
Opposing Actions of Octopamine and Tyramine on Honeybee Vision



Abstract
:1. Introduction
2. Materials and Methods
2.1. Honeybees
2.2. Electroretinography
2.3. Phototaxis Assay
2.4. Data Analysis
3. Results
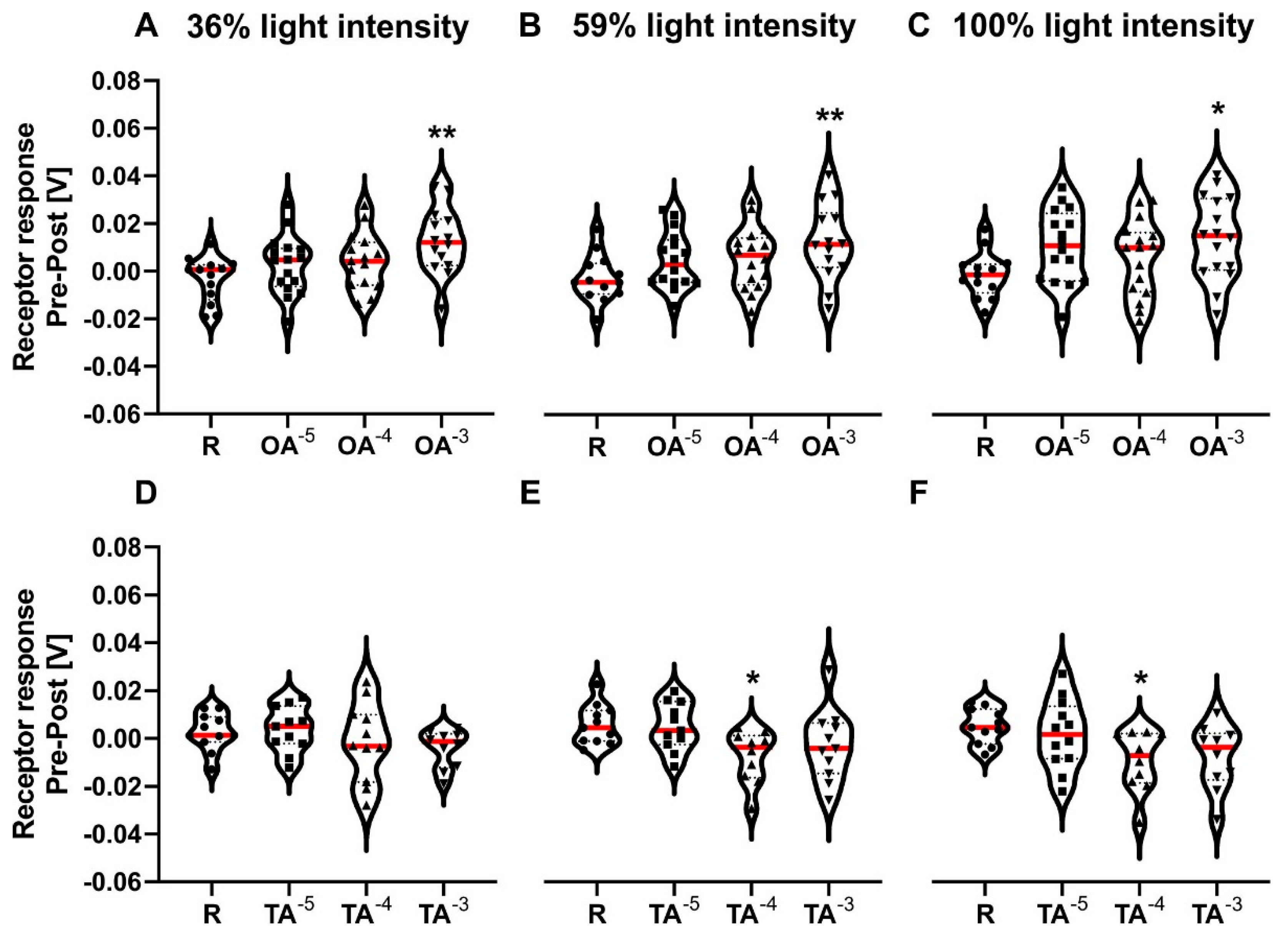
3.1. The Effect of Octopamine and Tyramine on the ERG Response
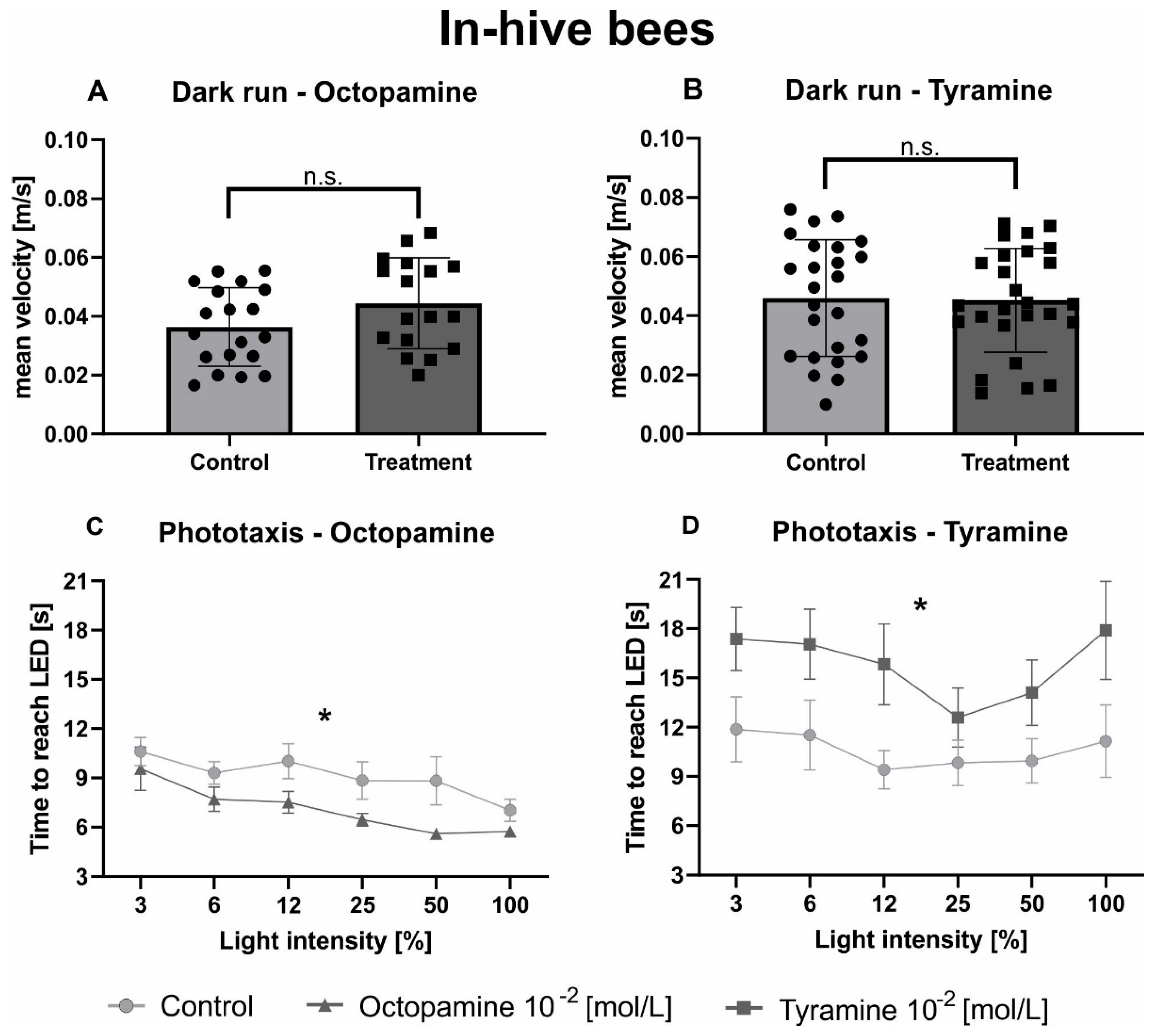
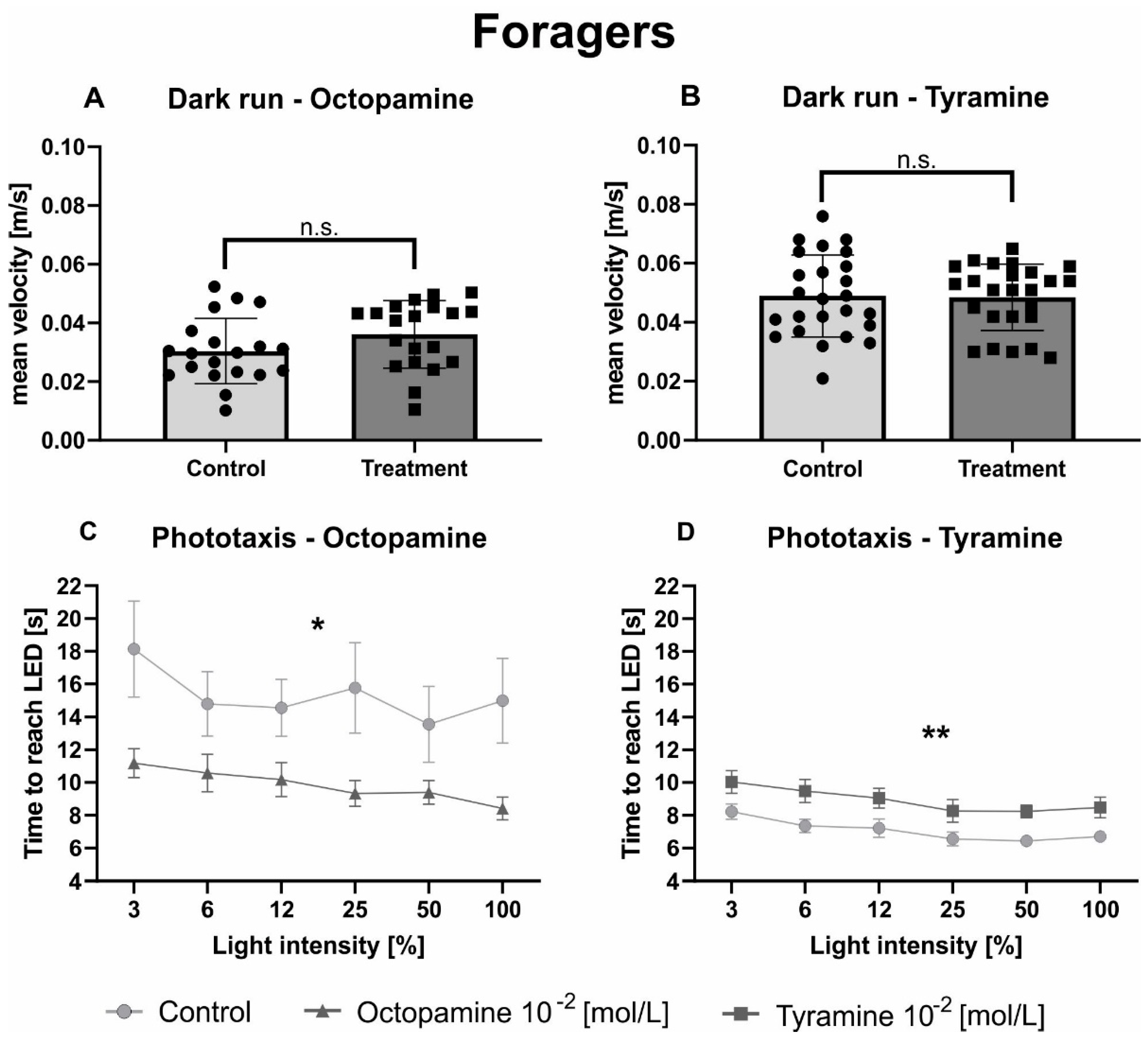
3.2. Tyramine and Octopamine Have Opposite Effects on the Phototaxis of In-Hive Bees and Foragers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blenau, W.; Baumann, A. Molecular and pharmacological properties of insect biogenic amine receptors: Lessons from Drosophila melanogaster and Apis mellifera. Arch. Insect Biochem. Physiol. 2001, 48, 13–38. [Google Scholar] [CrossRef]
- Blenau, W.; Baumann, A. Aminergic signal transduction in invertebrates: Focus on tyramine and octopamine receptors. Recent Res. Dev. Neurochem. 2003, 6, 225–240. [Google Scholar]
- Evans, P.D. Biogenic Amines in the Insect Nervous System. Adv. Insect Phys. 1980, 15, 317–473. [Google Scholar] [CrossRef]
- Roeder, T.; Seifert, M.; Kähler, C.; Gewecke, M. Tyramine and octopamine: Antagonistic modulators of behavior and metabolism. Arch. Insect Biochem. Physiol. 2003, 54, 1–13. [Google Scholar] [CrossRef]
- Roeder, T. Tyramine and octopamine: Ruling behavior and metabolism. Annu. Rev. Entomol. 2005, 50, 447–477. [Google Scholar] [CrossRef]
- Roeder, T. The control of metabolic traits by octopamine and tyramine in invertebrates. J. Exp. Biol. 2020, 223, jeb194282. [Google Scholar] [CrossRef]
- Scheiner, R.; Baumann, A.; Blenau, W. Aminergic Control and Modulation of Honeybee Behaviour. Curr. Neuropharmacol. 2006, 4, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.W.; Woodring, J. Effects of stress, age, season, and source colony on levels of octopamine, dopamine and serotonin in the honey bee (Apis mellifera L.) brain. J. Insect Physiol. 1992, 38, 29–35. [Google Scholar] [CrossRef]
- Wagener-Hulme, C.; Kuehn, J.C.; Schulz, D.J.; Robinson, G.E. Biogenic amines and division of labor in honey bee colonies. J. Comp. Physiol. A 1999, 184, 471–479. [Google Scholar] [CrossRef]
- Scheiner, R.; Reim, T.; Søvik, E.; Entler, B.V.; Barron, A.B.; Thamm, M. Learning, gustatory responsiveness and tyramine differences across nurse and forager honeybees. J. Exp. Biol. 2017, 220, 1443–1450. [Google Scholar] [CrossRef] [Green Version]
- Grohmann, L.; Blenau, W.; Erber, J.; Ebert, P.R.; Strünker, T.; Baumann, A. Molecular and functional characterization of an octopamine receptor from honeybee (Apis mellifera) brain. J. Neurochem. 2003, 86, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Blenau, W.; Balfanz, S.; Baumann, A. Amtyr1: Characterization of a Gene from Honeybee (Apis mellifera) Brain Encoding a Functional Tyramine Receptor. J. Neurochem. 2000, 74, 900–908. [Google Scholar] [CrossRef]
- Blenau, W.; Wilms, J.A.; Balfanz, S.; Baumann, A. AmOctα2R: Functional characterization of a honeybee octopamine receptor inhibiting adenylyl cyclase activity. Int. J. Mol. Sci. 2020, 21, 9334. [Google Scholar] [CrossRef]
- Balfanz, S.; Jordan, N.; Langenstück, T.; Breuer, J.; Bergmeier, V.; Baumann, A. Molecular, pharmacological, and signaling properties of octopamine receptors from honeybee (Apis mellifera) brain. J. Neurochem. 2014, 129, 284–296. [Google Scholar] [CrossRef]
- Reim, T.; Balfanz, S.; Baumann, A.; Blenau, W.; Thamm, M.; Scheiner, R. AmTAR2: Functional characterization of a honeybee tyramine receptor stimulating adenylyl cyclase activity. Insect Biochem. Mol. Biol. 2017, 80, 91–100. [Google Scholar] [CrossRef]
- Behrends, A.; Scheiner, R. Octopamine improves learning in newly emerged bees but not in old foragers. J. Exp. Biol. 2012, 215, 1076–1083. [Google Scholar] [CrossRef] [Green Version]
- Bauknecht, P.; Jékely, G. Ancient coexistence of norepinephrine, tyramine, and octopamine signaling in bilaterians. BMC Biol. 2017, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Saraswati, S.; Fox, L.E.; Soll, D.R.; Wu, C.-F. Tyramine and octopamine have opposite effects on the locomotion of Drosophila larvae. J. Neurobiol. 2004, 58, 425–441. [Google Scholar] [CrossRef]
- Uzzan, A.; Dudai, Y. Aminergic Receptors in Drosophila melanogaster: Responsiveness of Adenylate Cyclase to Putative Neurotransmitters. J. Neurochem. 1982, 38, 1542–1550. [Google Scholar] [CrossRef]
- Scheiner, R.; Toteva, A.; Reim, T.; Søvik, E.; Barron, A.B. Differences in the phototaxis of pollen and nectar foraging honey bees are related to their octopamine brain titers. Front. Physiol. 2014, 5, 116. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Steinbach, A.; Claßen, G.; Strudthoff, N.; Scholz, H. Octopamine indirectly affects proboscis extension response habituation in Drosophila melanogaster by controlling sucrose responsiveness. J. Insect Physiol. 2014, 69, 107–117. [Google Scholar] [CrossRef]
- Menzel, R.; Hammer, M.; Müller, U.; Rosenboom, H. Behavioral, neural and cellular components underlying olfactory learning in the honeybee. Proc. J. Physiol. Paris 1996, 90, 395–398. [Google Scholar] [CrossRef]
- Hammer, M.; Menzel, R. Learning and memory in the honeybee. J. Neurosci. 1995, 15, 1617–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.C.; Lee, H.G.; Lim, J.; Han, K.A. Appetitive learning requires the alpha1-like octopamine receptor oamb in the drosophila mushroom body neurons. J. Neurosci. 2013, 33, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Kutsukake, M.; Komatsu, A.; Yamamoto, D.; Ishiwa-Chigusa, S. A tyramine receptor gene mutation causes a defective olfactory behavior in Drosophila melanogaster. Gene 2000, 245, 31–42. [Google Scholar] [CrossRef]
- McClung, C.; Hirsh, J. The trace amine tyramine is essential for sensitization to cocaine in Drosophila. Curr. Biol. 1999, 9, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Braun, G.; Bicker, G. Habituation of an appetitive reflex in the honeybee. J. Neurophysiol. 1992, 67, 588–598. [Google Scholar] [CrossRef]
- Scheiner, R.; Plückhahn, S.; Öney, B.; Blenau, W.; Erber, J. Behavioural pharmacology of octopamine, tyramine and dopamine in honey bees. Behav. Brain Res. 2002, 136, 545–553. [Google Scholar] [CrossRef]
- Scheiner, R.; Entler, B.V.; Barron, A.B.; Scholl, C.; Thamm, M. The Effects of Fat Body Tyramine Level on Gustatory Responsiveness of Honeybees (Apis mellifera) Differ between Behavioral Castes. Front. Syst. Neurosci. 2017, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Kuritz-Kaiser, A.; Menzel, R.; Erber, J. Sensory responsiveness and the effects of equal subjective rewards on tactile learning and memory of honeybees. Learn. Mem. 2005, 12, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Barron, A.B.; Robinson, G.E. Selective modulation of task performance by octopamine in honey bee (Apis mellifera) division of labour. J. Comp. Physiol. A Neuroethol. 2005, 191, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Schulz, D.J.; Robinson, G.E. Octopamine influences division of labor in honey bee colonies. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 2001, 187, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Reim, T.; Scheiner, R. Division of labour in honey bees: Age- and task-related changes in the expression of octopamine receptor genes. Insect Mol. Biol. 2014, 23, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.J.; Amdam, G.V.; Schlipalius, D.; Emore, C.; Sardesai, N.; Williams, C.E.; Rueppell, O.; Guzmán-Novoa, E.; Arechavaleta-Velasco, M.; Chandra, S.; et al. Behavioral genomics of honeybee foraging and nest defense. Naturwissenschaften 2007, 94, 247–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloppenburg, P.; Erber, J. The modulatory effects of serotonin and octopamine in the visual system of the honey bee (Apis mellifera L.). J. Comp. Physiol. A 1995, 176, 119–129. [Google Scholar] [CrossRef]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis mellifera) and involvement of serotonin in phototactic behavior. Cell. Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef]
- Erber, J.; Hoormann, J.; Scheiner, R. Phototactic behaviour correlates with gustatory responsiveness in honey bees (Apis mellifera L.). Behav. Brain Res. 2006, 174, 174–180. [Google Scholar] [CrossRef]
- Barron, A.B.; Maleszka, R.; Vander Meer, R.K.; Robinson, G.E. Octopamine modulates honey bee dance behavior. Proc. Natl. Acad. Sci. USA 2007, 104, 1703–1707. [Google Scholar] [CrossRef] [Green Version]
- Dudai, Y.; Buxbaum, J.; Corfas, G.; Ofarim, M. Formamidines interact with Drosophila octopamine receptors, alter the flies’ behavior and reduce their learning ability. J. Comp. Physiol. A 1987, 161, 739–746. [Google Scholar] [CrossRef]
- Sinakevitch, I.; Mustard, J.A.; Smith, B.H. Distribution of the octopamine receptor AmOA1 in the honey bee brain. PLoS ONE 2011, 6, e14536. [Google Scholar] [CrossRef]
- Thamm, M.; Scholl, C.; Reim, T.; Grübel, K.; Möller, K.; Rössler, W.; Scheiner, R. Neuronal distribution of tyramine and the tyramine receptor AmTAR1 in the honeybee brain. J. Comp. Neurol. 2017, 525, 2615–2631. [Google Scholar] [CrossRef]
- El-Kholy, S.; Stephano, F.; Li, Y.; Bhandari, A.; Fink, C.; Roeder, T. Expression analysis of octopamine and tyramine receptors in Drosophila. Cell Tissue Res. 2015, 361, 669–684. [Google Scholar] [CrossRef] [PubMed]
- Balfanz, S.; Strünker, T.; Frings, S.; Baumann, A. A family of octopamine [corrected] receptors that specifically induce cyclic AMP production or Ca2+ release in Drosophila melanogaster. J. Neurochem. 2005, 93, 440–451. [Google Scholar] [CrossRef]
- Bayliss, A.; Roselli, G.; Evans, P.D. A comparison of the signalling properties of two tyramine receptors from Drosophila. J. Neurochem. 2013, 125, 37–48. [Google Scholar] [CrossRef]
- Değirmenci, L.; Geiger, D.; Rogé Ferreira, F.L.; Keller, A.; Krischke, B.; Beye, M.; Steffan-Dewenter, I.; Scheiner, R. CRISPR/Cas9 mediated mutations as a new tool for studying taste in honeybees. bioRxiv 2020. [Google Scholar] [CrossRef]
- Scheiner, R.; Kulikovskaja, L.; Thamm, M. The honey bee tyramine receptor AmTYR1 and division of foraging labour. J. Exp. Biol. 2014, 217, 1215–1217. [Google Scholar] [CrossRef] [Green Version]
- Degen, J.; Gewecke, M.; Roeder, T. Octopamine receptors in the honey bee and locust nervous system: Pharmacological similarities between homologous receptors of distantly related species. Br. J. Pharmacol. 2000, 130, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barron, A.B.; Maleszka, J.; Vander Meer, R.K.; Robinson, G.E.; Maleszka, R. Comparing injection, feeding and topical application methods for treatment of honeybees with octopamine. J. Insect Physiol. 2007, 53, 187–194. [Google Scholar] [CrossRef]
- Wanka, L.; Behr, V.; Beck-Sickinger, A.G. Arrestin-dependent internalization of rhodopsin-like G protein-coupled receptors. Biol. Chem. 2021. [Google Scholar] [CrossRef]
- Farooqui, T.; Robinson, K.; Vaessin, H.; Smith, B.H. Modulation of early olfactory processing by an octopaminergic reinforcement pathway in the honeybee. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 5370–5380. [Google Scholar] [CrossRef]
- Scholl, C.; Kübert, N.; Muenz, T.S.; Rössler, W. CaMKII knockdown affects both early and late phases of olfactory long-term memory in the honeybee. J. Exp. Biol. 2015, 218, 3788–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.C.M.; Wasserman, G.S. Categorical and Prolonged Potentials Are Evoked when Brief, Intermediate-Intensity Flashes Stimulate Horseshoe Crab Lateral Eye Photoreceptors during Octopamine Neuromodulation. Neurosignals 2001, 10, 399–415. [Google Scholar] [CrossRef]
- Battelle, B.A.; Edwards, S.C.; Kass, L.; Maresch, H.M.; Pierce, S.K.; Wishart, A.C. Identification and Function of Octopamine and Tyramine Conjugates in the Limulus Visual System. J. Neurochem. 1988, 51, 1240–1251. [Google Scholar] [CrossRef] [PubMed]
- Erber, J.; Kloppenburg, P.; Scheidler, A. Neuromodulation by serotonin and octopamine in the honeybee: Behaviour, neuroanatomy and electrophysiology. Experientia 1993, 49, 1073–1083. [Google Scholar] [CrossRef]
- Chyb, S.; Hevers, W.; Forte, M.; Wolfgang, W.J.; Selinger, Z.; Hardie, R.C. Modulation of the light response by cAMP in Drosophila photoreceptors. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 8799–8807. [Google Scholar] [CrossRef] [Green Version]
- Fussnecker, B.L.; Smith, B.H.; Mustard, J.A. Octopamine and tyramine influence the behavioral profile of locomotor activity in the honey bee (Apis mellifera). J. Insect Physiol. 2006, 52, 1083–1092. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| OA—Pre-Response | Friedmann Test | χ2(2) | p Value | ||
|---|---|---|---|---|---|
| Ringer—Control | 14.63 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 16 | 16 | 0.024 | * | |
| 36% vs. 100% | 16 | 16 | <0.001 | *** | |
| 59% vs. 100% | 16 | 16 | 0.867 | n.s. | |
| OA—post response | Friedmann test | χ2(2) | p value | ||
| Ringer—Control | 16.63 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 16 | 16 | 0.008 | ** | |
| 36% vs. 100% | 16 | 16 | <0.001 | *** | |
| 59% vs. 100% | 16 | 16 | 0.99 | n.s. | |
| OA—pre-response | Friedmann test | χ2(2) | p value | ||
| OA—Treatment | 26.79 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 48 | 48 | 0.001 | ** | |
| 36% vs. 100% | 48 | 48 | <0.001 | *** | |
| 59% vs. 100% | 48 | 48 | 0.99 | n.s. | |
| OA—post response | Friedmann test | χ2(2) | p value | ||
| OA—Treatment | 38.17 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 48 | 48 | <0.001 | *** | |
| 36% vs. 100% | 48 | 48 | <0.001 | *** | |
| 59% vs. 100% | 48 | 48 | 0.459 | n.s. | |
| TA—pre-response | Friedmann test | χ2(2) | p value | ||
| Ringer—Control | 24 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 12 | 12 | 0.043 | * | |
| 36% vs. 100% | 12 | 12 | <0.001 | *** | |
| 59% vs. 100% | 12 | 12 | 0.043 | * | |
| TA—post response | Friedmann test | χ2(2) | p value | ||
| Ringer—Control | 24 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 12 | 12 | 0.043 | * | |
| 36% vs. 100% | 12 | 12 | <0.001 | *** | |
| 59% vs. 100% | 12 | 12 | 0.043 | * | |
| TA—pre-response | Friedmann test | χ2(2) | p value | ||
| TA—Treatment | 70.06 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 36 | 36 | <0.001 | *** | |
| 36% vs. 100% | 36 | 36 | <0.001 | *** | |
| 59% vs. 100% | 36 | 36 | <0.001 | *** | |
| TA—post response | Friedmann test | χ2(2) | p value | ||
| TA—Treatment | 62.39 | <0.001 | *** | ||
| Dunn´s test | n (Ringer) | n (Treatment) | |||
| 36% vs. 59% | 36 | 36 | <0.001 | *** | |
| 36% vs. 100% | 36 | 36 | <0.001 | *** | |
| 59% vs. 100% | 36 | 36 | <0.001 | *** |
| OA—Pre-Post Response | 1way ANOVA | F (3, 54) | p Value | ||||
|---|---|---|---|---|---|---|---|
| - 36% light intensity | 4091 | 0.011 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. OA 10-3 | 13 | 14 | 3457 | 54 | 0.003 | ** | |
| R vs. OA 10-4 | 13 | 15 | 1667 | 54 | 0.237 | n.s. | |
| R vs. OA 10-5 | 13 | 16 | 1.37 | 54 | 0.386 | n.s. | |
| OA—pre-post response | 1way ANOVA | F (3, 55) | p value | ||||
| - 59% light intensity | 3176 | 0.031 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. OA 10-3 | 13 | 14 | 3078 | 55 | 0.009 | ** | |
| R vs. OA 10-4 | 13 | 16 | 1768 | 55 | 0.197 | n.s. | |
| R vs. OA 10-5 | 13 | 16 | 1544 | 55 | 0.293 | n.s. | |
| OA—pre-post response | 1way ANOVA | F (3, 57) | p value | ||||
| - 100% light intensity | 2821 | 0.0469 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. OA 10-3 | 13 | 16 | 2744 | 57 | 0.022 | * | |
| R vs. OA 10-4 | 13 | 16 | 1232 | 57 | 0.471 | n.s. | |
| R vs. OA 10-5 | 13 | 16 | 2118 | 57 | 0.098 | n.s. | |
| TA—pre-post response | 1way ANOVA | F (3, 39) | p value | ||||
| - 36% light intensity | 1.16 | 0.337 | n.s. | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | statistics | DF | |||
| R vs. TA 10-3 | 11 | 10 | 1404 | 39 | 0.37 | n.s. | |
| R vs. TA 10-4 | 11 | 11 | 0.892 | 39 | 0.703 | n.s. | |
| R vs. TA 10-5 | 11 | 11 | 0.232 | 39 | 0.991 | n.s. | |
| TA—pre-post response | 1way ANOVA | F (3, 39) | p value | ||||
| - 59% light intensity | 3304 | 0.03 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. TA 10-3 | 11 | 11 | 2 | 39 | 0.211 | n.s. | |
| R vs. TA 10-4 | 11 | 10 | 2677 | 39 | 0.029 | * | |
| R vs. TA 10-5 | 11 | 11 | 0.162 | 39 | 0.997 | n.s. | |
| TA—pre-post response | 1way ANOVA | F (3, 39) | p value | ||||
| - 100% light intensity | 3492 | 0.025 | * | ||||
| Dunnett´s test | n (Ringer) | n (Treatment) | q | DF | |||
| R vs. TA 10-3 | 11 | 10 | 2373 | 39 | 0.059 | n.s. | |
| R vs. TA 10-4 | 11 | 10 | 2702 | 39 | 0.027 | * | |
| R vs. TA 10-5 | 11 | 12 | 0.6678 | 39 | 0.844 | n.s. |
| Octopamine (In-Hive) | Unpaired t Test | Statistic | DF | n (Ringer) | n (Treatment) | p Value | |
|---|---|---|---|---|---|---|---|
| Dark run | t = 1.679 | 34 | 19 | 17 | 0.102 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 170) = 6.131 | <0.001 | *** | ||||
| Treatment | F(1, 34) = 5.750 | 0.022 | * | ||||
| Interaction | F(5, 170) = 0.625 | 0.681 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 0.824 | 204 | 19 | 17 | 0.999 | n.s. | |
| 6% | 1258 | 204 | 19 | 17 | 0.999 | n.s. | |
| 12% | 1960 | 204 | 19 | 17 | 0.308 | n.s. | |
| 25% | 1874 | 204 | 19 | 17 | 0.374 | n.s. | |
| 50% | 2531 | 204 | 19 | 17 | 0.073 | n.s. | |
| 100% | 1017 | 204 | 19 | 17 | 0.999 | n.s. | |
| Tyramine (in-hive) | unpaired t test | statistic | DF | n (Ringer) | n (Treatment) | p value | |
| Dark run | t = 0.139 | 49 | 25 | 26 | 0.890 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 245) = 2.564 | 0.028 | * | ||||
| Treatment | F(1, 49) = 4.919 | 0.031 | * | ||||
| Interaction | F(5, 245) = 0.669 | 0.647 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 1916 | 294 | 25 | 26 | 0.338 | n.s. | |
| 6% | 1931 | 294 | 25 | 26 | 0.327 | n.s. | |
| 12% | 2236 | 294 | 25 | 26 | 0.157 | n.s. | |
| 25% | 0.964 | 294 | 25 | 26 | 0.999 | n.s. | |
| 50% | 1452 | 294 | 25 | 26 | 0.886 | n.s. | |
| 100% | 2352 | 294 | 25 | 26 | 0.116 | n.s. | |
| Octopamine (forager) | unpaired t test | statistic | DF | n (Ringer) | n (Treatment) | p value | |
| Dark run | t = 1.595 | 38 | 20 | 20 | 0.119 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 190) = 4.342 | < 0.001 | *** | ||||
| Treatment | F(1, 38) = 5.223 | 0.028 | * | ||||
| Interaction | F(5, 190) = 1.514 | 0.187 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 2695 | 228 | 20 | 20 | 0.045 | * | |
| 6% | 1630 | 228 | 20 | 20 | 0.627 | n.s. | |
| 12% | 1700 | 228 | 20 | 20 | 0.543 | n.s. | |
| 25% | 2492 | 228 | 20 | 20 | 0.081 | n.s. | |
| 50% | 1610 | 228 | 20 | 20 | 0.653 | n.s. | |
| 100% | 2545 | 228 | 20 | 20 | 0.070 | n.s. | |
| Tyramine (forager) | unpaired t test | statistic | DF | n (Ringer) | n (Treatment) | p value | |
| Dark run | t = 0.123 | 48 | 25 | 25 | 0.903 | n.s. | |
| Phototaxis | 2way ANOVA | ||||||
| Intensity | F(5, 245) = 5.986 | < 0.001 | *** | ||||
| Treatment | F(1, 49) = 11.29 | 0.002 | ** | ||||
| Interaction | F(5, 245) = 0.072 | 0.996 | n.s. | ||||
| Bonferroni test | |||||||
| 3% | 2412 | 294 | 26 | 25 | 0.099 | n.s. | |
| 6% | 2838 | 294 | 26 | 25 | 0.029 | * | |
| 12% | 2435 | 294 | 26 | 25 | 0.093 | n.s. | |
| 25% | 2268 | 294 | 26 | 25 | 0.144 | n.s. | |
| 50% | 2377 | 294 | 26 | 25 | 0.109 | n.s. | |
| 100% | 2334 | 294 | 26 | 25 | 0.122 | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schilcher, F.; Thamm, M.; Strube-Bloss, M.; Scheiner, R. Opposing Actions of Octopamine and Tyramine on Honeybee Vision. Biomolecules 2021, 11, 1374. https://doi.org/10.3390/biom11091374
Schilcher F, Thamm M, Strube-Bloss M, Scheiner R. Opposing Actions of Octopamine and Tyramine on Honeybee Vision. Biomolecules. 2021; 11(9):1374. https://doi.org/10.3390/biom11091374
Chicago/Turabian StyleSchilcher, Felix, Markus Thamm, Martin Strube-Bloss, and Ricarda Scheiner. 2021. "Opposing Actions of Octopamine and Tyramine on Honeybee Vision" Biomolecules 11, no. 9: 1374. https://doi.org/10.3390/biom11091374
APA StyleSchilcher, F., Thamm, M., Strube-Bloss, M., & Scheiner, R. (2021). Opposing Actions of Octopamine and Tyramine on Honeybee Vision. Biomolecules, 11(9), 1374. https://doi.org/10.3390/biom11091374