Vitamin K in Vertebrates’ Reproduction: Further Puzzling Pieces of Evidence from Teleost Fish Species



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
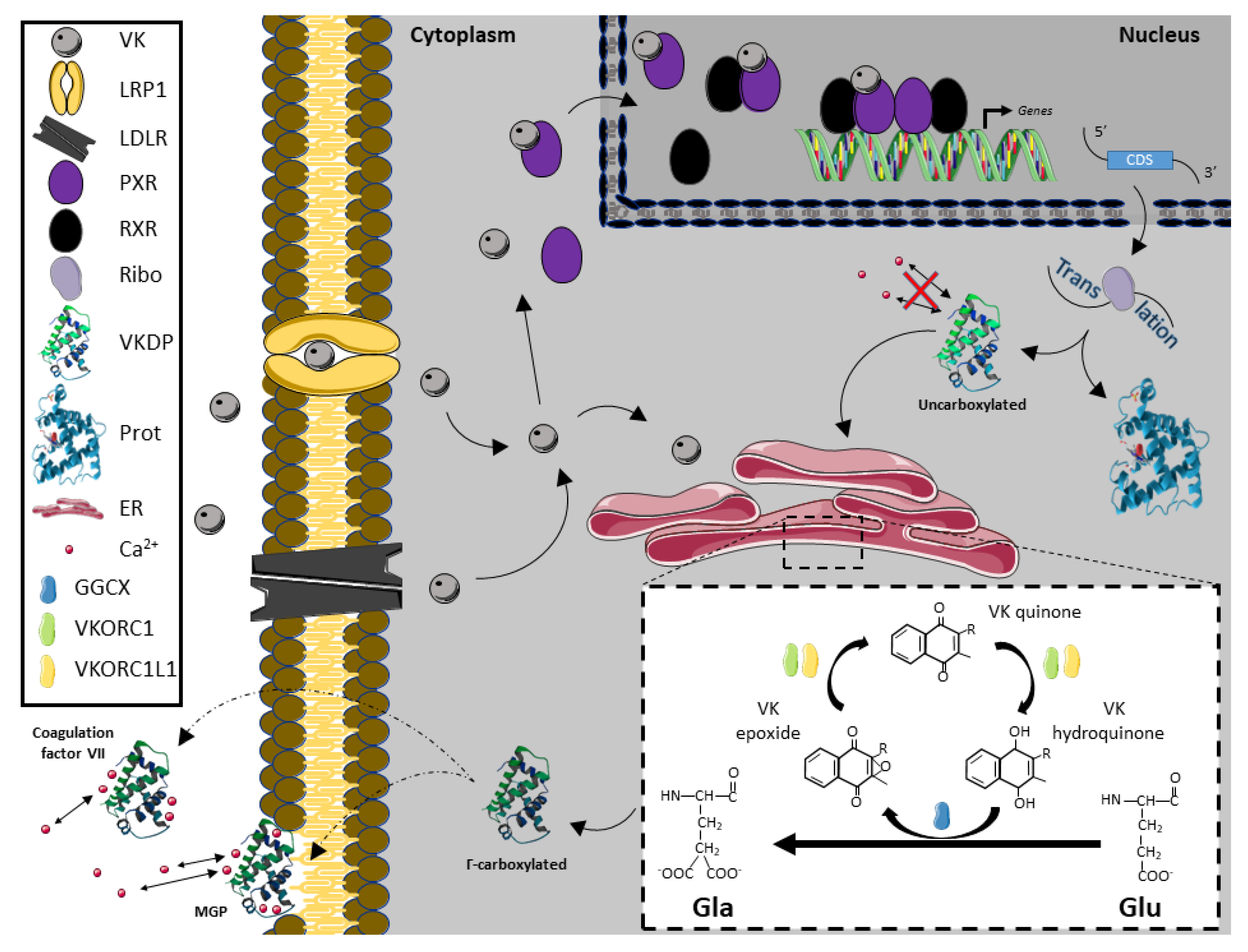
:1. Vitamin K Metabolites, Sources and Metabolism
2. Vitamin K Molecular Pathways
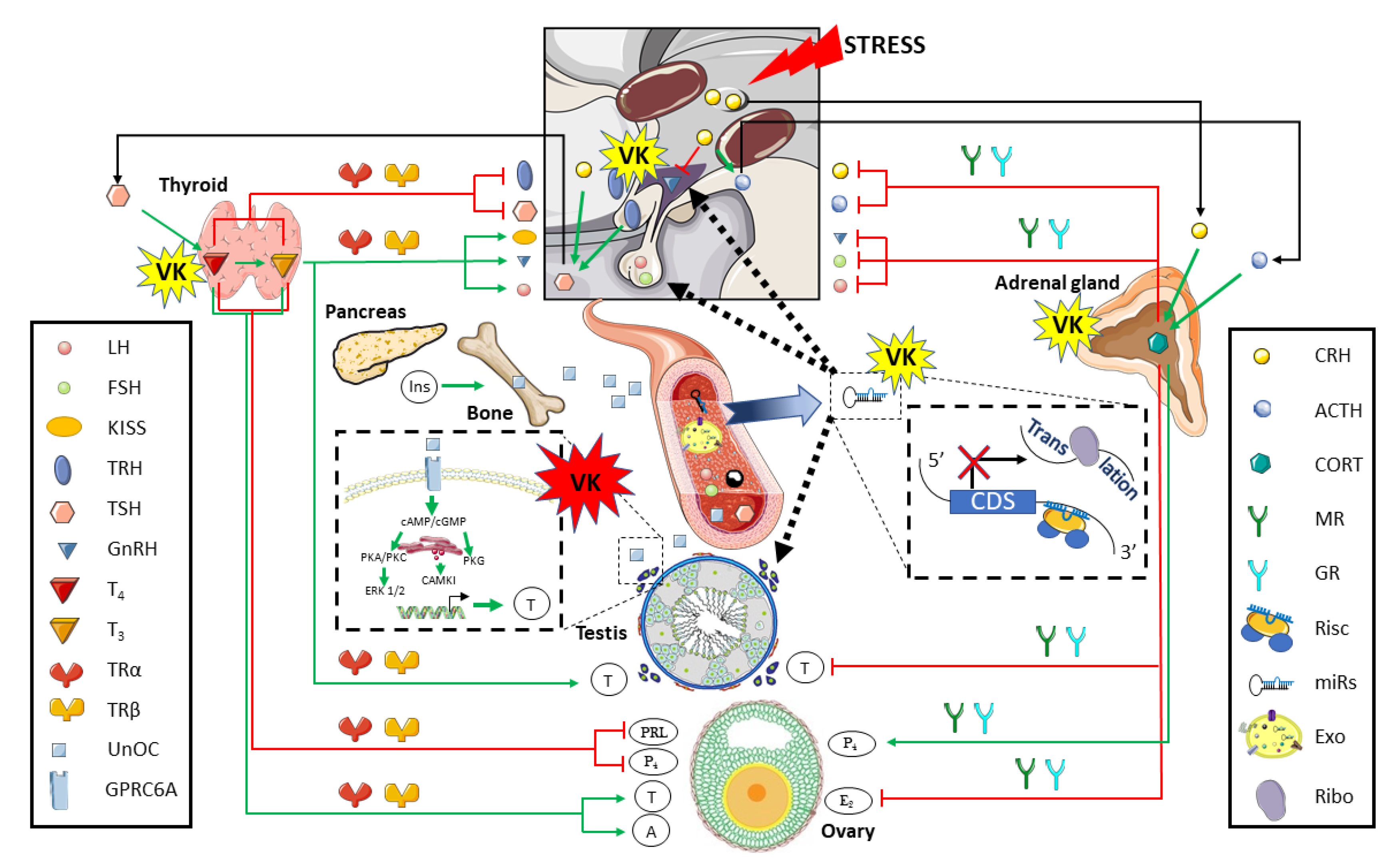
3. The Impact of Vitamin K in Reproduction
3.1. Direct Impact of Vitamin K on the Gonads
3.2. The Vitamin K Impact on the Hypothalamus-Pituitary-Gonad Axis
3.3. The Vitamin K Impact on Tissue Crosstalk Relevant for Reproductive Performance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fernández, I.; Gavaia, P.; Darias, M.J.; Gisbert, E. Fat-soluble vitamins in fish: A transcriptional tissue-specific crosstalk that remains to be unveiled and characterized. In Emerging Issues in Fish Larvae Research; Yúfera, M., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 159–208. [Google Scholar]
- Harshman, S.G.; Saltzman, E.; Booth, S.L. Vitamin K: Dietary intake and requirements in different clinical conditions. Curr. Opin. Clin. Nutr. Metab. Care. 2014, 17, 438–531. [Google Scholar] [CrossRef] [PubMed]
- Lambert, W.E.; De Leenher, A.P. Vitamin K. In Modern Chromatographic Analysis of Vitamins, 2nd ed.; De Leenher, A.P., Lamberts, W.E., Nelis, H.J., Eds.; Marcel Dekker: New York, NY, USA, 1992; Volume 84, pp. 214–251. [Google Scholar]
- Booth, S.L.; Suttie, J.W. Dietary intake and adequacy of vitamin K. J. Nutr. 1998, 128, 785–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fodor, D.; Albu, A.; Poanta, L.; Porojan, M. Vitamin K and vascular calcifications. Acta Physiol. Hung. 2010, 97, 256–266. [Google Scholar] [CrossRef]
- Sobinoff, A.P.; Pye, V.; Nixon, B.; Roman, S.D.; McLaughlin, E.A. Adding insult to injury: Effects of xenobiotic-induced preantral ovotoxicity on ovarian development and oocyte fusibility. Toxicol. Sci. 2010, 118, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Food, E.; Authority, S. Scientific opinion on the safety and efficacy of vitamin K3 (menadione sodium bisulphite and menadione nicotinamide bisulphite) as a feed additive for all animal species. EFSA J. 2014, 12. [Google Scholar]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef]
- Shearer, M.J.; Okano, T. Key pathways and regulators of vitamin K function and intermediary metabolism. Annu. Rev. Nutr. 2018, 38, 127–151. [Google Scholar] [CrossRef]
- Kohlmeier, M.; Salomon, A.; Saupe, J.; Shearer, M.J. Transport of vitamin K to bone in Humans. J. Nutr. 1996, 126, 1192S–1196S. [Google Scholar] [CrossRef] [Green Version]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (Vitamin K3) is a catabolic product of oral phylloquinone (vitamin K1) in the intestine and a circulating precursor of tissue menaquinone-4 (vitamin K2) in rats. Int. J. Biol. Chem. 2013, 288, 33071–33080. [Google Scholar] [CrossRef] [Green Version]
- Thijssen, H.H.; Vervoort, L.M.; Schurgers, L.J.; Shearer, M.J. Menadione is a metabolite of oral vitamin K. Br. J. Nutr. 2006, 95, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Funahashi, N.; Hirota, Y.; Nakagawa, K.; Sawada, N.; Watanabe, M.; Suhara, Y.; Okano, T. YY1 Positively regulates human UBIAD1 expression. Biochem. Biophys. Res. Commun. 2015, 460, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Kim, S.S.; Garland, K.; Fuentes, I.; DiCarlo, L.M.; Ellis, J.L.; Fu, X.; Booth, S.L.; Evers, B.M.; DeBose-Boyd, R.A. Enhanced ER-associated degradation of HMG CoA reductase causes embryonic lethality associated with Ubiad1 deficiency. eLife 2020, 9, e54841. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Vitamin and mineral requirements in human nutrition Second Edition; World Health Organization: Geneva, Switzerland, 1998; pp. 1–20. ISBN 9241546123. [Google Scholar]
- Ferland, G.; Doucet, I.; Mainville, D. Phylloquinone and menaquinone-4 tissue distribution at different life stages in male and female Sprague–Dawley rats fed different VK levels sinceweaning or subjected to a 40% calorie restriction since adulthood. Nutrients 2016, 8, 141. [Google Scholar] [CrossRef]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grun, F.; Errandi, J.L.; Romero, K.M.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 regulation of bone homeostasis is mediated by the steroid and xenobiotic receptor, SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef] [Green Version]
- Lamba, V.; Yasuda, K.; Lamba, J.K.; Assem, M.; Davila, J.; Strom, S.; Schuetz, E.G. PXR (NR1I2): Splice variants in human tissues, including brain, and identification of neurosteroids and nicotine as PXR activators. Toxicol. Appl. Pharmacol. 2004, 199, 251–265. [Google Scholar] [CrossRef]
- Azuma, K.; Casey, S.C.; Ito, M.; Urano, T.; Horie, K.; Ouchi, Y.; Kirchner, S.; Blumberg, B.; Inoue, S. Pregnane X receptor knockout mice display osteopenia with reduced bone formation and enhanced bone resorption. J. Endocrinol. 2010, 207, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Roberto, V.P.; Granadeiro, L.; Trindade, M.; Gavaia, P.J.; Laizé, V.; Cancela, M.L.; Fernández, I. The xenobiotic sensor PXR in a marine flatfish species (Solea senegalensis): Gene expression patterns and its regulation under different physiological conditions. Mar. Environ. Res. 2017, 130, 187–199. [Google Scholar] [CrossRef]
- Shearer, M.J. Vitamin K | Physiology. In Encyclopedia of Food Sciences and Nutrition (Second Edition); Caballero, B., Ed.; Academic Press: Cambridge, MA, USA, 2003; pp. 6039–6045. ISBN 9780122270550. [Google Scholar] [CrossRef]
- Krossoy, C.; Waagbo, R.; Fjelldal, P.G.; Wargelius, A.; Lock, E.J.; Graff, I.E.; Ornsrund, R. Dietary menadione nicotinamide bisulphite (vitamin K3) does not affect growth or bone health in first feeding fry of Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2009, 15, 638–649. [Google Scholar] [CrossRef]
- Thijssen, H.H.; Drittij-Reijnders, M.J.; Fischer, M.A. Phylloquinone and menaquinone-4 distribution in rats: Synthesis rather than uptake determines menaquinone-4 organ concentrations. J. Nutr. 1996, 126, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Halder, M.; Petsophonsakul, P.; Akbulut, A.C.; Pavlic, A.; Bohan, F.; Anderson, E.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K: Double bonds beyond coagulation insights into differences between vitamin K1 and K2 in health and disease. Int. J. Mol. Sci. 2019, 20, 896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beulens, J.W.J.; Booth, S.L.; Van Den Heuvel, E.G.H.M.; Stoecklin, E.; Baka, A.; Vermeer, C. The role of menaquinones (vitamin K2) in human health. Br. J. Nutr. 2013, 110, 1357–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shea, M.K.; Booth, S.L.; Nettleton, J.A.; Burke, G.L.; Chen, H.; Kritchevsky, S.B. Circulating phylloquinone concentrations of adults in the united states differ according to race and ethnicity. J. Nutr. 2012, 142, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Westerman, K.; Kelly, J.M.; Ordovás, J.M.; Booth, S.L.; De Meo, D.L. Epigenome-Wide association study reveals a molecular signature of response to phylloquinone (vitamin K1) supplementation. Epigenetics 2020, 15, 859–870. [Google Scholar] [CrossRef]
- Harshman, S.G.; Shea, M.K. The role of Vitamin K in chronic aging diseases: Inflammation, cardiovascular disease, and osteoarthritis. Curr. Nutr. Rep. 2016, 5, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Lacombe, J.; Ferron, M. VKORC1L1, an enzyme mediating the effect of vitamin K in liver and extrahepatic tissues. Nutrients 2018, 10, 970. [Google Scholar] [CrossRef] [Green Version]
- Lacombe, J.; Rishavy, M.A.; Berkner, K.L.; Ferron, M. VKOR paralog VKORC1L1 supports vitamin K-dependent protein carboxylation in vivo. JCI Insight 2018, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Beato, S.; Marques, C.; Laizé, V.; Gavaia, P.J.; Fernández, I. New insights on vitamin K metabolism in Senegalese sole (Solea Senegalensis) based on ontogenetic and tissue-specific vitamin K epoxide reductase molecular data. Int. J. Mol. Sci. 2020, 21, 3489. [Google Scholar] [CrossRef]
- Finnan, E.G.; Harshman, S.G.; Haytowitz, D.B.; Booth, S.L. Mixed dishes are an unexpected source of dietary Vitamin K. J. Food Compos. Anal. 2017, 64, 127–131. [Google Scholar] [CrossRef]
- Turck, D.; Bresson, J.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Dietary reference values for vitamin K. EFSA J. 2017, 15, 1–78. [Google Scholar] [CrossRef]
- Shea, M.K.; Booth, S.L. Concepts and controversies in evaluating vitamin K status in population-based studies. Nutrients 2016, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.; Sun, H.; Raymond, R.M.; Furie, B.C.; Furie, B.; Bronstein, M.; Kaufman, R.J.; Westrick, R.; Ginsburg, D. Fatal hemorrhage in mice lacking γ-Glutamyl carboxylase. Blood 2007, 109, 5270–5275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkner, K.L. Vitamin K-Dependent Carboxylation. Vitam. Horm. 2008, 78, 131–156. [Google Scholar] [CrossRef]
- Fernández, I.; Santos, A.; Cancela, M.L.; Laizé, V.; Gavaia, P.J. Warfarin, a potential pollutant in aquatic environment acting through Pxr signaling pathway and γ-glutamyl carboxylation of vitamin K-dependent proteins. Environ. Pollut. 2014, 194, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Atkins, J.G.; Welldon, J.K.; Wijenayaka, R.A.; Bonewald, F.L.; Findlay, M.D. Vitamin K promotes mineralization, osteoblast-to-osteocyte transition, and an anticatabolic phenotype by γ-carboxylation-dependent and -independent mechanisms. Am. J. Physiol. Cell Physiol. 2009, 297, C1358–C1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, N.; Fernández, I.; Wulff, T.; Hamre, K.; Cancela, L.; Conceição, L.E.; Gavaia, P.J. Dietary supplementation with vitamin K affects transcriptome and proteome of Senegalese sole, improving larval performance and quality. Mar. Biotechnol. 2014, 16, 522–537. [Google Scholar] [CrossRef]
- Cardeira, J.; Gavaia, P.J.; Fernández, I.; Cengiz, I.F.; Moreira-Silva, J.; Oliveira, J.M.; Reis, R.L.; Cancela, M.L.; Laizé, V. Quantitative assessment of the regenerative and mineralogenic performances of the zebrafish caudal fin. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Granadeiro, L.; Dirks, R.P.; Ortiz-Delgado, J.B.; Gavaia, P.J.; Sarasquete, C.; Laizé, V.; Cancela, M.L.; Fernández, I. Warfarin-exposed zebrafish embryos resembles human warfarin embryopathy in a dose and developmental-time dependent manner – From molecular mechanisms to environmental concerns. Ecotox. Environ. Saf. 2019, 181, 559–571. [Google Scholar] [CrossRef]
- Shea, M.K.; Booth, S.L.; Harshman, S.G.; Smith, D.; Carlson, C.S.; Harper, L.; Armstrong, A.R.; Fang, M.; Cancela, M.L.; Simão, M.; et al. The effect of vitamin K insufficiency on histological and structural properties of knee joints in aging mice. Osteoarthr. Cartil. Open 2020, 100078. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Uitto, J.; Reutelingsperger, C.P. Vitamin K-dependent carboxylation of matrix gla-protein: A crucial switch to control ectopic mineralization. Trends Mol. Med. 2013, 19, 217–226. [Google Scholar] [CrossRef]
- Nowak, J.K.; Grzybowska-Chlebowczyk, U.; Landowski, P.; Szaflarska-Poplawska, A.; Klincewicz, B.; Adamczak, D.; Banasiewicz, T.; Plawski, A.; Walkowiak, J. Prevalence and correlates of vitamin K deficiency in children with inflammatory bowel disease. Sci. Rep. 2014, 4, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Scheiber, D.; Veulemans, V.; Horn, P.; Chatrou, M.L.; Potthoff, S.A.; Kelm, M.; Schurgers, L.J.; Westenfeld, R. High-dose menaquinone-7 supplementation reduces cardiovascular calcification in a murine model of extraosseous calcification. Nutrients 2015, 7, 6991–7011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westhofen, P.; Watzka, M.; Marinova, M.; Hass, M.; Kirfel, G.; Muller, J.; Bevans, C.G.; Muller, C.R.; Oldenburg, J. Human vitamin K 2,3-Epoxide reductase complex subunit 1-like 1 (VKORC1L1) mediates vitamin K-Dependent intracellular antioxidant function. J. Biol. Chem. 2011, 286, 15085–15094. [Google Scholar] [CrossRef] [Green Version]
- Sidorova, Y.A.; Perepechaeva, M.L.; Pivovarova, E.N.; Markel, A.L.; Lyakhovich, V.V.; Grishanova, A.Y. Menadione suppresses benzo(α)pyrene-induced activation of cytochromes P450 1A: Insights into a possible molecular mechanism. PLoS ONE 2016, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ferland, G. Vitamin K, an emerging nutrient in brain function. Biofactors 2012, 38, 151–157. [Google Scholar] [CrossRef]
- Fernández, I.; Fernandes, J.M.O.; Roberto, V.P.; Kopp, M.; Oliveira, C.; Riesco, M.F.; Dias, J.; Cox, C.J.; Leonor-Cancela, M.; Cabrita, E.; et al. Circulating small non-coding RNAs provide new insights into vitamin K nutrition and reproductive physiology in teleost fish. Biochim. Biophys. Acta Gen Subj. 2019, 1863, 39–51. [Google Scholar] [CrossRef]
- Karsenty, G.; Ferron, M. The contribution of bone to whole-organism physiology. Nature 2012, 481, 314–320. [Google Scholar] [CrossRef]
- Alisi, L.; Cao, R.; De Angelis, C.; Cafolla, A.; Caramia, F.; Cartocci, G.; Librando, A.; Fiorelli, M. The relationships between vitamin k and cognition: A review of current evidence. Front. Neurol. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Hirota, Y.; Suhara, Y. New aspects of vitamin K research with synthetic ligands: Transcriptional activity via SXR and neural differentiation activity. Int. J. Mol. Sci. 2019, 20, 3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saghiri, M.A.; Asatourian, A.; Ershadifar, S.; Moghadam, M.M.; Sheibani, N. Vitamins and regulation of angiogenesis: [A, B1, B2, B3, B6, B9, B12, C, D, E, K]. J. Funct. Foods 2017, 38, 180–196. [Google Scholar] [CrossRef]
- Shirakawa, H.; Ohsaki, Y.; Minegishi, Y.; Takumi, N.; Ohinata, K.; Furukawa, Y.; Mizutani, T.; Komai, M. Vitamin K deficiency reduces testosterone production in the testis through down-regulation of the Cyp11a a cholesterol side chain cleavage enzyme in rats. Biochim. Biophys. Acta. 2006, 1760, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Shirakawa, H.; Takumi, N.; Minegishi, Y.; Ohashi, A.; Howlader, Z.H.; Ohsaki, Y.; Sato, T.; Goto, T.; Komai, M. Menaquinone-4 enhances testosterone production in rats and testis-derived tumor cells. Lipids Health Dis. 2011, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, I.; Vijayakumar, P.; Marques, C.; Cancela, M.L.; Gavaia, P.J.; Laizé, V. Zebrafish vitamin K epoxide reductases: Expression in vivo, along extracellular matrix mineralization and under phylloquinone and warfarin in vitro exposure. Fish Physiol. Biochem. 2015, 41, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Bevans, C.G.; Krettler, C.; Reinhart, C.; Watzka, M.; Oldenburg, J. Phylogeny of the vitamin K 2,3-Epoxide Reductase (VKOR) family and evolutionary relationship to the disulfide bond formation protein B (DsbB) family. Nutrients 2015, 7, 6224–6249. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, J.; Watzka, M.; Bevans, C.G. VKORC1 and VKORC1L1: Why do vertebrates have two vitamin K 2,3-epoxide reductases? Nutrients 2015, 7, 6250–6280. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.D.; Betts, L.; Talmage, G.; Pollet, R.M.; Holman, N.S.; Redinbo, M.R. Structural and functional analysis of the human nuclear xenobiotic receptor PXR in complex with RXRa. J. Mol. Biol. 2013, 425, 2561–2577. [Google Scholar] [CrossRef] [Green Version]
- Hammed, A.; Matagrin, B.; Spohn, G.; Prouillac, C.; Benoit, E.; Lattard, V. VKORC1L1, an enzyme rescuing the vitamin K 2,3-Epoxide reductase activity in some extrahepatic tissues during anticoagulation therapy. J. Biol. Chem. 2013, 288, 28733–28742. [Google Scholar] [CrossRef] [Green Version]
- Rishavy, M.A.; Hallgren, K.W.; Wilson, L.A.; Usubalieva, A.; Runge, K.W.; Berkner, K.L. The vitamin K oxidoreductase is a multimer that efficiently reduces vitamin k epoxide to hydroquinone to allow vitamin K-dependent protein carboxylation. J. Biol. Chem. 2013, 288, 31556–31566. [Google Scholar] [CrossRef] [Green Version]
- Oldenburg, J.; Marinova, M.; Müller-Reible, C.; Watzka, M. The vitamin K cycle. Vitam. Horm. 2008, 78, 35–62. [Google Scholar] [CrossRef]
- Tie, J.K.; Stafford, D.W. Structural and functional insights into enzymes of the vitamin K cycle. J. Thromb. Haemost. 2016, 14, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Kulman, J.D.; Harris, J.E.; Nakazawa, N.; Ogasawara, M.; Satake, M.; Davie, E.W. Vitamin K-dependent proteins in Ciona intestinalis, a basal chordate lacking a blood coagulation cascade. Proc. Natl. Acad. Sci. USA 2006, 103, 15794–15799. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Yang, C.T.; Jin, D.; Stafford, D.W. Identification of a drosophila vitamin K-dependent γ-Glutamyl Carboxylase. J. Biol. Chem. 2000, 275, 18291–18296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandyopadhyay, P.K.; Garrett, J.E.; Shetty, R.P.; Keate, T.; Walker, C.S.; Olivera, B.M. γ-Glutamyl Carboxylation: An extracellular posttranslational modification that antedates the divergence of molluscs, arthropods, and chordates. Proc. Natl. Acad. Sci. USA 2002, 99, 1264–1269. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Chen, J.; Duan, L.; Li, S. Vitamin K-Dependent proteins involved in bone and cardiovascular health. Mol. Med. Rep. 2018, 18, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Spohn, G.; Kleinridders, A.; Wunderlich, F.T.; Watzka, M.; Zaucke, F.; Blumbach, K.; Geisen, C.; Seifried, E.; Müller, C.; Paulsson, M.; et al. VKORC1 deficiency in mice causes early postnatal lethality due to severe bleeding. Thromb. Haemost. 2009, 101, 1044–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatron, N.; Chalmond, B.; Trouvé, A.; Benoît, E.; Caruel, H.; Lattard, V.; Tchertanov, L. Identification of the functional states of human vitamin K epoxide reductase from molecular dynamics simulations. RSC Adv. 2017, 7, 52071–52090. [Google Scholar] [CrossRef] [Green Version]
- Willson, T.M.; Kliewer, S.A. PXR, CAR and drug metabolism. Nat. Rev. Drug Discov. 2002, 1, 259–266. [Google Scholar] [CrossRef]
- Ekins, S.; Reschly, E.J.; Hagey, L.R.; Krasowski, M.D. Evolution of pharmacologic specificity in the pregnane X receptor. BMC Evol. Pharmacol. 2008, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Salanga, M.C.; Brun, N.R.; Francolini, R.D.; Stegeman, J.J.; Goldstone, J.V. CRISPR-Cas9-Mutated pregnane X receptor (pxr) retains pregnenolone-induced expression of cyp3a65 in Zebrafish (Danio rerio) larvae. Toxicol. Sci. 2020, 174, 51–62. [Google Scholar] [CrossRef]
- Cui, W.; Sun, M.; Zhang, S.; Shen, X.; Galeva, N.; Williams, T.D.; Staudinger, J.L.A. SUMO-Acetyl switch in PXR biology. Biochim. Biophys. Acta - Gene Regul. Mech. 2016, 1859, 1170–1182. [Google Scholar] [CrossRef] [Green Version]
- Rulcova, A.; Prokopova, I.; Krausova, L.; Bitman, M.; Vrzal, R.; Dvorak, Z.; Blahos, J.; Pavek, P. Stereoselective interactions of warfarin enantiomers with the pregnane X nuclear receptor in gene regulation of major drug-metabolizing cyto- chrome P450 enzymes. J. Thromb. Haemost. 2010, 8, 2708–2717. [Google Scholar] [CrossRef]
- Cui, J.Y.; Klaassen, C.D. RNA-Seq reveals common and unique PXR- and CAR-Target gene signatures in the mouse liver transcriptome. Biochim. Biophys. Acta - Gene Regul. Mech. 2016, 1859, 1198–1217. [Google Scholar] [CrossRef] [Green Version]
- Pavek, P. Pregnane X receptor (PXR)-mediated gene repression and cross-talk of PXR with other nuclear receptors via coactivator interactions. Front. Pharmacol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Vergara, P.; Ho, Y.S.; Espinoza, F.; Nualart, F.; Escudero, C.; Penny, J. The constitutive androstane receptor and pregnane X receptor in the brain. Br. J. Pharmacol. 2020, 177, 2666–2682. [Google Scholar] [CrossRef]
- Bertrand, S.; Thisse, B.; Tavares, R.; Sachs, L.; Chaumot, A.; Bardet, P.; Escrivà, H.; Duffraisse, M.; Marchand, O.; Safi, R.; et al. Unexpected novel relational links uncovered by extensive developmental profiling of nuclear receptor expression. PLoS Genet. 2007, 3, e188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gräns, J.; Wassmur, B.; Fernández-Santoscoy, M.; Zanette, J.; Woodin, B.R.; Karchner, S.I.; Nacci, D.E.; Champlin, D.; Jayaraman, S.; Hahn, M.E.; et al. Regulation of pregnane-X-receptor, CYP3A and P-glycoprotein genes in the PCB-resistant killifish (Fundulus heteroclitus) population from New Bedford Harbor. Aquat. Toxicol. 2015, 159, 198–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bainy, A.C.D.; Kubota, A.; Goldstone, J.V.; Lille-Langøy, R.; Karchner, S.I.; Celander, M.C.; Hahn, M.E.; Goksøyr, A.; Stegeman, J.J. Functional characterization of a full length pregnane X receptor, expression in vivo, and identification of PXR alleles, in zebrafish (Danio Rerio). Aquat. Toxicol. 2013, 142–143, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornbluth, S.; Fissore, R. Vertebrate reproduction. Cold Spring Harb. Perspect. Biol. 2015, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, G.; Carnevali, O.; Franzoni, M.F.; Cottone, E.; Lutz, I.; Kloas, W.; Yamamoto, K.; Kikuyama, S.; Polzonetti-Magni, A.M. Environmental estrogens and reproductive biology in amphibians. Gen. Comp. Endocrinol. 2002, 126, 125–129. [Google Scholar] [CrossRef]
- Giesy, J.P.; Lori, A.F.; Jones, P.D.; Kannan, K.; Sanderson, T. Review of the effects of endocrine-disrupting chemicals in birds. Pure Appl. Chem. 2003, 75, 2287–2303. [Google Scholar] [CrossRef]
- Hong, J.; Chen, F.; Wang, X.; Bai, Y.; Zhou, R.; Li, Y.; Chen, L. Exposure of preimplantation embryos to low-dose bisphenol A impairs testes development and suppresses histone acetylation of StAR promoter to reduce production of testosterone in mice. Mol. Cell. Endocrinol. 2016, 427, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Forner-Piquer, I.; Beato, S.; Piscitelli, F.; Santangeli, S.; Di Marzo, V.; Habibi, H.R.; Maradonna, F.; Carnevali, O. Effects of BPA on zebrafish gonads: Focus on the endocannabinoid system. Environ. Pollut. 2020, 264. [Google Scholar] [CrossRef] [PubMed]
- Almawi, W.Y.; Al-Shaikh, F.S.; Melemedjian, O.K.; Almawi, A.W. Protein Z, an anticoagulant protein with expanding role in reproductive biology. Reproduction 2013, 146, R73–R80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofikitis, N.; Giotitsas, N.; Tsounapi, P.; Baltogiannis, D.; Giannakis, D.; Pardalidis, N. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol. 2008, 109, 323–330. [Google Scholar] [CrossRef]
- Schulz, R.W.; De França, L.R.; Lareyre, J.J.; Le Gac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef]
- Chen, S.R.; Liu, Y.X. Regulation of spermatogonial stem cell self-renewal and spermatocyte meiosis by sertoli cell signaling. Reproduction 2015, 149, R159–R167. [Google Scholar] [CrossRef] [Green Version]
- Tovo-Neto, A.; Da Silva Rodrigues, M.; Habibi, H.R.; Nóbrega, R.H. Thyroid hormone actions on male reproductive system of teleost fish. Gen. Comp. Endocrinol. 2018, 265, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.B.; Walker, W.H. The regulation of spermatogenesis by androgens. Semin. Cell Dev. Biol. 2014, 30, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Caspers, M.; Czogalla, K.J.; Liphardt, K.; Müller, J.; Westhofen, P.; Watzka, M.; Oldenburg, J. Two enzymes catalyze vitamin K 2,3-Epoxide reductase activity in mouse: VKORC1 is highly expressed in exocrine tissues while VKORC1L1 is highly expressed in brain. Thromb. Res. 2015, 135, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Brewer, C.T.; Chen, T. PXR Variants: The impact on drug metabolism and therapeutic responses. Acta Pharm. Sin. B 2016, 6, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Wassmur, B.; Gräns, J.; Kling, P.; Celander, M.C. Interactions of pharmaceuticals and other xenobiotics on hepatic pregnane X receptor and cytochrome P450 3A signaling pathway in rainbow trout (Oncorhynchus mykiss). Aquat Toxicol. 2010, 100, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, I.; Gibson, G.G.; Plant, K.; Graham, M.; Plant, N. A PXR-Mediated negative feedback loop attenuates the expression of CYP3A in response to the PXR agonist pregnenalone-16α-Carbonitrile. PLoS ONE 2011, 6, e16703. [Google Scholar] [CrossRef] [Green Version]
- Frye, C.A.; Koonce, C.J.; Walf, A.A. Novel receptor targets for production and action of allopregnanolone in the central nervous system: A focus on pregnane xenobiotic receptor. Front. Cell. Neurosci. 2014, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihunnah, C.A.; Jiang, M.; Xie, W. Nuclear receptor PXR, transcriptional circuits and metabolic relevance. Biochim. Biophys. Acta-Mol. Basis Dis. 2011, 1812, 956–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, T.; Yabusaki, Y.; Honma, K.; Matsuo, N.; Tatsuta, K.; Ishibashi, F.; Katagiri, M. Contribution of human hepatic cytochrome P450 isoforms to regioselective hydroxylation of steroid hormones. Xenobiotica 1998, 28, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.A.; Squires, E.J. Effects of nuclear receptor transactivation on boar taint metabolism and gene expression in porcine hepatocytes. J. Steroid Biochem. Mol. Biol. 2013, 133, 110–119. [Google Scholar] [CrossRef]
- Sanyaolu, A.O.; Oremosu, A.A.; Osinubi, A.A.; Vermeer, C.; Daramola, A.O. Warfarin-induced vitamin K deficiency affects spermatogenesis in Sprague–Dawley rats. Andrologia 2019, 51, 1–9. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Yuan, J.; Feng, L.; Jiang, W.D.; Liu, Y.; Jiang, J.; Li, S.H.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Effects of dietary vitamin K levels on growth performance, enzyme activities and antioxidant status in the hepatopancreas and intestine of juvenile jian carp (Cyprinus Carpio Var. Jian). Aquac. Nutr. 2016, 22, 352–366. [Google Scholar] [CrossRef]
- Czogalla, K.J.; Liphardt, K.; Höning, K.; Hornung, V.; Biswas, A.; Watzka, M.; Oldenburg, J. VKORC1 and VKORC1L1 have distinctly different oral anticoagulant dose-response characteristics and binding sites. Blood Adv. 2018, 2, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Kotaja, N. MicroRNAs and spermatogenesis. Fertil. Steril. 2014, 101, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Machtinger, R.; Laurent, L.C.; Baccarelli, A.A. Extracellular vesicles: Roles in gamete maturation, fertilization and embryo implantation. Hum. Reprod. Update 2016, 22, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Edson, M.A.; Nagaraja, A.K.; Matzuk, M.M. The mammalian ovary from genesis to revelation. Endocr. Rev. 2009, 30, 624–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoldo, M.J.; Walters, K.A.; Ledger, W.L.; Gilchrist, R.B.; Mermillod, P.; Locatelli, Y. In-vitro regulation of primordial follicle activation: Challenges for fertility preservation strategies. Reprod. Biomed. Online 2018, 36, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fang, Y.; Liu, Y.; Yang, X. MicroRNAs in ovarian function and disorders. J. Ovarian Res. 2015, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luderer, U. Ovarian toxicity from reactive oxygen species. Vitam. Horm. 2014, 94, 99–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, J.L.; Tilly, K.I. Inhibitors of oxidative stress mimic the ability of follicle-stimulating hormone to suppress apoptosis in cultured rat ovarian follicles. Endocrinology 1995, 136, 242–252. [Google Scholar] [CrossRef]
- Burstyn-Cohen, T. TAM receptor signaling in development. Int. J. Dev. Biol. 2017, 61, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Saedi, S.; Khoradmehr, A.; Mohammad Reza, J.S.; Tamadon, A. The role of neuropeptides and neurotransmitters on Kisspeptin/Kiss1r-Signaling in female reproduction. J. Chem. Neuroanat. 2018, 92, 71–82. [Google Scholar] [CrossRef]
- Stamatiades, G.A.; Kaiser, U.B. Gonadotropin regulation by pulsatile GnRH: Signaling and gene expression. Mol. Cell Endocrinol. 2018, 463, 131–141. [Google Scholar] [CrossRef]
- Fallah, H.P.; Rodrigues, M.S.; Corchuelo, S.; Nóbrega, R.H.; Habibi, H.R. Role of GnRH isoforms in paracrine/autocrine control of zebrafish (Danio rerio) spermatogenesis. Endocrinology 2020, 161, bqaa004. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine leydig cells. Steroids 2015, 103, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.H.; Cheng, J. FSH and testosterone signaling in sertoli cells. Reproduction 2005, 130, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilbrook, A.J.; Clarke, I.J. Negative feedback regulation of the secretion and actions of gonadotropin-releasing hormone in males. Biol. Reprod. 2001, 64, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Chimento, A.; Sirianni, R.; Casaburi, I.; Pezzi, V. Role of estrogen receptors and G protein-coupled estrogen receptor in regulation of hypothalamus-pituitary-testis axis and spermatogenesis. Front. Endocrinol. 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Iwasa, T.; Matsuzaki, T.; Yano, K.; Yanagihara, R.; Mayila, Y.; Irahara, M. The effects of chronic testosterone administration on hypothalamic gonadotropin-releasing hormone regulatory factors (Kiss1, NKB, pDyn and RFRP) and their receptors in female rats. Gynecol. Endocrinol. 2017, 34, 437–441. [Google Scholar] [CrossRef]
- He, W.; Li, X.; Adekunbi, D.; Liu, Y.; Long, H.; Wang, L.; Lyu, Q.; Kuang, Y.; O’Byrne, K.T. Hypothalamic effects of progesterone on regulation of the pulsatile and surge release of luteinising hormone in female rats. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Rodriguez, A.; Kauffman, A.S.; Cherrington, B.D.; Borges, C.S.; Roepke, T.A.; Laconi, M. Emerging insights into hypothalamic-pituitary-gonadal axis regulation and interaction with stress signalling. J. Neuroendocrinol. 2018, 30, e12590. [Google Scholar] [CrossRef]
- Kirilov, M.; Clarkson, J.; Liu, X.; Roa, J.; Campos, P.; Porteous, R.; Schütz, G.; Herbison, A.E. Dependence of fertility on kisspeptin-Gpr54 signaling at the GnRH neuron. Nat Commun. 2013, 4, 2492. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Shao, R.; Weijdegård, B.; Wang, T.; Johansson, J.; Sun, S.; Wang, W.; Egecioglu, E.; Billig, H.; Stener-Victorin, E. Effects of androgen and leptin on behavioral and cellular responses in female rats. Horm. Behav. 2011, 60, 427–438. [Google Scholar] [CrossRef]
- Venancio, J.C.; Margatho, L.O.; Rorato, R.; Rosales, R.R.C.; Debarba, L.K.; Coletti, R.; Antunes-Rodrigues, J.; Elias, C.F.; Elias, L.L.K. Short-term high-fat diet increases leptin activation of CART neurons and advances puberty in female mice. Endocrinology 2017, 158, 3929–3942. [Google Scholar] [CrossRef] [PubMed]
- Shutter, J.R.; Graham, M.; Kinsey, A.C.; Scully, S.; Lüthy, R.; Stark, K.L. Hypothalamic expression of ART, a novel gene related to agouti, is up-regulated in obese and diabetic mutant mice. Genes Dev. 1997, 11, 593–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, S.L.; Qiu, J.; Nestor, C.C.; Zhang, C.; Smith, A.W.; Whiddon, B.B.; Rønnekleiv, O.K.; Kelly, M.J.; Palmiter, R.D. AgRP to Kiss1 neuron signaling links nutritional state and fertility. Proc. Natl. Acad. Sci. USA 2017, 114, 2413–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque, R.M.; Kineman, R.D.; Tena-Sempere, M. Regulation of hypothalamic expression of KiSS-1 and GPR54 genes by metabolic factors: Analyses using mouse models and a cell line. Endocrinology 2007, 148, 4601–4611. [Google Scholar] [CrossRef] [PubMed]
- Topaloglu, A.K.; Reimann, F.; Guclu, M.; Yalin, A.S.; Kotan, L.D.; Porter, K.M.; Serin, A.; Mungan, N.O.; Cook, J.R.; Ozbek, M.N.; et al. TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for neurokinin b in the central control of reproduction. Nat. Genet. 2009, 41, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Min, L.; Adeola, O.; Carroll, R.S.; Kaiser, U.B. Glutamate acts as a cofactor in the activation of KISS1R by kisspeptin. In Signaling Originating from Membrane Receptors; Endocrine Society: San Francisco, CA, USA, 2013; Volume 3, p. SUN–402. [Google Scholar]
- Lehman, M.N.; Merkley, C.M.; Coolen, L.M.; Goodman, R.L. Anatomy of the kisspeptin neural network in mammals. Brain Res. 2010, 1364, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo-Lopes, R.; Crampton, J.R.; Aquino, N.S.; Miranda, R.M.; Kokay, I.C.; Reis, A.M.; Franci, C.R.; Grattan, D.R.; Szawka, R.E. Prolactin regulates kisspeptin neurons in the arcuate nucleus to suppress LH secretion in female rats. Endocrinology 2014, 155, 1010–1020. [Google Scholar] [CrossRef] [Green Version]
- Moenter, S.M.; Chu, Z.; Christian, C.A. Neurobiological mechanisms underlying oestradiol negative and positive feedback regulation of gonadotrophin-releasing hormone neurones. J. Neuroendocrinol. 2009, 21, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Ciechanowska, M.; Łapot, M.; Malewski, T.; Mateusiak, K.; Misztal, T.; Przekop, F. Effects of GABAA receptor modulation on the expression of GnRH gene and GnRH receptor (GnRH-R) gene in the hypothalamus and GnRH-R gene in the anterior pituitary gland of follicular-phase ewes. Anim. Reprod. Sci. 2009, 111, 235–248. [Google Scholar] [CrossRef]
- Pielecka-Fortuna, J.; Moenter, S.M. Kisspeptin increases γ-Aminobutyric acidergic and glutamatergic transmission directly to gonadotropin-releasing hormone neurons in an estradiol-dependent manner. Endocrinology 2010, 151, 291–300. [Google Scholar] [CrossRef]
- Herbison, A.E.; Moenter, S.M. Depolarising and hyperpolarising actions of GABAA receptor activation on gonadotrophin-releasing hormone neurones: Towards an emerging consensus. J. Neuroendocrinol. 2011, 23, 557–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spruston, N.; McBain, C.J. Structural and functional properties of hippocampal neurons. In The Hippocampus Book; Andersen, P., Morris, R., Amaral, D., Bliss, T., O’Keefe, J., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 1–852. [Google Scholar]
- Fallah, H.P.; Tovo-Neto, A.; Yeung, E.C.; Nóbrega, R.H.; Habibi, H.R. Paracrine/Autocrine control of spermatogenesis by gonadotropin-inhibitory hormone. Mol. Cell. Endocrinol. 2019, 492, 110440. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, J.P.; Roa, J.; Herbison, A.E.; Han, S.K. Serotonin acts through 5-HT1 and 5-HT2 receptors to exert biphasic actions on GnRH neuron excitability in the mouse. Endocrinology 2014, 155, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.M.; McMaster, C.R. PC and PE synthesis: Mixed micellar analysis of the cholinephosphotransferase and ethanolaminephosphotransferase activities of human choline/ethanolamine phosphotransferase 1 (CEPT1). Lipids 2002, 37, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Reue, K. Lipin proteins and glycerolipid metabolism: Roles at the ER membrane and beyond. Biochim. Biophys. Acta - Biomembr. 2017, 1859, 1583–1595. [Google Scholar] [CrossRef]
- Law, N.C.; Weck, J.; Kyriss, B.; Nilson, J.H.; Hunzicker-Dunn, M. Lhcgr expression in granulosa cells: Roles for PKA-phosphorylated β-catenin, TCF3, and FOXO1. Mol. Endocrinol. 2013, 27, 1295–1310. [Google Scholar] [CrossRef] [Green Version]
- Palermo, R. Differential actions of FSH and LH during folliculogenesis. Reprod. Biomed. Online 2007, 15, 326–337. [Google Scholar] [CrossRef]
- Ortiga-Carvalho, T.M.; Chiamolera, M.I.; Pazos-Moura, C.C.; Wondisford, F.E. Hypothalamus-pituitary-thyroid axis. Compr. Physiol. 2016, 6, 1387–1428. [Google Scholar] [CrossRef]
- Gogakos, A.I.; Duncan-Bassett, J.H.; Williams, G.R. Thyroid and bone. Arch. Biochem. Biophys. 2010, 503, 129–136. [Google Scholar] [CrossRef]
- Robbins, J. Thyroid hormone transport proteins and the physiology of hormone binding. In Werner and Ingbar’s, The Thyroid, 8th ed.; Braverman, L.E., Utiger, R.D., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000; pp. 105–120. [Google Scholar]
- Choksi, N.Y.; Jahnke, G.D.; St Hilaire, C.; Shelby, M. Role of thyroid hormones in human and laboratory animal reproductive health. Birth Defects Res. B Dev. Reprod. Toxicol. 2003, 68, 479–491. [Google Scholar] [CrossRef]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Thyroid hormones and female reproduction. Biol. Reprod. 2018, 99, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, D.C.; Langlois, V.S.; Fernandino, J.I. Crossover of the hypothalamic pituitary-adrenal/interrenal,-thyroid, and -gonadal axes in testicular development. Front. Endocrinol. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dutt, M.; Wehrle, C.J.; Jialal, I. Physiology, Adrenal Gland. In StatPearls. Treasure Island (FL): StatPearls. Available online: https://www.ncbi.nlm.nih.gov/books/NBK537260/ (accessed on 18 July 2020).
- Majzoub, J.A. Corticotropin-releasing hormone physiology. Eur. J. Endocrinol. Suppl. 2006, 155, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.D.; O’Byrne, K.T.; Chiappini, S.E.; Hotchkiss, J.; Knobil, E. Hypoglycemic ‘stress’ and gonadotropin-releasing hormone pulse generator activity in the rhesus monkey: Role of the ovary. Neuroendocrinology 1992, 56, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Yates, R.; Katugampola, H.; Cavlan, D.; Cogger, K.; Meimaridou, E.; Hughes, C.; Metherell, L.; Guasti, L.; King, P. Adrenocortical development, maintenance, and disease. In Current Topics in Developmental Biology, 1st ed.; Thomas, P., Ed.; Elsevier: London, UK, 2013; Volume 106, pp. 239–312. [Google Scholar]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids, stress, and fertility. Minerva Endocrinol. 2010, 35, 109–125. [Google Scholar]
- Kalantaridou, S.N.; Zoumakis, E.; Makrigiannakis, A.; Lavasidis, L.G.; Vrekoussis, T.; Chrousos, G.P. Corticotropin-releasing hormone, stress and human reproduction: An Update. J. Reprod. Immunol. 2010, 85, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Oyola, M.G.; Handa, R.J. Hypothalamic–pituitary–adrenal and hypothalamic–pituitary–gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- Hassani-Nezhad-Gashti, F.; Kummu, O.; Karpale, M.; Rysä, J.; Hakkola, J. Nutritional status modifies pregnane X receptor regulated transcriptome. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Pai, H.V.; Zhou, J.; Amico, J.A.; Vollmer, R.R.; Xie, W. Activation of pregnane X receptor disrupts glucocorticoid and mineralocorticoid homeostasis. Mol. Endocrinol. 2007, 21, 138–147. [Google Scholar] [CrossRef]
- Tokarz, V.L.; MacDonald, P.E.; Klip, A. The cell biology of systemic insulin function. J. Cell Biol. 2018, 217, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Starup-Linde, J.; Gregersen, S.; Frost, M.; Vestergaard, P. Use of glucose-lowering drugs and risk of fracture in patients with type 2 diabetes. Bone 2017, 95, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunnerhagen, M.; Drakenberg, T.; Forsen, S.; Stenflo, J. Effect of Ca2+ on the structure of vitamin K-dependent coagulation factors. Haemostasis 1996, 26, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Tangseefa, P.; Martin, S.K.; Fitter, S.; Baldock, P.A.; Proud, C.G.; Zannettino, A.C.W. Osteocalcin-dependent regulation of glucose metabolism and fertility: Skeletal implications for the development of insulin resistance. J. Cell. Physiol. 2018, 233, 3769–3783. [Google Scholar] [CrossRef]
- Oury, F.; Sumara, G.; Sumara, O.; Ferron, M.; Chang, H.; Smith, C.E.; Hermo, L.; Suarez, S.; Roth, B.L.; Ducy, P.; et al. Endocrine regulation of male fertility by the skeleton. Cell 2011, 144, 796–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oury, F.; Ferron, M.; Huizhen, W.; Confavreux, C.; Xu, L.; Lacombe, J.; Srinivas, P.; Chamouni, A.; Lugani, F.; Lejeune, H.; et al. Osteocalcin regulates murine and human fertility through a pancreas-bone-testis axis. J. Clin. Investig. 2013, 123, 2421–2433. [Google Scholar] [CrossRef] [PubMed]
- De Toni, L.; Guidolin, D.; De Filippis, V.; Peterle, D.; Rocca, M.S.; Di Nisio, A.; De Rocco Ponce, M.; Foresta, C. SHBG141–161 domain-peptide stimulates GPRC6A-mediated response in leydig and β-Langerhans cell lines. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Rossi, M.; Battafarano, G.; Pepe, J.; Minisola, S.; Del Fattore, A. The endocrine function of osteocalcin regulated by bone resorption: A lesson from reduced and increased bone mass diseases. Int. J. Mol. Sci. 2019, 20, 4502. [Google Scholar] [CrossRef] [Green Version]
- Coskun, G.; Sencar, L.; Tuli, A.; Saker, D.; Alparslan, M.M.; Polat, S.; Silvestrini, A. Effects of osteocalcin on synthesis of testosterone and INSL3 during adult leydig cell differentiation. Int. J. Endocrinol. 2019, 2019, 1–18. [Google Scholar] [CrossRef]
- Spruiell, K.; Gyamfi, A.A.; Yeyeodu, S.T.; Richardson, R.M.; Gonzalez, F.J.; Gyamfi, M.A. Pregnane X receptor-humanized mice recapitulate gender differences in ethanol metabolism but not hepatotoxicity. J. Pharmacol. Exp. Ther. 2015, 354, 459–470. [Google Scholar] [CrossRef]
- Taborska, E.; Pasulka, J.; Malik, R.; Horvat, F.; Jenickova, I.; Matošević, Z.J.; Svoboda, P. Restricted and non-essential redundancy of RNAi and PiRNA pathways in mouse oocytes. PLoS Genet. 2019, 15, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.H.; Li, Y.; Deng, S.L.; Liu, Y.X.; Lian, Z.X.; Yu, K. Recent research advances in mitosis during mammalian gametogenesis. Cells 2019, 8, 567. [Google Scholar] [CrossRef] [Green Version]
- Skaftnesmo, K.O.; Edvardsen, R.B.; Furmanek, T.; Crespo, D.; Andersson, E.; Kleppe, L.; Taranger, G.L.; Bogerd, J.; Schulz, R.W.; Wargelius, A. Integrative testis transcriptome analysis reveals differentially expressed miRNAs and their mRNA targets during early puberty in Atlantic salmon. BMC Genom. 2017, 18, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Jenkins, T.G.; Carrell, D.T. Diet and sperm quality: Nutrients, foods and dietary patterns. Reprod. Biol. 2019, 19, 219–224. [Google Scholar] [CrossRef]
- Tong, M.H.; Mitchell, D.; Evanoff, R.; Griswold, M.D. Expression of Mirlet7 family microRNAs in response to retinoic acid-induced spermatogonial differentiation in mice. Biol. Reprod. 2011, 85, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Huszar, J.M.; Payne, C.J. MicroRNA 146 (Mir146) modulates spermatogonial differentiation by retinoic acid in mice. Biol. Reprod. 2013, 15, 1–10. [Google Scholar] [CrossRef]
- Lanham, S.A.; Cagampang, F.R.; Oreffo, R.O.C. Maternal high fat diet and offspring expression levels of vitamin K-dependent proteins. Endocrinology 2014, 155, 4749–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beato, S.; Toledo-Solís, F.J.; Fernández, I. Vitamin K in Vertebrates’ Reproduction: Further Puzzling Pieces of Evidence from Teleost Fish Species. Biomolecules 2020, 10, 1303. https://doi.org/10.3390/biom10091303
Beato S, Toledo-Solís FJ, Fernández I. Vitamin K in Vertebrates’ Reproduction: Further Puzzling Pieces of Evidence from Teleost Fish Species. Biomolecules. 2020; 10(9):1303. https://doi.org/10.3390/biom10091303
Chicago/Turabian StyleBeato, Silvia, Francisco Javier Toledo-Solís, and Ignacio Fernández. 2020. "Vitamin K in Vertebrates’ Reproduction: Further Puzzling Pieces of Evidence from Teleost Fish Species" Biomolecules 10, no. 9: 1303. https://doi.org/10.3390/biom10091303
APA StyleBeato, S., Toledo-Solís, F. J., & Fernández, I. (2020). Vitamin K in Vertebrates’ Reproduction: Further Puzzling Pieces of Evidence from Teleost Fish Species. Biomolecules, 10(9), 1303. https://doi.org/10.3390/biom10091303