Biol. Trace Elem. Res. 2019, 192, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jang, J.K.; Ko, K.Y.; Jin, Y.; Ham, M.; Kang, H.; Kim, I.Y. Degradation of selenoprotein S and selenoprotein K through PPARγ-mediated ubiquitination is required for adipocyte differentiation. Cell Death Differ. 2019, 26, 1007. [Google Scholar] [CrossRef]
- Olsson, M.; Olsson, B.; Jacobson, P.; Thelle, D.S.; Björkegren, J.; Walley, A.; Sjöholm, K. Expression of the selenoprotein S (SELS) gene in subcutaneous adipose tissue and SELS genotype are associated with metabolic risk factors. Metabolism 2011, 60, 114–120. [Google Scholar] [CrossRef]
- Xie, X.; Yi, Z.; Bowen, B.; Wolf, C.; Flynn, C.R.; Sinha, S.; Meyer, C. Characterization of the human adipocyte proteome and reproducibility of protein abundance by one-dimensional gel electrophoresis and HPLC−ESI−MS/MS. J. Proteome Res. 2010, 9, 4521–4534. [Google Scholar] [CrossRef] [Green Version]
- Hung, S.C.; Chang, C.F.; Ma, H.L.; Chen, T.H.; Ho, L.L.T. Gene expression profiles of early adipogenesis in human mesenchymal stem cells. Gene 2004, 340, 141–150. [Google Scholar] [CrossRef]
- Yamasaki, T.; Tahara, K.; Takano, S.; Inoue-Murayama, M.; Rose, M.T.; Minashima, T.; Ito, S.I. Mechanism of plasma glutathione peroxidase production in bovine adipocytes. Cell Tissue Res. 2006, 326, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Calzadilla, P.; Sapochnik, D.; Cosentino, S.; Diz, V.; Dicelio, L.; Calvo, J.C.; Guerra, L.N. N-acetylcysteine reduces markers of differentiation in 3T3-L1 adipocytes. Int. J. Mol. 2011, 12, 6936–6951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundholm, L.; Putnik, M.; Otsuki, M.; Andersson, S.; Ohlsson, C.; Gustafsson, J.A.; Dahlman-Wright, K. Effects of estrogen on gene expression profiles in mouse hypothalamus and white adipose tissue: Target genes include glutathione peroxidase 3 and cell death-inducing DNA fragmentation factor, alpha-subunit-like effector A. J. Endocrinol. 2008, 196, 547–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankovic, A.; Korac, A.; Srdic-Galic, B.; Buzadzic, B.; Otasevic, V.; Stancic, A.; Korac, B. Differences in the redox status of human visceral and subcutaneous adipose tissues–relationships to obesity and metabolic risk. Metabolism 2014, 63, 661–671. [Google Scholar] [CrossRef]
- Sriram, S.; Yuan, C.; Chakraborty, S.; Tay, W.; Park, M.; Shabbir, A.; Sugii, S. Oxidative stress mediates depot-specific functional differences of human adipose-derived stem cells. Stem Cell Res. Ther. 2019, 10, 141. [Google Scholar] [CrossRef]
- Langhardt, J.; Flehmig, G.; Klöting, N.; Lehmann, S.; Ebert, T.; Kern, M.; Fasshauer, M. Effects of Weight Loss on Glutathione Peroxidase 3 Serum Concentrations and Adipose Tissue Expression in Human Obesity. Obes. Facts 2018, 11, 475–490. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Khemka, V.K.; Chatterjee, G.; Ganguly, A.; Mukhopadhyay, S.; Chakrabarti, S. Enhanced ROS production and oxidative damage in subcutaneous white adipose tissue mitochondria in obese and type 2 diabetes subjects. Mol. Cell. Biochem. 2015, 399, 95–103. [Google Scholar] [CrossRef]
- Yang, S.; Zhu, H.; Li, Y.; Lin, H.; Gabrielson, K.; Trush, M.A.; Diehl, A.M. Mitochondrial adaptations to obesity-related oxidant stress. Arch. Biochem. Biophys. 2000, 378, 259–268. [Google Scholar] [CrossRef]
- Kobayashi, H.; Matsuda, M.; Fukuhara, A.; Komuro, R.; Shimomura, I. Dysregulated glutathione metabolism links to impaired insulin action in adipocytes. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1326–E1334. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest 2017, 114, 1752–1761. [Google Scholar] [CrossRef]
- Asayama, K.; Nakane, T.; Dobashi, K.; Kodera, K.; Hayashibe, H.; Uchida, N.; Nakazawa, S. Effect of obesity and troglitazone on expression of two glutathione peroxidases: Cellular and extracellular types in serum, kidney and adipose tissue. Free Radic. Res. 2001, 34, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Galinier, A.; Carrière, A.; Fernandez, Y.; Carpéné, C.; André, M.; Caspar-Bauguil, S.; Casteilla, L. Adipose tissue proadipogenic redox changes in obesity. J. Biol. Chem. 2006, 281, 12682–12687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobgen, W.; Fu, W.J.; Gao, H.; Li, P.; Meininger, C.J.; Smith, S.B.; Wu, G. High fat feeding and dietary L-arginine supplementation differentially regulate gene expression in rat white adipose tissue. Amino Acids 2009, 37, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Kim, J.B. Dysregulation of adipose glutathione peroxidase 3 in obesity contributes to local and systemic oxidative stress. Mol. Endocrinol. 2008, 22, 2176–2189. [Google Scholar] [CrossRef] [Green Version]
- Franck, N.; Gummesson, A.; Jernås, M.; Glad, C.; Svensson, P.A.; Guillot, G.; Olsson, B. Identification of adipocyte genes regulated by caloric intake. J. Clin. Endocrinol. Metab. 2011, 96, E413–E418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Cao, J.; Li, J.; He, W. Adipose tissue-specific PPARγ deficiency increases resistance to oxidative stress. Exp. Gerontol. 2008, 43, 154–163. [Google Scholar] [CrossRef]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.D.; Vatamaniuk, M.Z.; Wang, S.K.; Roneker, C.A.; Simmons, R.A.; Lei, X.G. Molecular mechanisms for hyperinsulinaemia induced by overproduction of selenium-dependent glutathione peroxidase-1 in mice. Diabetologia 2008, 51, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Merry, T.L.; Tran, M.; Stathopoulos, M.; Wiede, F.; Fam, B.C.; Dodd, G.T.; Tiganis, T. High-fat-fed obese glutathione peroxidase 1-deficient mice exhibit defective insulin secretion but protection from hepatic steatosis and liver damage. Antioxid. Redox Signal. 2014, 20, 2114–2129. [Google Scholar] [CrossRef]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Stepto, N. Reactive oxygen species enhance insulin sensitivity. Cell Metab. 2009, 10, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Choi, E.J.; Kie, J.H.; Lee, J.H.; Seoh, J.Y. Deficiency of glutathione peroxidase-1 and catalase attenuated diet-induced obesity and associated metabolic disorders. Acta Diabetol. 2019, 57, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Katunga, L.A.; Gudimella, P.; Efird, J.T.; Abernathy, S.; Mattox, T.A.; Beatty, C.; Virag, J.A. Obesity in a model of gpx4 haploinsufficiency uncovers a causal role for lipid-derived aldehydes in human metabolic disease and cardiomyopathy. Mol. Metab. 2015, 4, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Pepper, M.P.; Vatamaniuk, M.Z.; Yan, X.; Roneker, C.A.; Lei, X.G. Impacts of dietary selenium deficiency on metabolic phenotypes of diet-restricted gpx1-overexpressing mice. Antioxid. Redox. Signal. 2011, 14, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Pepper, M.P.; Vatamaniuk, M.Z.; Roneker, C.A.; Li, L.; Lei, X.G. Dietary selenium deficiency partially rescues type 2 diabetes-like phenotypes of glutathione peroxidase-1-overexpressing male mice. J. Nutr. 2012, 142, 1975–1982. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Yun, J.W.; Lei, X.G. Novel regulation and signaling of glucose-stimulated insulin secretion in islets by Se-glutathione peroxidase-1 mimic ebselen. Antioxid. Redox. Signal. 2014, 20, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Tinkov, A.A.; Bjørklund, G.; Skalny, A.V.; Holmgren, A.; Skalnaya, M.G.; Chirumbolo, S.; Aaseth, J. The role of the thioredoxin/thioredoxin reductase system in the metabolic syndrome: Towards a possible prognostic marker? Cell Mol. Life Sci. 2018, 75, 1567–1586. [Google Scholar] [CrossRef]
- Rajalin, A.M.; Micoogullari, M.; Sies, H.; Steinbrenner, H. Upregulation of the thioredoxin-dependent redox system during differentiation of 3T3-L1 cells to adipocytes. Biol. Chem. 2014, 395, 667–677. [Google Scholar] [CrossRef]
- Chang, C.H.; Liao, H.X.Q.; Hsu, F.L.; Ho, C.T.; Liao, V.H.C. N-γ-(l-Glutamyl)-l-Selenomethionine Inhibits Fat Storage via the Stearoyl-CoA Desaturases FAT-6 and FAT-7 and the Selenoprotein TRXR-1 in Caenorhabditis elegans. Mol. Nutr. Food Res. 2019, 63, 1800784. [Google Scholar] [CrossRef]
- Peng, X.; Giménez-Cassina, A.; Petrus, P.; Conrad, M.; Rydén, M.; Arnér, E.S. Thioredoxin reductase 1 suppresses adipocyte differentiation and insulin responsiveness. Sci. Rep. 2016, 6, 28080. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, B.R.; Hansen, J.M. Differential redox potential profiles during adipogenesis and osteogenesis. Cell Mol. Biol. Lett. 2011, 16, 149–161. [Google Scholar] [CrossRef]
- Ovadia, H.; Haim, Y.; Nov, O.; Almog, O.; Kovsan, J.; Bashan, N.; Rudich, A. Increased Adipocyte S-Nitrosylation Targets Anti-lipolytic Action of Insulin Relevance to Adipose Tissue Dysfunction in Obesity. J. Biol. Chem. 2011, 286, 30433–30443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.S.; Cho, H.H.; Lee, B.J.; Bae, Y.C.; Jung, J.S. Role of thioredoxin 1 and thioredoxin 2 on proliferation of human adipose tissue-derived mesenchymal stem cells. Stem Cells Dev. 2010, 20, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Chutkow, W.A.; Lee, R.T. Thioredoxin regulates adipogenesis through thioredoxin-interacting protein (Txnip) protein stability. J. Biol. Chem. 2011, 286, 29139–29145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.K.; Sharma, N.K.; Hasstedt, S.J.; Mondal, A.K.; Ma, L.; Langberg, K.A.; Elbein, S.C. An integrative genomics approach identifies activation of thioredoxin/thioredoxin reductase-1-mediated oxidative stress defense pathway and inhibition of angiogenesis in obese nondiabetic human subjects. J. Clin. Endocrinol. Metab. 2014, 96, E1308–E1313. [Google Scholar] [CrossRef]
- Peña-Orihuela, P.; Camargo, A.; Rangel-Zuñiga, O.A.; Perez-Martinez, P.; Cruz-Teno, C.; Delgado-Lista, J.; Roche, H.M. Antioxidant system response is modified by dietary fat in adipose tissue of metabolic syndrome patients. J. Nutr. Biochem. 2013, 24, 1717–1723. [Google Scholar] [CrossRef]
- Santini, F.; Marzullo, P.; Rotondi, M.; Ceccarini, G.; Pagano, L.; Ippolito, S.; Biondi, B. The crosstalk between thyroid gland and adipose tissue: Signal integration in health and disease. Eur. J. Endocrinol. 2014, 171, R137–R152. [Google Scholar] [CrossRef]
- Nomura, E.; Toyoda, N.; Harada, A.; Nishimura, K.; Ukita, C.; Morimoto, S.; Nishikawa, M. Type 2 iodothyronine deiodinase is expressed in human preadipocytes. Thyroid 2011, 21, 305–310. [Google Scholar] [CrossRef]
- Calvo, R.M.; Obregon, M.J. Presence and regulation of D1 and D2 deiodinases in rat white adipose tissue. Metabolism 2011, 60, 1207–1210. [Google Scholar] [CrossRef] [Green Version]
- Obregon, M.J. Adipose tissues and thyroid hormones. Front. Physiol. 2014, 5, 479. [Google Scholar] [CrossRef] [Green Version]
- Ortega, F.J.; Jilkova, Z.M.; Moreno-Navarrete, J.M.; Pavelka, S.; Rodriguez-Hermosa, J.I.; Kopeckỳ, J.; Fernandez-Real, J.M. Type I iodothyronine 5′-deiodinase mRNA and activity is increased in adipose tissue of obese subjects. Int. J. Obes. 2012, 36, 320–324. [Google Scholar] [CrossRef] [Green Version]
- Jilkova, Z.M.; Pavelka, S.; Flachs, P.; Hensler, M.; Kus, V.; Kopecký, J. Modulation of type I iodothyronine 5’-deiodinase activity in white adipose tissue by nutrition: Possible involvement of leptin. Physiol. Res. 2010, 59, 561–569. [Google Scholar]
- Bradley, D.; Liu, J.; Blaszczak, A.; Wright, V.; Jalilvand, A.; Needleman, B.; Hsueh, W. Adipocyte DIO2 Expression Increases in Human Obesity but Is Not Related to Systemic Insulin Sensitivity. J. Diabetes Res. 2018, 2018, 2464652. [Google Scholar] [CrossRef] [PubMed]
- Kurylowicz, A.; Jonas, M.; Lisik, W.; Jonas, M.; Wicik, Z.A.; Wierzbicki, Z.; Puzianowska-Kuznicka, M. Obesity is associated with a decrease in expression but not with the hypermethylation of thermogenesis-related genes in adipose tissues. J. Transl. Med. 2015, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Akarsu, E.; Korkmaz, H.; Balci, S.O.; Borazan, E.; Korkmaz, S.; Tarakcioglu, M. Subcutaneous adipose tissue type II deiodinase gene expression reduced in obese individuals with metabolic syndrome. Exp. Clin. Endocrinol. 2016, 124, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Marsili, A.; Aguayo-Mazzucato, C.; Chen, T.; Kumar, A.; Chung, M.; Lunsford, E.P.; Chou, C. Mice with a targeted deletion of the type 2 deiodinase are insulin resistant and susceptible to diet induced obesity. PLoS ONE 2011, 6, e20832. [Google Scholar] [CrossRef]
- Castillo, M.; Hall, J.A.; Correa-Medina, M.; Ueta, C.; Kang, H.W.; Cohen, D.E.; Bianco, A.C. Disruption of thyroid hormone activation in type 2 deiodinase knockout mice causes obesity with glucose intolerance and liver steatosis only at thermoneutrality. Diabetes 2011, 60, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, T.L.; Fernandes, G.W.; McAninch, E.A.; Bocco, B.M.; Abdalla, S.M.; Ribeiro, M.O.; Wang, T. Perinatal deiodinase 2 expression in hepatocytes defines epigenetic susceptibility to liver steatosis and obesity. Proc. Natl. Acad. Sci. USA 2015, 112, 14018–14023. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, F.; Gentile, A.; Silvestri, E.; Goglia, F.; Lombardi, A. Effect of iodothyronines on thermogenesis: Focus on brown adipose tissue. Front. Endocrinol. 2018, 9, 254. [Google Scholar] [CrossRef] [Green Version]
- Cabanelas, A.; Lisboa, P.C.; Moura, E.G.; Pazos-Moura, C.C. Leptin acute modulation of the 5′-deiodinase activities in hypothalamus, pituitary and brown adipose tissue of fed rats. Horm. Metab. Res. 2006, 38, 481–485. [Google Scholar] [CrossRef]
- Boelen, A.; Kwakkel, J.; Vos, X.G.; Wiersinga, W.M.; Fliers, E. Differential effects of leptin and refeeding on the fasting-induced decrease of pituitary type 2 deiodinase and thyroid hormone receptor β2 mRNA expression in mice. J. Endocrinol. 2006, 190, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Coppola, A.; Meli, R.; Diano, S. Inverse shift in circulating corticosterone and leptin levels elevates hypothalamic deiodinase type 2 in fasted rats. Endocrinology 2005, 146, 2827–2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, I.; Garcia-Fuentes, E. Iodine and adipocytokines: Cellular aspects. In Molecular, Genetic, and Nutritional Aspects of Major and Trace Minerals; Collins, J.F., Ed.; Academic Press: Pittsburgh, PA, USA, 2017; pp. 151–157. [Google Scholar]
- Yokoyama, J.; So, K.H.; Suzuki, Y.; Morioka, M.; Suzuki, K.; Katoh, K.; Roh, S. Anti-obese effect of iodine-enriched yolk in cultured adipocytes. Nihon Chikusan Gakkaiho 2016, 87, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Solovyev, N.; Vanhaecke, F.; Michalke, B. Selenium and iodine in diabetes mellitus with a focus on the interplay and speciation of the elements. J. Trace Elem. Med. Biol. 2019, 56, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wu, Y.; Zeng, R.; Liao, K. Proteomic profiling of lipid droplet-associated proteins in primary adipocytes of normal and obese mouse. Acta Biochim. Biophys. Sin. 2012, 44, 394–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.S.; Jung, J.S.; Jang, J.H.; Kang, K.S.; Kang, S.K. Nuclear Argonaute 2 regulates adipose tissue-derived stem cell survival through direct control of miR10b and selenoprotein N1 expression. Aging Cell 2011, 10, 277–291. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.; Wu, Y.; Ren, F.; Lei, X.G. Loss of Selenoprotein V Predisposes Mice to Body Fat Accumulation (OR11-02-19). Curr. Dev. Nutr. 2019, 3 (Suppl. 1), nzz044.OR11-02-19. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.L.; Huang, J.Q.; Xiao, Y.; Wu, Y.Y.; Ren, F.Z.; Lei, X.G. Knockout of Selenoprotein V Affects Regulation of Selenoprotein Expression by Dietary Selenium and Fat Intakes in Mice. J. Nutr. 2019, 150, 483–491. [Google Scholar] [CrossRef]
- Pitts, M.W.; Reeves, M.A.; Hashimoto, A.C.; Ogawa, A.; Kremer, P.; Seale, L.A.; Berry, M.J. Deletion of selenoprotein M leads to obesity without cognitive deficits. J. Biol. Chem. 2013, 288, 26121–26134. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.Y.; Wen, G.B.; Feng, S.D.; Tuo, Q.H.; Ou, H.S.; Yao, C.H.; Liao, D.F. MicroRNA-375 promotes 3T3-L1 adipocyte differentiation through modulation of extracellular signal-regulated kinase signalling. Clin. Exp. Pharmacol. Physiol. 2011, 38, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Uthus, E.O.; Picklo Sr, M.J. Obesity reduces methionine sulphoxide reductase activity in visceral adipose tissue. Free Radic. Res. 2011, 45, 1052–1060. [Google Scholar] [CrossRef]
- Novoselov, S.V.; Kim, H.Y.; Hua, D.; Lee, B.C.; Astle, C.M.; Harrison, D.E.; Gladyshev, V.N. Regulation of selenoproteins and methionine sulfoxide reductases A and B1 by age, calorie restriction, and dietary selenium in mice. Antioxid. Redox Signal. 2010, 12, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.G.; Cha, H.N.; Park, S.; Kim, Y.W.; Kim, J.Y.; Park, S.Y. Selenoprotein W Deficiency Does Not Affect Oxidative Stress and Insulin Sensitivity in the Skeletal Muscle of High-fat Diet-fed Obese Mice. Am. J. Physiol. Cell Physiol. 2019, 317, 1172–1182. [Google Scholar] [CrossRef] [PubMed]
- Govindarajah, V.; Leung, Y.K.; Ying, J.; Gear, R.; Bornschein, R.L.; Medvedovic, M.; Ho, S.M. In utero exposure of rats to high-fat diets perturbs gene expression profiles and cancer susceptibility of prepubertal mammary glands. J. Nutr. Biochem. 2016, 29, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Yang, T.; Pan, T.; Liu, C.; Li, S. Effect of low-selenium/high-fat diet on pig peripheral blood lymphocytes: Perspectives from selenoproteins, heat shock proteins, and cytokines. Biol. Trace Elem. Res. 2018, 183, 102–113. [Google Scholar] [CrossRef]
- Takamura, T.; Misu, H.; Matsuzawa-Nagata, N.; Sakurai, M.; Ota, T.; Shimizu, A.; Kaneko, S. Obesity upregulates genes involved in oxidative phosphorylation in livers of diabetic patients. Obesity 2008, 16, 2601–2609. [Google Scholar] [CrossRef]
- Slattery, M.L.; Lundgreen, A.; Welbourn, B.; Corcoran, C.; Wolff, R.K. Genetic variation in selenoprotein genes, lifestyle, and risk of colon and rectal cancer. PLoS ONE 2012, 7, e37312. [Google Scholar] [CrossRef]
- Donovan, J.; Copeland, P.R. The efficiency of selenocysteine incorporation is regulated by translation initiation factors. J. Mol. Biol. 2010, 400, 659–664. [Google Scholar] [CrossRef] [Green Version]
- Schoenmakers, E.; Schoenmakers, N.; Chatterjee, K. Mutations in humans that adversely affect the Selenoprotein synthesis pathway. In Selenium; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: Cham, Switzerland, 2016; pp. 523–538. [Google Scholar]
- Çatli, G.; Fujisawa, H.; Kirbiyik, Ö.; Mimoto, M.S.; Gençpinar, P.; Özdemir, T.R.; Dumitrescu, A.M. A novel homozygous selenocysteine insertion sequence binding protein 2 (SECISBP2, SBP2) gene mutation in a turkish boy. Thyroid 2018, 28, 1221–1223. [Google Scholar] [CrossRef]
- Wang, N.; Tan, H.Y.; Li, S.; Wang, D.; Xu, Y.; Zhang, C.; Feng, Y. SBP2 deficiency in adipose tissue macrophages drives insulin resistance in obesity. Sci. Adv. 2019, 5, eaav0198. [Google Scholar] [CrossRef] [Green Version]
- Seale, L.A. Selenocysteine β-Lyase: Biochemistry, regulation and physiological role of the selenocysteine decomposition enzyme. Antioxidants 2019, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Seale, L.A.; Gilman, C.L.; Hashimoto, A.C.; Ogawa-Wong, A.N.; Berry, M.J. Diet-induced obesity in the selenocysteine lyase knockout mouse. Antioxid. Redox Signal. 2015, 23, 761–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seale, L.; Khadka, V.; Menor, M.; Sasuclark, A.; Guirguis, K.; Hashimoto, A.; Deng, Y. RNA-Seq Analysis in Liver of Selenium-deficient Selenocysteine Lyase Knockout Mouse. Curr. Dev. Nutr. 2019, 3 (Suppl. 1), nzz044-P24. [Google Scholar] [CrossRef]
- Seale, L.A.; Hashimoto, A.C.; Kurokawa, S.; Gilman, C.L.; Seyedali, A.; Bellinger, F.P.; Berry, M.J. Disruption of the selenocysteine lyase-mediated selenium recycling pathway leads to metabolic syndrome in mice. Mol. Cell. Biol. 2012, 32, 4141–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, B.W.; Nam, E.; Org, E.; Kostem, E.; Norheim, F.; Hui, S.T.; Mehrabian, M. Genetic control of obesity and gut microbiota composition in response to high-fat, high-sucrose diet in mice. Cell Metab. 2013, 17, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, R.K.C.; Kingwell, B.A.; Carey, A.L. Human brown adipose tissue as a target for obesity management; beyond cold-induced thermogenesis. Obes. Rev. 2017, 18, 1227–1242. [Google Scholar] [CrossRef]
- Cettour-Rose, P.; Burger, A.G.; Meier, C.A.; Visser, T.J.; Rohner-Jeanrenaud, F. Central stimulatory effect of leptin on T3production is mediated by brown adipose tissue type II deiodinase. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E980–E987. [Google Scholar] [CrossRef] [Green Version]
- Christian, M. Transcriptional fingerprinting of “browning” white fat identifies NRG4 as a novel adipokine. Adipocyte 2015, 4, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.H.; Nicol, F.; Beckett, G.J. Selenium and iodine deficiencies: Effects on brain and brown adipose tissue selenoenzyme activity and expression. J. Endocrinol. 1997, 155, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Korać, B.; Buzadžić, B. Selenium supplementation and GSH-Px activity in the IBAT and erythrocytes of cold-adapted rats. Food Res. Int. 2002, 35, 221–224. [Google Scholar] [CrossRef]
- Sohn, J.W. Network of hypothalamic neurons that control appetite. BMB Rep. 2015, 48, 229. [Google Scholar] [CrossRef]
- Gong, T.; Torres, D.J.; Berry, M.J.; Pitts, M.W. Hypothalamic redox balance and leptin signaling-emerging role of selenoproteins. Free Radic. Biol. Med. 2018, 127, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Henry, F.E.; Sugino, K.; Tozer, A.; Branco, T.; Sternson, S.M. Cell type-specific transcriptomics of hypothalamic energy-sensing neuron responses to weight-loss. Elife 2015, 4, e09800. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Hashimoto, A.C.; Sasuclark, A.R.; Khadka, V.S.; Gurary, A.; Pitts, M.W. Selenoprotein M promotes hypothalamic leptin signaling and thioredoxin antioxidant activity. Antioxid. Redox Signal 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Yagishita, Y.; Uruno, A.; Fukutomi, T.; Saito, R.; Saigusa, D.; Pi, J.; Yamamoto, M. Nrf2 improves leptin and insulin resistance provoked by hypothalamic oxidative stress. Cell Rep. 2017, 18, 2030–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schriever, S.C.; Zimprich, A.; Pfuhlmann, K.; Baumann, P.; Giesert, F.; Klaus, V.; Wurst, W. Alterations in neuronal control of body weight and anxiety behavior by glutathione peroxidase 4 deficiency. Neuroscience 2017, 357, 241–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolatto, C.F.; Heck, S.O.; Zborowski, V.A.; Gai, B.M.; Neto, J.S.; Nogueira, C.W. Evidence for the contribution of multiple mechanisms in the feeding pattern of rats exposed to p-chloro-diphenyl diselenide-supplemented diets. Physiol. Behav. 2015, 151, 298–307. [Google Scholar] [CrossRef]
- Bortolatto, C.F.; Nogueira, C.W.; Porteiro, B.; Imbernón, M.; Nogueiras, R. Hypothalamic pathways regulate the anorectic action of p-chloro-diphenyl diselenide in rats. Eur. J. Pharmacol. 2017, 815, 241–250. [Google Scholar] [CrossRef]
- Li, J.; Cao, C.; Wang, L.; Jiang, X.; Zhang, C.; Xu, J. Changes in Growth and Feed Efficiency in Chickens Supplemented with Supernutritional and Excess Selenium are Associated with Changes in Appetite. FASEB J. 2015, 29, 136.5. [Google Scholar]
- Xia, S.F.; Duan, X.M.; Hao, L.Y.; Li, L.T.; Cheng, X.R.; Xie, Z.X.; Le, G.W. Role of thyroid hormone homeostasis in obesity-prone and obesity-resistant mice fed a high-fat diet. Metabolism 2015, 64, 566–579. [Google Scholar] [CrossRef]
- Wu, Z.; Martinez, M.E.; St. Germain, D.L.; Hernandez, A. Type 3 deiodinase role on central thyroid hormone action affects the leptin-melanocortin system and circadian activity. Endocrinology 2017, 158, 419–430. [Google Scholar]
- Yoon, B.I.; Kim, S.J.; Cho, H.J.; Ha, U.; Hong, S.H.; Lee, J.Y.; Kim, S.W. The Effect of Selenium-rich Hot Spring Water on Serum Leptin and IGF-1 in a Rat Model. Korean J. Androl. 2010, 28, 132–138. [Google Scholar]
- Ojeda, M.L.; Carreras, O.; Díaz-Castro, J.; Murillo, M.L.; Nogales, F. High-and low-selenium diets affect endocrine energy balance during early programming. Toxicol. Appl. Pharmacol. 2019, 382, 114744. [Google Scholar] [CrossRef] [PubMed]
- Chanoine, J.P.; Wong, A.C.; Lavoie, J.C. Selenium deficiency impairs corticosterone and leptin responses to adrenocorticotropin in the rat. Biofactors 2004, 20, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Liu, Q.; Xu, H.B.; Zhu, Y.S.; Yang, X.L. Effects of selenium overexposure on glutathione peroxidase and thioredoxin reductase gene expressions and activities. Biol. Trace Elem. Res. 2002, 89, 165–175. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Lee, B.C.; Handy, D.E.; Loscalzo, J.; Hatfield, D.L.; Gladyshev, V.N. Both maximal expression of selenoproteins and selenoprotein deficiency can promote development of type 2 diabetes-like phenotype in mice. Antioxid. Redox Signal. 2011, 14, 2327–2336. [Google Scholar] [CrossRef]
- Michalke, B. Metallomics: Analytical Techniques and Speciation Methods; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2016. [Google Scholar]
- Bierla, K.; Szpunar, J.; Lobinski, R. Biological selenium species and selenium speciation in biological samples. In Selenium; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: Cham, Switzerland, 2016; pp. 413–424. [Google Scholar]
- Panchal, S.K.; Wanyonyi, S.; Brown, L. Selenium, vanadium, and chromium as micronutrients to improve metabolic syndrome. Curr. Hypertens. Rep. 2017, 19, 10. [Google Scholar] [CrossRef]
- Vinceti, M.; Maraldi, T.; Bergomi, M.; Malagoli, C. Risk of chronic low-dose selenium overexposure in humans: Insights from epidemiology and biochemistry. Rev. Environ. Health 2009, 24, 231–248. [Google Scholar] [CrossRef]
- Wronska, A.; Kmiec, Z. Structural and biochemical characteristics of various white adipose tissue depots. Acta Physiol. 2012, 205, 194–208. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tinkov, A.A.; Ajsuvakova, O.P.; Filippini, T.; Zhou, J.-C.; Lei, X.G.; Gatiatulina, E.R.; Michalke, B.; Skalnaya, M.G.; Vinceti, M.; Aschner, M.; et al. Selenium and Selenoproteins in Adipose Tissue Physiology and Obesity. Biomolecules 2020, 10, 658. https://doi.org/10.3390/biom10040658
Tinkov AA, Ajsuvakova OP, Filippini T, Zhou J-C, Lei XG, Gatiatulina ER, Michalke B, Skalnaya MG, Vinceti M, Aschner M, et al. Selenium and Selenoproteins in Adipose Tissue Physiology and Obesity. Biomolecules. 2020; 10(4):658. https://doi.org/10.3390/biom10040658
Chicago/Turabian StyleTinkov, Alexey A., Olga P. Ajsuvakova, Tommaso Filippini, Ji-Chang Zhou, Xin Gen Lei, Eugenia R. Gatiatulina, Bernhard Michalke, Margarita G. Skalnaya, Marco Vinceti, Michael Aschner, and et al. 2020. "Selenium and Selenoproteins in Adipose Tissue Physiology and Obesity" Biomolecules 10, no. 4: 658. https://doi.org/10.3390/biom10040658
APA StyleTinkov, A. A., Ajsuvakova, O. P., Filippini, T., Zhou, J. -C., Lei, X. G., Gatiatulina, E. R., Michalke, B., Skalnaya, M. G., Vinceti, M., Aschner, M., & Skalny, A. V. (2020). Selenium and Selenoproteins in Adipose Tissue Physiology and Obesity. Biomolecules, 10(4), 658. https://doi.org/10.3390/biom10040658