
Identification of TSPAN4 as Novel Histamine H4 Receptor Interactor


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
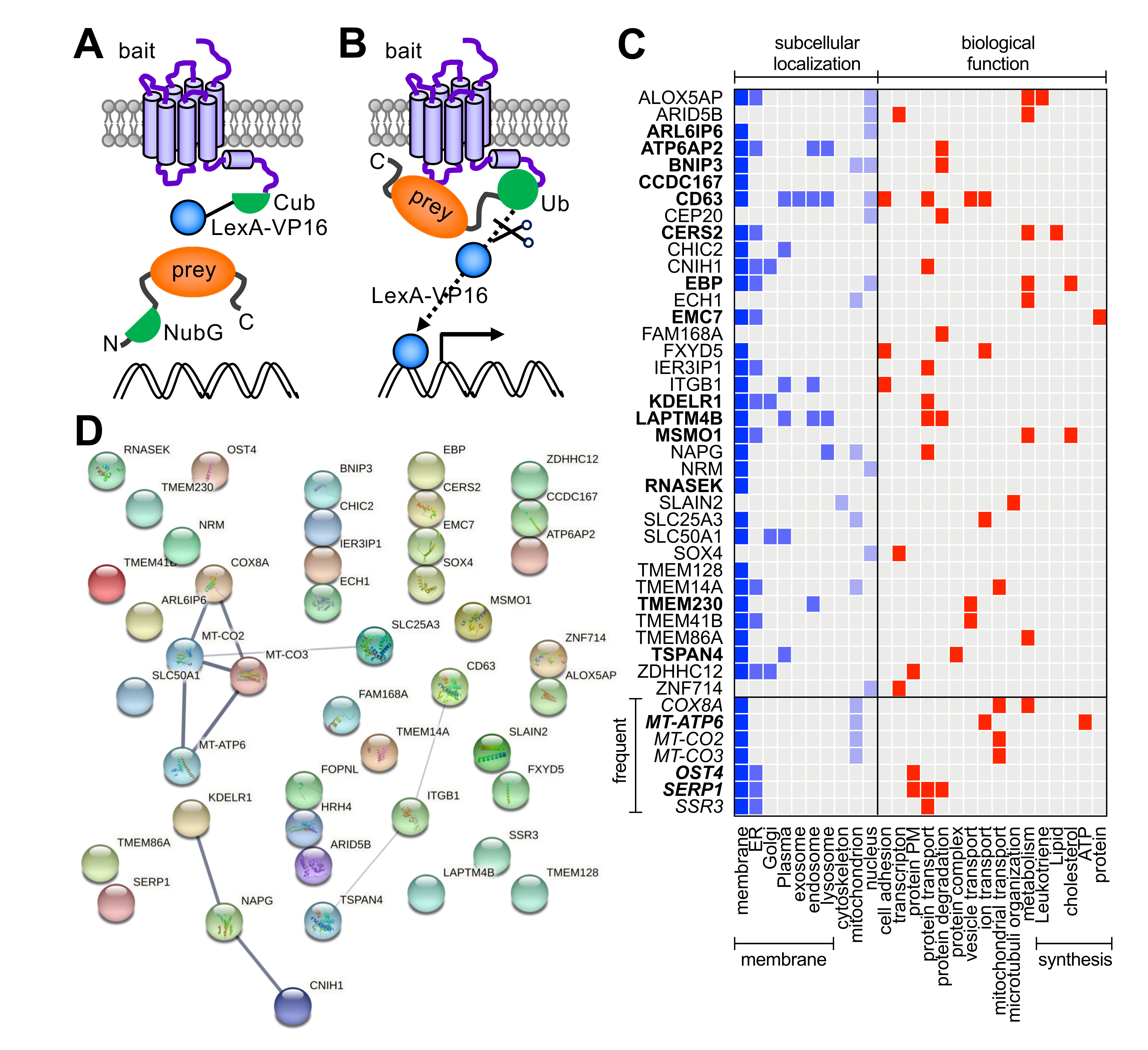
2.2. MYTH Constructs and Screen
2.3. MYTH Strains and Growth Conditions
2.4. Cell Culture and Transfection
2.5. Mammalian Expression Constructs
2.6. Bioluminescence Resonance Energy Transfer (BRET)-Based Close Proximity Detection
2.7. Biomolecular Fluorescence Complementation (BiFC)-Based Close Proximity Detection
2.8. Co-Immunoprecipitation
2.9. [3H]Histamine Binding Assay
2.10. BRET-Based G Protein Activation Biosensor
2.11. Data Analysis
3. Results
3.1. Identification of H4R Interactors by MYTH Screen of Jurkat T Cell cDNA Library
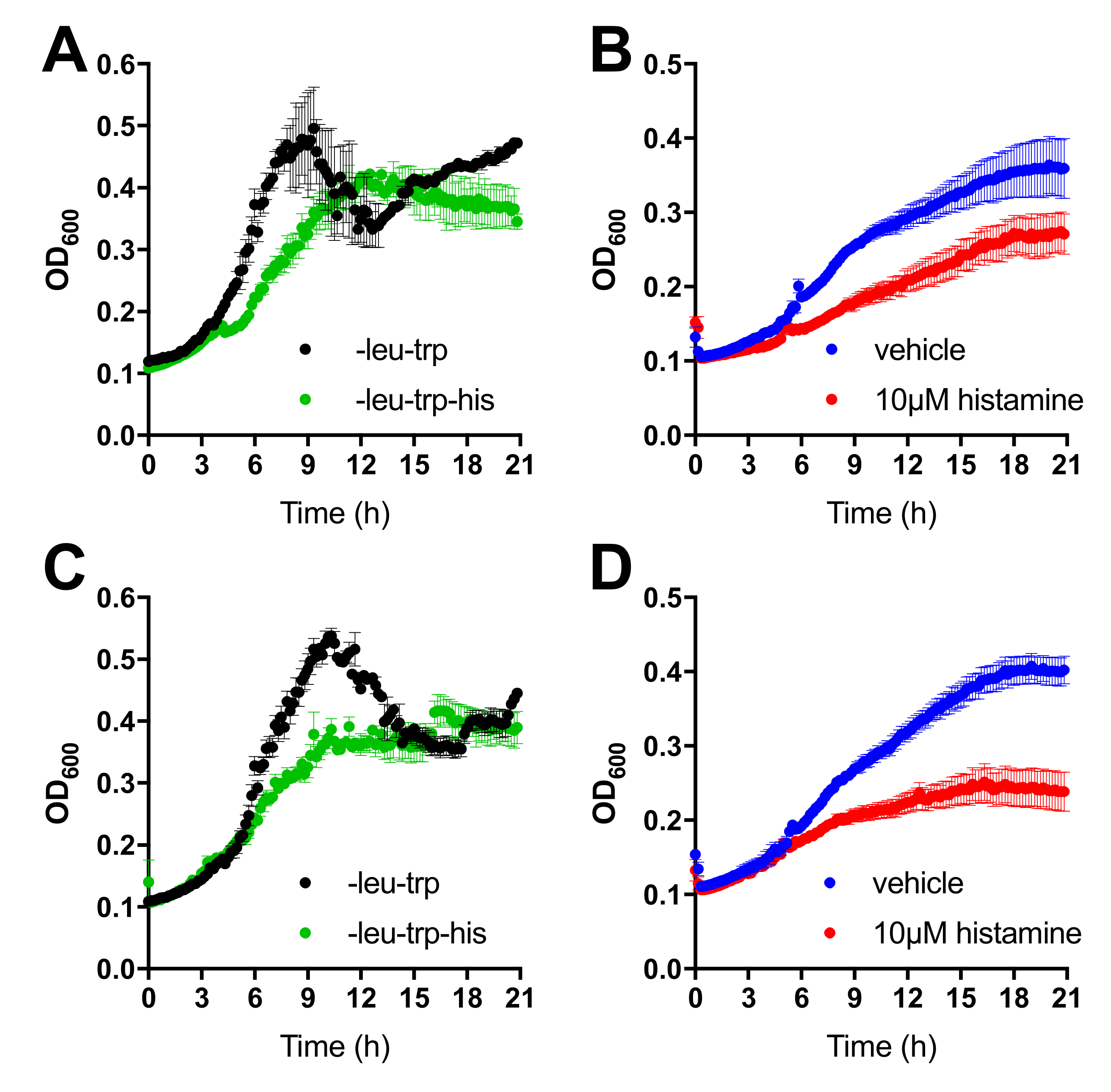
3.2. MYTH between H4R and Tetraspanins TSPAN4 and CD63 Is Decreased by Histamine Stimulation
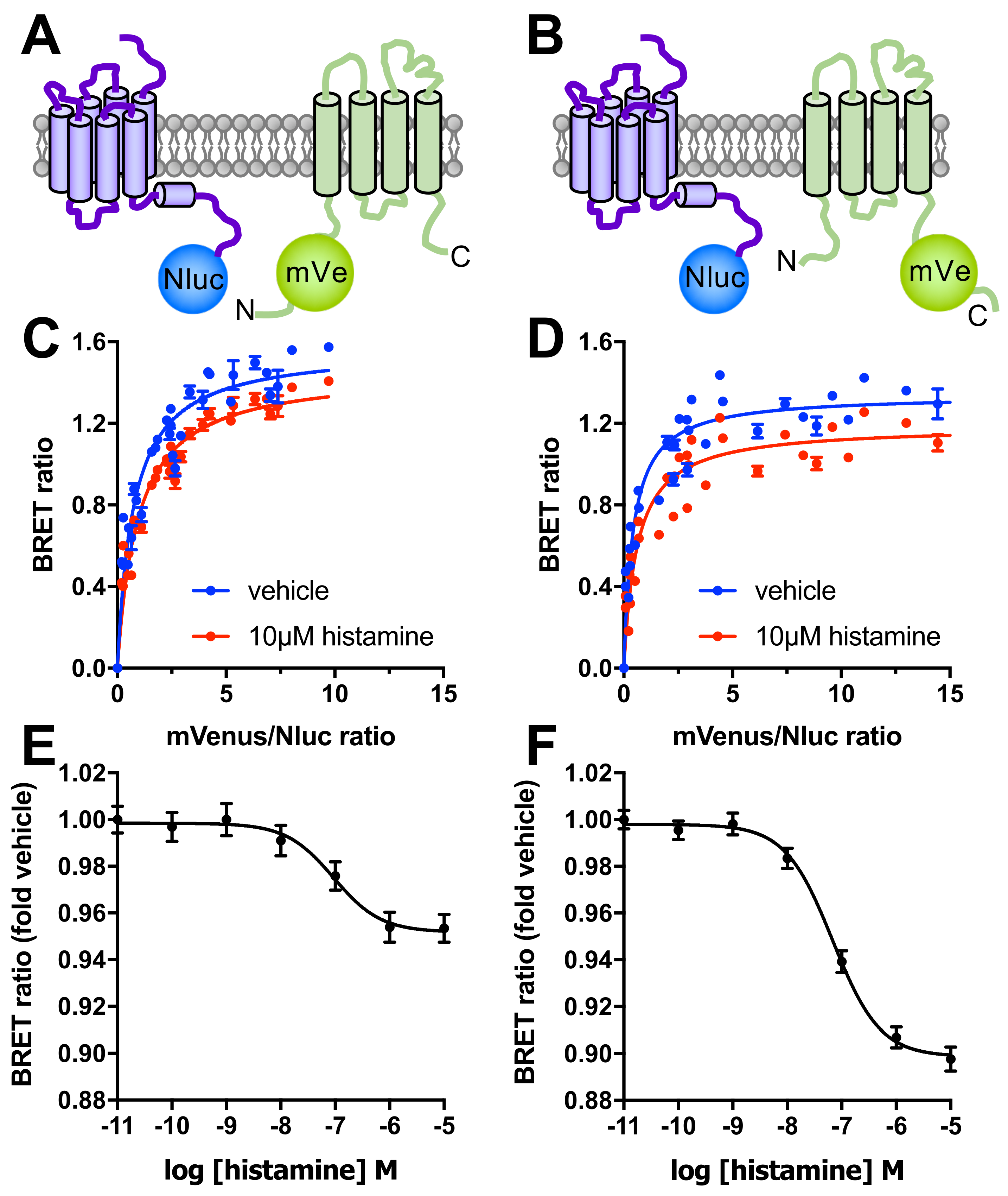
3.3. Saturation BRET Confirms Interaction between H4R and TSPAN4 in HEK293T Cells and Can Be Reduced by Histamine Stimulation
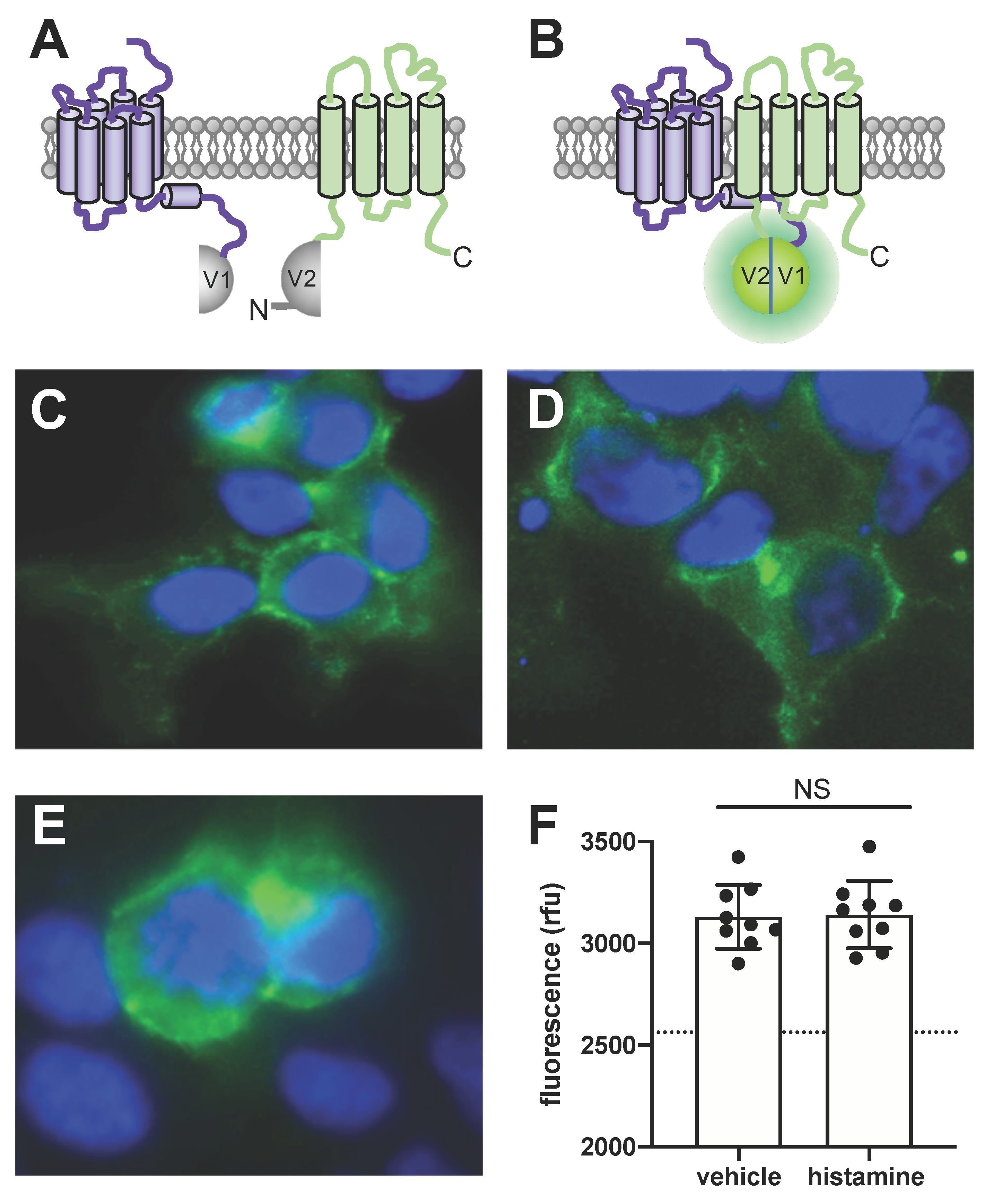
3.4. BiFC Microscopy Reveals H4R-TSPAN4 Complexes in HEK293T Cells
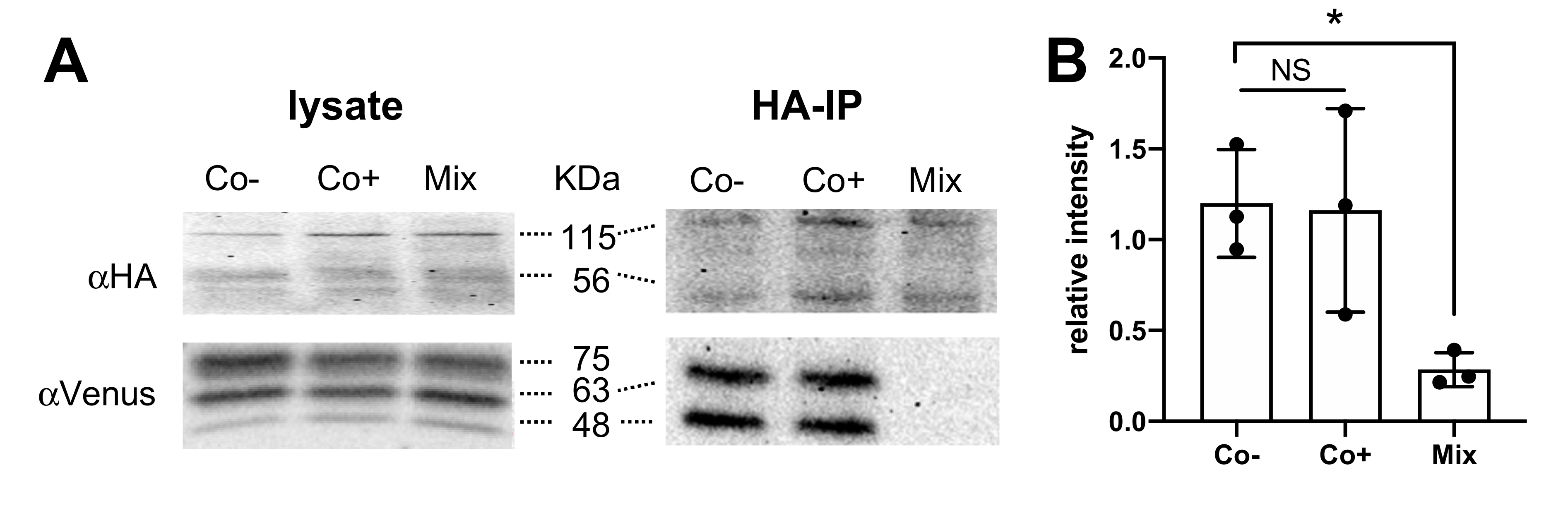
3.5. TSPAN4 Co-Immunoprecipitates with H4R from Co-Expressing HEK293T Cells
3.6. Overexpression of TSPAN4 Does Not Affect Histamine Binding to H4R or Signaling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.S.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [Green Version]
- Thurmond, R.L. The histamine H4 receptor: From orphan to the clinic. Front. Pharmacol. 2015, 6, 65. [Google Scholar] [CrossRef]
- Nakamura, T.; Itadani, H.; Hidaka, Y.; Ohta, M.; Tanaka, K. Molecular cloning and characterization of a new human histamine receptor, HH4R. Biochem. Biophys. Res. Commun. 2000, 279, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Morikawa, N.; Saito, Y.; Masuho, Y.; Matsumoto, S. Molecular cloning and characterization of a novel type of histamine receptor preferentially expressed in leukocytes. J. Biol. Chem. 2000, 275, 36781–36786. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ma, X.-J.; Jiang, X.; Wilson, S.J.; Hofstra, C.L.; Blevitt, J.; Pyati, J.; Li, X.; Chai, W.; Carruthers, N.; et al. Cloning and pharmacological characterization of a fourth histamine receptor (H(4)) expressed in bone marrow. Mol. Pharmacol. 2001, 59, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.D.; van Rijn, R.M.; Ling, P.; Bakker, R.A.; Thurmond, R.L.; Leurs, R. Evaluation of histamine H1-, H2-, and H3-receptor ligands at the human histamine H4 receptor: Identification of 4-methylhistamine as the first potent and selective H4 receptor agonist. J. Pharmacol. Exp. Ther. 2005, 314, 1310–1321. [Google Scholar] [CrossRef] [Green Version]
- Nijmeijer, S.; Vischer, H.F.; Rosethorne, E.M.; Charlton, S.J.; Leurs, R. Analysis of Multiple Histamine H4 Receptor Compound Classes Uncovers Gαi Protein- and β-Arrestin2-Biased Ligands. Mol. Pharmacol. 2012, 82, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Buckland, K.F.; Williams, T.J.; Conroy, D.M. Histamine induces cytoskeletal changes in human eosinophils via the H4 receptor. Br. J. Pharmacol. 2003, 140, 1117–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofstra, C.L.; Desai, P.J.; Thurmond, R.L.; Fung-Leung, W.-P. Histamine H4 receptor mediates chemotaxis and calcium mobilization of mast cells. J. Pharmacol. Exp. Ther. 2003, 305, 1212–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, D.; Leurs, R.; Chazot, P.; Shenton, F.C.; Stark, H.; Werfel, T.; Gutzmer, R. Histamine downregulates monocyte CCL2 production through the histamine H4 receptor. J. Allergy Clin. Immunol. 2007, 120, 300–307. [Google Scholar] [CrossRef]
- Gutzmer, R.; Diestel, C.; Mommert, S.; Köther, B.; Stark, H.; Wittmann, M.; Werfel, T. Histamine H4 receptor stimulation suppresses IL-12p70 production and mediates chemotaxis in human monocyte-derived dendritic cells. J. Immunol. 2005, 174, 5224–5232. [Google Scholar] [CrossRef]
- Desai, P.; Thurmond, R.L. Histamine H4 receptor activation enhances LPS-induced IL-6 production in mast cells via ERK and PI3K activation. Eur. J. Immunol. 2011, 41, 1764–1773. [Google Scholar] [CrossRef]
- Karlstedt, K.; Jin, C.; Panula, P. Expression of histamine receptor genes Hrh3 and Hrh4 in rat brain endothelial cells. Br. J. Pharmacol. 2013, 170, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Verweij, E.W.E.; Al Araaj, B.; Prabhata, W.R.; Prihandoko, R.; Nijmeijer, S.; Tobin, A.B.; Leurs, R.; Vischer, H.F. Differential Role of Serines and Threonines in Intracellular Loop 3 and C-Terminal Tail of the Histamine H4 Receptor in β-Arrestin and G Protein-Coupled Receptor Kinase Interaction, Internalization, and Signaling. ACS Pharmacol. Transl. Sci. 2020, 3, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Rosethorne, E.M.; Charlton, S.J. Agonist-Biased Signaling at the Histamine H4 Receptor: JNJ7777120 Recruits {beta}-Arrestin without Activating G Proteins. Mol. Pharmacol. 2011, 79, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Ritter, S.L.; Hall, R.A. Fine-tuning of GPCR activity by receptor-interacting proteins. Nat. Rev. Mol. Cell Biol. 2009, 10, 819–830. [Google Scholar] [CrossRef] [Green Version]
- Bockaert, J.; Perroy, J.; Bécamel, C.; Marin, P.; Fagni, L. GPCR interacting proteins (GIPs) in the nervous system: Roles in physiology and pathologies. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 89–109. [Google Scholar] [CrossRef]
- Magalhaes, A.C.; Dunn, H.; Ferguson, S.S.G. Regulation of GPCR activity, trafficking and localization by GPCR-interacting proteins. Br. J. Pharmacol. 2012, 165, 1717–1736. [Google Scholar] [CrossRef] [Green Version]
- Heydorn, A.; Søndergaard, B.P.; Ersbøll, B.; Holst, B.; Nielsen, F.C.; Haft, C.R.; Whistler, J.; Schwartz, T.W. A library of 7TM receptor C-terminal tails. Interactions with the proposed post-endocytic sorting proteins ERM-binding phosphoprotein 50 (EBP50), N-ethylmaleimide-sensitive factor (NSF), sorting nexin 1 (SNX1), and G protein-coupled receptor-associated sorting protein (GASP). J. Biol. Chem. 2004, 279, 54291–54303. [Google Scholar] [PubMed] [Green Version]
- Maurice, P.; Daulat, A.M.; Broussard, C.; Mozo, J.; Clary, G.; Hotellier, F.; Chafey, P.; Guillaume, J.-L.; Ferry, G.; Boutin, J.A.; et al. A generic approach for the purification of signaling complexes that specifically interact with the carboxyl-terminal domain of G protein-coupled receptors. Mol. Cell Proteom. 2008, 7, 1556–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, K.Y.; Day, P.W.; Vélez-Ruiz, G.; Sunahara, R.K.; Kobilka, B.K. Identification of GPCR-Interacting Cytosolic Proteins Using HDL Particles and Mass Spectrometry-Based Proteomic Approach. PLoS ONE 2013, 8, e54942. [Google Scholar] [CrossRef] [PubMed]
- Lobingier, B.T.; Hüttenhain, R.; Eichel, K.; Miller, K.B.; Ting, A.Y.; von Zastrow, M.; Krogan, N.J. An Approach to Spatiotemporally Resolve Protein Interaction Networks in Living Cells. Cell 2017, 169, 350–360.e12. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, C.T.; Wang, J.; Paulo, J.A.; Jiang, X.; Gygi, S.P.; Rockman, H.A. Mapping Angiotensin II Type 1 Receptor-Biased Signaling Using Proximity Labeling and Proteomics Identifies Diverse Actions of Biased Agonists. J. Proteome Res. 2021, 20, 3256–3267. [Google Scholar] [CrossRef]
- Paek, J.; Kalocsay, M.; Staus, D.P.; Wingler, L.; Pascolutti, R.; Paulo, J.A.; Gygi, S.P.; Kruse, A.C. Multidimensional Tracking of GPCR Signaling via Peroxidase-Catalyzed Proximity Labeling. Cell 2017, 169, 338–349.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Kittanakom, S.; Wong, V.; Reyes, B.A.; Van Bockstaele, E.J.; Stagljar, I.; Berrettini, W.; Levenson, R. Interaction of the mu-opioid receptor with GPR177 (Wntless) inhibits Wnt secretion: Potential implications for opioid dependence. BMC Neurosci. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petko, J.; Justice-Bitner, S.; Jin, J.; Wong, V.; Kittanakom, S.; Ferraro, T.N.; Stagljar, I.; Levenson, R. MOR is not enough: Identification of novel mu-opioid receptor interacting proteins using traditional and modified membrane yeast two-hybrid screens. PLoS ONE 2013, 8, e67608. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Dai, F.F.; Gaisano, G.; Giglou, K.; Han, J.; Zhang, M.; Kittanakom, S.; Wong, V.; Wei, L.; Showalter, A.D.; et al. The identification of novel proteins that interact with the GLP-1 receptor and restrain its activity. Mol. Endocrinol. 2013, 27, 1550–1563. [Google Scholar] [CrossRef] [Green Version]
- Kittanakom, S.; Barrios-Rodiles, M.; Petschnigg, J.; Arnoldo, A.; Wong, V.; Kotlyar, M.; Heisler, L.E.; Jurisica, I.; Wrana, J.L.; Nislow, C.; et al. CHIP-MYTH: A novel interactive proteomics method for the assessment of agonist-dependent interactions of the human β2-adrenergic receptor. Biochem. Biophys. Res. Commun. 2014, 445, 746–756. [Google Scholar] [CrossRef] [Green Version]
- Sokolina, K.; Kittanakom, S.; Snider, J.; Kotlyar, M.; Maurice, P.; Gandía, J.; Benleulmi-Chaachoua, A.; Tadagaki, K.; Oishi, A.; Wong, V.; et al. Systematic protein-protein interaction mapping for clinically relevant human GPCRs. Mol. Syst. Biol. 2017, 13, 918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, F.F.; Bhattacharjee, A.; Liu, Y.; Batchuluun, B.; Zhang, M.; Wang, X.S.; Huang, X.; Luu, L.; Zhu, D.; Gaisano, H.; et al. A Novel GLP1 Receptor Interacting Protein ATP6ap2 Regulates Insulin Secretion in Pancreatic Beta Cells. J. Biol. Chem. 2015, 290, 25045–25061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Han, J.; Wang, Q.; Mao, Y.; Wei, M.; Jia, W.; Wei, L. A Novel Interacting Protein SERP1 Regulates the N-Linked Glycosylation and Function of GLP-1 Receptor in the Liver. J. Cell Biochem. 2017, 118, 3616–3626. [Google Scholar] [CrossRef]
- Snider, J.; Kittanakom, S.; Damjanovic, D.; Curak, J.; Wong, V.; Stagljar, I. Detecting interactions with membrane proteins using a membrane two-hybrid assay in yeast. Nat. Protoc. 2010, 5, 1281–1293. [Google Scholar] [CrossRef]
- UniProt Consortium UniProt: The universal protein knowledgebase in 2021. Nucleic. Acids Res. 2021, 49, D480–D489. [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Klebe, R.J.; Harriss, J.V.; Sharp, Z.D.; Douglas, M.G. A general method for polyethylene-glycol-induced genetic transformation of bacteria and yeast. Gene 1983, 25, 333–341. [Google Scholar] [CrossRef]
- Vischer, H.F.; Nijmeijer, S.; Smit, M.J.; Leurs, R. Viral hijacking of human receptors through heterodimerization. Biochem. Biophys. Res. Commun. 2008, 377, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Leurs, R.; Vischer, H.F. NanoLuc-Based Methods to Measure β-Arrestin2 Recruitment to G Protein-Coupled Receptors. Methods Mol. Biol. 2021, 2268, 233–248. [Google Scholar] [PubMed]
- Nijmeijer, S.; Leurs, R.; Smit, M.J.; Vischer, H.F. The Epstein-Barr virus-encoded G protein-coupled receptor BILF1 hetero-oligomerizes with human CXCR4, scavenges Gαi proteins, and constitutively impairs CXCR4 functioning. J. Biol. Chem. 2010, 285, 29632–29641. [Google Scholar] [CrossRef] [Green Version]
- van Rijn, R.M.; Chazot, P.L.; Shenton, F.C.; Sansuk, K.; Bakker, R.A.; Leurs, R. Oligomerization of recombinant and endogenously expressed human histamine H(4) receptors. Mol. Pharmacol. 2006, 70, 604–615. [Google Scholar] [CrossRef]
- Schihada, H.; Shekhani, R.; Schulte, G. Quantitative assessment of constitutive G protein-coupled receptor activity with BRET-based G protein biosensors. bioRxiv 2021. [Google Scholar] [CrossRef]
- Mercier, J.-F.; Salahpour, A.; Angers, S.; Breit, A.; Bouvier, M. Quantitative assessment of beta 1- and beta 2-adrenergic receptor homo- and heterodimerization by bioluminescence resonance energy transfer. J. Biol. Chem. 2002, 277, 44925–44931. [Google Scholar] [CrossRef] [Green Version]
- Termini, C.M.; Gillette, J.M. Tetraspanins Function as Regulators of Cellular Signaling. Front. Cell Dev. Biol. 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Robert, J.-M.H.; Amoussou, N.G.; Mai, H.L.; Logé, C.; Brouard, S. Tetraspanins: Useful multifunction proteins for the possible design and development of small-molecule therapeutic tools. Drug. Discov. Today 2020, 26, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Urizar, E.; Kralikova, M.; Mobarec, J.C.; Shi, L.; Filizola, M.; Javitch, J.A. Dopamine D2 receptors form higher order oligomers at physiological expression levels. EMBO J. 2008, 27, 2293–2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyu, Y.J.; Hu, C.-D. Fluorescence complementation: An emerging tool for biological research. Trends Biotechnol. 2008, 26, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Kerppola, T.K. Visualization of molecular interactions by fluorescence complementation. Nat. Rev. Mol. Cell Biol. 2006, 7, 449–456. [Google Scholar] [CrossRef]
- Kawano, Y.; Yoshida, T.; Hieda, K.; Aoki, J.; Miyoshi, H.; Koyanagi, Y. A lentiviral cDNA library employing lambda recombination used to clone an inhibitor of human immunodeficiency virus type 1-induced cell death. J. Virol. 2004, 78, 11352–11359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Kawano, Y.; Sato, K.; Ando, Y.; Aoki, J.; Miura, Y.; Komano, J.; Tanaka, Y.; Koyanagi, Y. A CD63 mutant inhibits T-cell tropic human immunodeficiency virus type 1 entry by disrupting CXCR4 trafficking to the plasma membrane. Traffic 2008, 9, 540–558. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Kitayama, D.; Arima, M.; Sakamoto, A.; Inamine, A.; Watanabe-Takano, H.; Hatano, M.; Koike, T.; Tokuhisa, T. CXCR4 expression on activated B cells is downregulated by CD63 and IL-21. J. Immunol. 2011, 186, 2800–2808. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Ebina, H.; Koyanagi, Y. N-linked glycan-dependent interaction of CD63 with CXCR4 at the Golgi apparatus induces downregulation of CXCR4. Microbiol. Immunol. 2009, 53, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zucker, B.; Zhang, S.; Elias, S.; Zhu, Y.; Chen, H.; Ding, T.; Li, Y.; Sun, Y.; Lou, J.; et al. Migrasome formation is mediated by assembly of micron-scale tetraspanin macrodomains. Nat. Cell Biol. 2019, 21, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Jiang, Z.; Lu, D.; Wang, X.; Liang, H.; Zhang, J.; Meng, Y.; Li, Y.; Wu, D.; Huang, Y.; et al. Migrasomes provide regional cues for organ morphogenesis during zebrafish gastrulation. Nature 2019, 21, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Ling, P.; Ngo, K.; Nguyen, S.; Thurmond, R.L.; Edwards, J.P.; Karlsson, L.; Fung-Leung, W.-P. Histamine H4 receptor mediates eosinophil chemotaxis with cell shape change and adhesion molecule upregulation. Br. J. Pharmacol. 2004, 142, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, D.; Stark, H.; Chazot, P.L.; Shenton, F.C.; Leurs, R.; Werfel, T.; Gutzmer, R. Human inflammatory dendritic epidermal cells express a functional histamine H4 receptor. J. Investig. Dermatol. 2008, 128, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäumer, W.; Wendorff, S.; Gutzmer, R.; Werfel, T.; Dijkstra, D.; Chazot, P.; Stark, H.; Kietzmann, M. Histamine H4 receptors modulate dendritic cell migration through skin--immunomodulatory role of histamine. Allergy 2008, 63, 1387–1394. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Rossbach, K.; Dijkstra, D.; Bäumer, W.; Kietzmann, M.; Stark, H.; Werfel, T.; Gutzmer, R. Murine and human Langerhans cells express a functional histamine H4 receptor: Modulation of cell migration and function. Allergy 2010, 65, 840–849. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Mommert, S.; Köther, B.; Werfel, T.; Gutzmer, R. The histamine H4 receptor is highly expressed on plasmacytoid dendritic cells in psoriasis and histamine regulates their cytokine production and migration. J. Investig. Dermatol. 2011, 131, 1668–1676. [Google Scholar] [CrossRef] [Green Version]
- Kay, L.J.; Suvarna, S.K.; Peachell, P.T. Histamine H4 receptor mediates chemotaxis of human lung mast cells. Eur. J. Pharmacol. 2018, 837, 38–44. [Google Scholar] [CrossRef]
- Kuramasu, A.; Wakabayashi, M.; Inui, M.; Yanai, K. Distinct roles of small GTPases RAC1 and RAC2 in histamine H4 receptor–mediated chemotaxis of mast cells. J. Pharmacol. Exp. Ther. 2018, 367, 9–19. [Google Scholar] [CrossRef]
- Moya-Garcia, A.A.; Rodriguez, C.E.; Morilla, I.; Sanchez-Jiménez, F.; Ranea, J.A.G. The function of histamine receptor H4R in the brain revealed by interaction partners. Front. Biosci. 2011, 3, 1058–1066. [Google Scholar] [CrossRef]
- White, C.W.; Caspar, B.; Vanyai, H.K.; Pfleger, K.D.G.; Hill, S.J. CRISPR-Mediated Protein Tagging with Nanoluciferase to Investigate Native Chemokine Receptor Function and Conformational Changes. Cell Chem. Biol. 2020, 27, 499–510.e7. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (co-)Expressed Constructs | pKd | Bmax (fmol/mg) |
|---|---|---|
| H4R | 7.9 ± 0.1 | 430 ± 74 |
| H4R + mVenus-TSPAN4 | 8.0 ± 0.3 | 86 ± 40 |
| H4R + TSPAN4-mVenus | 8.0 ± 0.4 | 75 ± 43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Verweij, E.W.E.; Siderius, M.; Leurs, R.; Vischer, H.F. Identification of TSPAN4 as Novel Histamine H4 Receptor Interactor. Biomolecules 2021, 11, 1127. https://doi.org/10.3390/biom11081127
Ma X, Verweij EWE, Siderius M, Leurs R, Vischer HF. Identification of TSPAN4 as Novel Histamine H4 Receptor Interactor. Biomolecules. 2021; 11(8):1127. https://doi.org/10.3390/biom11081127
Chicago/Turabian StyleMa, Xiaoyuan, Eléonore W. E. Verweij, Marco Siderius, Rob Leurs, and Henry F. Vischer. 2021. "Identification of TSPAN4 as Novel Histamine H4 Receptor Interactor" Biomolecules 11, no. 8: 1127. https://doi.org/10.3390/biom11081127
APA StyleMa, X., Verweij, E. W. E., Siderius, M., Leurs, R., & Vischer, H. F. (2021). Identification of TSPAN4 as Novel Histamine H4 Receptor Interactor. Biomolecules, 11(8), 1127. https://doi.org/10.3390/biom11081127