
Role of Fibroblasts and Myofibroblasts on the Pathogenesis and Treatment of Pelvic Organ Prolapse


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
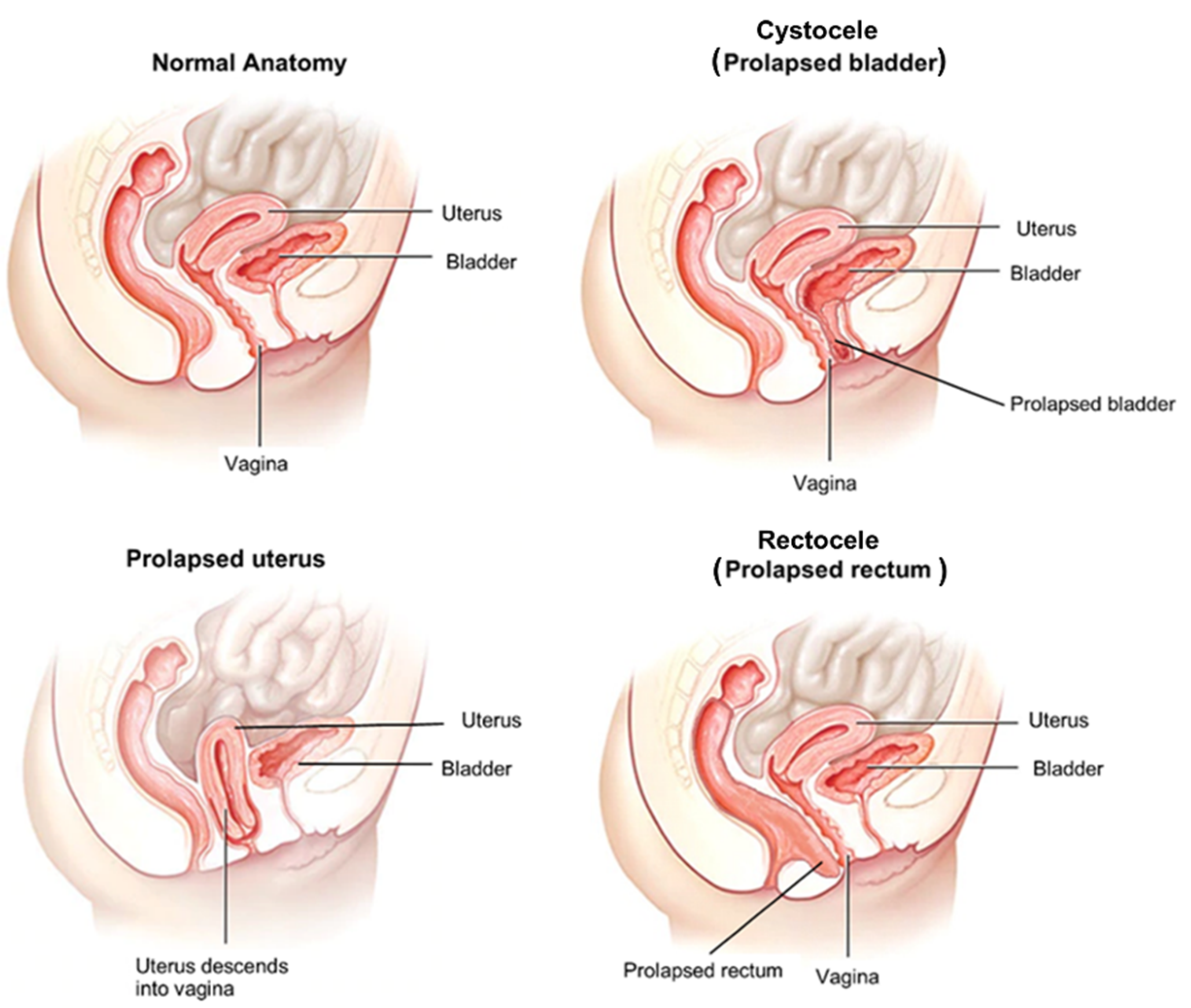
2. Pelvic Organ Prolapse as a Connective Tissue Disorder
2.1. Background/Cause
2.2. Changes in Connective Tissue in POP
3. Role of Fibroblasts in Wound Healing and Tissue Regeneration
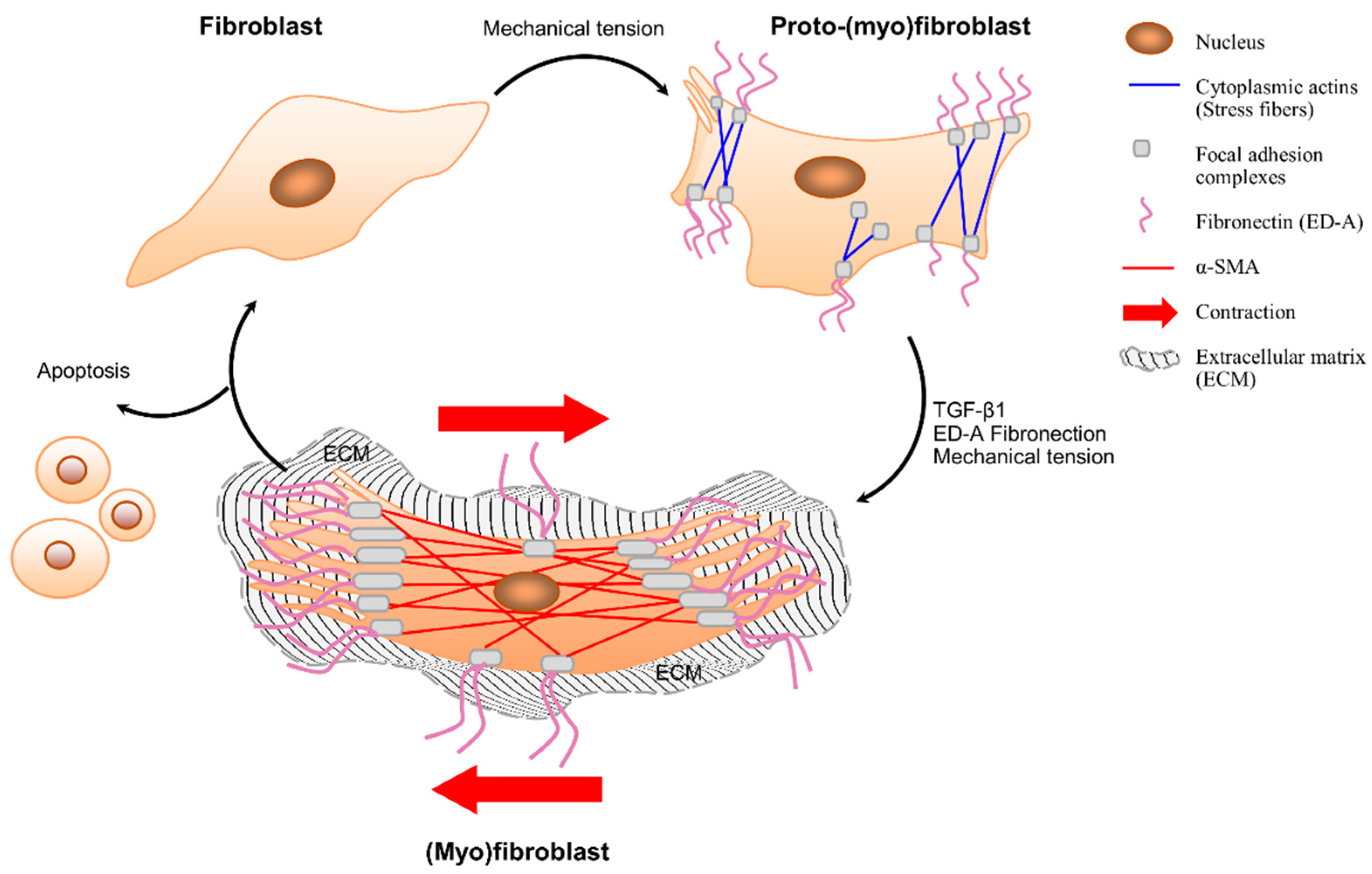
3.1. Fibroblast–(Myo)Fibroblast Transition
3.2. Unpaired Healing—Fibrosis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lakeman, M.M.; van der Vaart, C.H.; Laan, E.; Roovers, J.P.W. The effect of prolapse surgery on vaginal sensibility. J. Sex. Med. 2011, 8, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Digesu, G.A.; Chaliha, C.; Salvatore, S.; Hutchings, A.; Khullar, V. The relationship of vaginal prolapse severity tosymptoms and quality of life. BJOG Int. J. Obstet. Gynaecol. 2005, 112, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Groenendijk, A.G.; Birnie, E.; Roovers, J.-P.W.; Bonsel, G.J. Contribution of primary pelvic organ prolapse to micturition and defecation symptoms. Obstet. Gynecol. Int. 2012, 2012, 798035. [Google Scholar] [CrossRef] [PubMed]
- Medel, S.; Alarab, M.; Kufaishi, H.; Drutz, H.; Shynlova, O. Attachment of primary vaginal fibroblasts to absorbable and nonabsorbable implant materials coated with platelet-rich plasma: Potential application in pelvic organ prolapse surgery. Female Pelvic Med. Reconstr. Surg. 2015, 21, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Poncet, S.; Meyer, S.; Richard, C.; Aubert, J.-D.; Juillerat-Jeanneret, L. The expression and function of the endothelin system in contractile properties of vaginal myofibroblasts of women with uterovaginal prolapse. Am. J. Obstet. Gynecol. 2005, 192, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Mant, J.; Painter, R.; Vessey, M. Epidemiology of genital prolapse: Observations from the Oxford Family Planning Association Study. BJOG Int. J. Obstet. Gynaecol. 1997, 104, 579–585. [Google Scholar] [CrossRef]
- Twiss, C.; Triaca, V.; Rodríguez, L.V. Familial transmission of urogenital prolapse and incontinence. Curr. Opin. Obstet. Gynecol. 2007, 19, 464–468. [Google Scholar] [CrossRef]
- Lang, J.; Zhu, L.; Sun, Z.; Chen, J. Estrogen levels and estrogen receptors in patients with stress urinary incontinence and pelvic organ prolapse. Int. J. Gynecol. Obstet. 2003, 80, 35–39. [Google Scholar] [CrossRef]
- Diedrich, C.M.; Roovers, J.-P.; Smit, T.H.; Guler, Z. Fully absorbable poly-4-hydroxybutyrate implants exhibit more favorable cell-matrix interactions than polypropylene. Mater. Sci. Eng. C 2021, 120, 111702. [Google Scholar] [CrossRef]
- Ewies, A.A.; Al-Azzawi, F.; Thompson, J. Changes in extracellular matrix proteins in the cardinal ligaments of post-menopausal women with or without prolapse: A computerized immunohistomorphometric analysis. Hum. Reprod. 2003, 18, 2189–2195. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B. The myofibroblast in connective tissue repair and regeneration. In Regenerative Medicine and Biomaterials for the Repair of Connective Tissues; Elsevier: Amsterdam, The Netherlands, 2010; pp. 39–80. [Google Scholar]
- Jackson, S.; Eckford, S.; Abrams, P.; Avery, N.; Tarlton, J.; Bailey, A. Changes in metabolism of collagen in genitourinary prolapse. Lancet 1996, 347, 1658–1661. [Google Scholar] [CrossRef]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Kalkan, U.; Yoldemir, T.; Ozyurek, E.; Daniilidis, A. Native tissue repair versus mesh repair in pelvic organ prolapse surgery. Climacteric 2017, 20, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, M.; Familusi, A.; Fielding, S.; Ford, J.; Bhattacharya, S. Primary and repeat surgical treatment for female pelvic organ prolapse and incontinence in parous women in the UK: A register linkage study. BMJ Open 2011, 1, e000206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denman, M.A.; Gregory, W.T.; Boyles, S.H.; Smith, V.; Edwards, S.R.; Clark, A.L. Reoperation 10 Years After Surgically Managed Pelvic Organ Prolapse and Urinary Incontinence. Am. J. Obstet. Gynecol. 2008, 198, 555.e1–555.e5. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.L.; Smith, V.J.; Bergstrom, J.O.; Colling, J.C.; Clark, A.L. Epidemiology of Surgically Managed Pelvic Organ Prolapse and Urinary Incontinence. Obstet. Gynecol. 1997, 89, 501–506. [Google Scholar] [CrossRef]
- Birch, C.; Fynes, M.M. The role of synthetic and biological prostheses in reconstructive pelvic floor surgery. Curr. Opin. Obstet. Gynecol. 2002, 14, 527–535. [Google Scholar] [CrossRef]
- Altman, D.; Väyrynen, T.; Engh, M.E.; Axelsen, S.; Falconer, C. Anterior colporrhaphy versus transvaginal mesh for pelvic-organ prolapse. N. Engl. J. Med. 2011, 364, 1826–1836. [Google Scholar] [CrossRef] [Green Version]
- Sivaslioglu, A.; Unlubilgin, E.; Dolen, I. A randomized comparison of polypropylene mesh surgery with site-specific surgery in the treatment of cystocoele. Int. Urogynecol. J. 2008, 19, 467–471. [Google Scholar] [CrossRef]
- Vollebregt, A.; Fischer, K.; Gietelink, D.; Van der Vaart, C. Primary surgical repair of anterior vaginal prolapse: A randomised trial comparing anatomical and functional outcome between anterior colporrhaphy and trocar-guided transobturator anterior mesh. BJOG Int. J. Obstet. Gynaecol. 2011, 118, 1518–1527. [Google Scholar] [CrossRef]
- De Tayrac, R.; Cornille, A.; Eglin, G.; Guilbaud, O.; Mansoor, A.; Alonso, S.; Fernandez, H. Comparison between trans-obturator trans-vaginal mesh and traditional anterior colporrhaphy in the treatment of anterior vaginal wall prolapse: Results of a French RCT. Int. Urogynecol. J. 2013, 24, 1651–1661. [Google Scholar] [CrossRef]
- Ramanah, R.; Mairot, J.; Clement, M.-C.; Parratte, B.; Maillet, R.; Riethmuller, D. Evaluating the porcine dermis graft InteXen® in three-compartment transvaginal pelvic organ prolapse repair. Int. Urogynecol. J. 2010, 21, 1151–1156. [Google Scholar] [CrossRef]
- Mahdy, A.; Karp, D.; Davila, G.W.; Ghoniem, G.M. The outcome of transobturator anterior vaginal wall prolapse repair using porcine dermis graft: Intermediate term follow-up. Int. Braz. J. Urol. 2013, 39, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Winters, J.C. InteXen tissue processing and laboratory study. Int. Urogynecol. J. 2006, 17, 34–38. [Google Scholar] [CrossRef]
- Liang, R.; Abramowitch, S.; Knight, K.; Palcsey, S.; Nolfi, A.; Feola, A.; Stein, S.; Moalli, P.A. Vaginal degeneration following implantation of synthetic mesh with increased stiffness. BJOG Int. J. Obstet. Gynaecol. 2013, 120, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Verhorstert, K.W.; Gudde, A.N.; Kortz, B.S.; Limpens, J.; Roovers, J.P.W.; Hooijmans, C.R.; Guler, Z. Animal experimental research assessing urogynecologic surgical mesh implants: Outcome measures describing the host response, a systematic review and meta-analysis. Neurourol. Urodyn. 2021, 40, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Diedrich, C.M.; Guler, Z.; Hympanova, L.; Vodegel, E.; Zündel, M.; Mazza, E.; Deprest, J.; Roovers, J.P. Evaluation of the short term host response and biomechanics of an absorbable poly-4-hydroxybutyrate scaffold in a sheep model following vaginal implantation. BJOG Int. J. Obstet. Gynaecol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Artsen, A.M.; Rytel, M.; Liang, R.; King, G.E.; Meyn, L.; Abramowitch, S.D.; Moalli, P.A. Mesh induced fibrosis: The protective role of T regulatory cells. Acta Biomater. 2019, 96, 203–210. [Google Scholar] [CrossRef]
- Raizada, V.; Mittal, R.K. Pelvic floor anatomy and applied physiology. Gastroenterol. Clin. N. Am. 2008, 37, 493–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visco, A.G.; Yuan, L. Differential gene expression in pubococcygeus muscle from patients with pelvic organ prolapse. Am. J. Obstet. Gynecol. 2003, 189, 102–112. [Google Scholar] [CrossRef]
- Jelovsek, J.E.; Maher, C.; Barber, M.D. Pelvic organ prolapse. Lancet 2007, 369, 1027–1038. [Google Scholar] [CrossRef]
- Min, J.; Li, B.; Liu, C.; Guo, W.; Hong, S.; Tang, J.; Hong, L. Extracellular matrix metabolism disorder induced by mechanical strain on human parametrial ligament fibroblasts. Mol. Med. Rep. 2017, 15, 3278–3284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Shangguan, A.; Kujawa, S.; Bochenska, K.; Zhang, L.; Zhao, H. Estrogen and pelvic organ prolapse. J. Mol. Genet. Med. 2016, 10, 1747-0862.1000221. [Google Scholar] [CrossRef]
- Weber, M.; Kleijn, M.; Langendam, M.; Limpens, J.; Heineman, M.; Roovers, J. Local oestrogen for pelvic floor disorders: A systematic review. PLoS ONE 2015, 10, e0136265. [Google Scholar]
- Soloyan, H.; De Filippo, R.E.; Sedrakyan, S. Tissue Engineering of the Reproductive System; Academic Press; Elsevier: Burlington, MA, USA, 2019. [Google Scholar]
- Ewies, A.A.; Elshafie, M.; Li, J.; Stanley, A.; Thompson, J.; Styles, J.; White, I.; Al-Azzawi, F. Changes in transcription profile and cytoskeleton morphology in pelvic ligament fibroblasts in response to stretch: The effects of estradiol and levormeloxifene. MHR-Basic Sci. Reprod. Med. 2008, 14, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Q.-Y.; Sun, B.-F.; Ma, Y.; Zhang, Y.; Wang, M.; Ma, C.; Shi, H.; Sun, Z.; Chen, J. Single-cell transcriptome profiling of the vaginal wall in women with severe anterior vaginal prolapse. Nat. Commun. 2021, 12, 87. [Google Scholar] [CrossRef]
- Kufaishi, H.; Alarab, M.; Drutz, H.; Lye, S.; Shynlova, O. Comparative characterization of vaginal cells derived from premenopausal women with and without severe pelvic organ prolapse. Reprod. Sci. 2016, 23, 931–943. [Google Scholar] [CrossRef]
- Reddy, K.; Mangale, S.S. Integrin receptors: The dynamic modulators of endometrial function. Tissue Cell 2003, 35, 260–273. [Google Scholar] [CrossRef]
- Kerkhof, M.; Hendriks, L.; Brölmann, H. Changes in connective tissue in patients with pelvic organ prolapse—A review of the current literature. Int. Urogynecol. J. 2009, 20, 461–474. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.Y.; Harmanli, O.H.; Agar, M.; Dandolu, V.; Grody, M.T. Collagen content of nonsupport tissue in pelvic organ prolapse and stress urinary incontinence. Am. J. Obstet. Gynecol. 2003, 189, 1597–1599. [Google Scholar] [CrossRef]
- Moalli, P.A.; Shand, S.H.; Zyczynski, H.M.; Gordy, S.C.; Meyn, L.A. Remodeling of vaginal connective tissue in patients with prolapse. Obstet. Gynecol. 2005, 106, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.J. Molecular mechanisms of ageing in connective tissues. Mech. Ageing Dev. 2001, 122, 735–755. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, R.; Li, H.; Gu, Y.; Wei, W. Expression and significance of metalloproteinase and collagen in vaginal wall tissues of patients with pelvic organ prolapse. Ann. Clin. Lab. Sci. 2017, 47, 698–705. [Google Scholar] [PubMed]
- Goh, J.T. Biomechanical and biochemical assessments for pelvic organ prolapse. Curr. Opin. Obstet. Gynecol. 2003, 15, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, M.W.; Falconer, C.; Byström, B.; Malmström, A.; Ekman, G. Young women with genital prolapse have a low collagen concentration. Acta Obstet. Gynecol. Scand. 2004, 83, 1193–1198. [Google Scholar] [PubMed]
- Gabriel, B.; Denschlag, D.; Göbel, H.; Fittkow, C.; Werner, M.; Gitsch, G.; Watermann, D. Uterosacral ligament in postmenopausal women with or without pelvic organ prolapse. Int. Urogynecol. J. 2005, 16, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Zapata, A.M.; Kerkhof, M.H.; Zandieh-Doulabi, B.; Brölmann, H.A.; Smit, T.H.; Helder, M.N. Fibroblasts from women with pelvic organ prolapse show differential mechanoresponses depending on surface substrates. Int. Urogynecol. J. 2013, 24, 1567–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarasa-Renedo, A.; Chiquet, M. Mechanical signals regulating extracellular matrix gene expression in fibroblasts. Scand. J. Med. Sci. Sports 2005, 15, 223–230. [Google Scholar] [CrossRef]
- Soo, C.; Shaw, W.W.; Zhang, X.; Longaker, M.T.; Howard, E.W.; Ting, K. Differential expression of matrix metalloproteinases and their tissue-derived inhibitors in cutaneous wound repair. Plast. Reconstr. Surg. 2000, 105, 638–647. [Google Scholar] [CrossRef]
- Ra, H.-J.; Parks, W.C. Control of matrix metalloproteinase catalytic activity. Matrix Biol. 2007, 26, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, B.; Watermann, D.; Hancke, K.; Gitsch, G.; Werner, M.; Tempfer, C.; Zur Hausen, A. Increased expression of matrix metalloproteinase 2 in uterosacral ligaments is associated with pelvic organ prolapse. Int. Urogynecol. J. 2006, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Rahajeng, R. The increased of MMP-9 and MMP-2 with the decreased of TIMP-1 on the uterosacral ligament after childbirth. Pan Afr. Med. J. 2018, 30, 283. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Chen, J.; Guo, X.; Guan, H.; Li, C. Differential expression profiling of matrix metalloproteinases and tissue inhibitors of metalloproteinases in females with or without pelvic organ prolapse. Mol. Med. Rep. 2014, 10, 2004–2008. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.M.; Visco, A.G.; Grass, E.A.; Craig, D.M.; Fulton, R.G.; Haynes, C.; Weidner, A.C.; Shah, S.H. Matrix metalloproteinase-9 genetic polymorphisms and the risk for advanced pelvic organ prolapse. Obstet. Gynecol. 2012, 120, 587. [Google Scholar] [CrossRef] [PubMed]
- Campeau, L.; Gorbachinsky, I.; Badlani, G.H.; Andersson, K.E. Pelvic floor disorders: Linking genetic risk factors to biochemical changes. BJU Int. 2011, 108, 1240–1247. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Pawlyk, B.; Damaser, M.; Li, T. Failure of elastic fiber homeostasis leads to pelvic floor disorders. Am. J. Pathol. 2006, 168, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.h.; Zhou, J.h. Decreased expression of elastin, fibulin-5 and lysyl oxidase-like 1 in the uterosacral ligaments of postmenopausal women with pelvic organ prolapse. J. Obstet. Gynaecol. Res. 2012, 38, 925–931. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yamamoto, M.; Akazawa, K.; Tajima, S.; Wakimoto, H.; Aoyagi, M. Decrease in elastin gene expression and protein synthesis in fibroblasts derived from cardinal ligaments of patients with prolapsus uteri. Cell Biol. Int. 1997, 21, 605–611. [Google Scholar] [CrossRef]
- Chen, B.; Wen, Y.; Polan, M.L. Elastolytic activity in women with stress urinary incontinence and pelvic organ prolapse. Neurourol. Urodyn. Off. J. Int. Cont. Soc. 2004, 23, 119–126. [Google Scholar] [CrossRef]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Ko, U.H.; Choi, J.; Choung, J.; Moon, S.; Shin, J.H. Physicochemically tuned myofibroblasts for wound healing strategy. Sci. Rep. 2019, 9, 16070. [Google Scholar] [CrossRef] [Green Version]
- Porter, S. The role of the fibroblast in wound contraction and healing. Wounds UK 2007, 3, 33. [Google Scholar]
- Hinz, B.; Gabbiani, G. Cell-matrix and cell-cell contacts of myofibroblasts: Role in connective tissue remodeling. Thromb. Haemost. 2003, 90, 993–1002. [Google Scholar]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, G.B.; Cliff, W.J.; Gabbiani, G.; Irle, C.; Montandon, D.; Statkov, P.R.; Majno, G. Myofibroblasts in human granulation tissue. Hum. Pathol. 1974, 5, 55–67. [Google Scholar] [CrossRef]
- Desmouliere, A.; Redard, M.; Darby, I.; Gabbiani, G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am. J. Pathol. 1995, 146, 56. [Google Scholar]
- Eastwood, M.; Mudera, V.; McGrouther, D.; Brown, R. Effect of precise mechanical loading on fibroblast populated collagen lattices: Morphological changes. Cell Motil. Cytoskelet. 1998, 40, 13–21. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Z.; Lü, D.; Xu, Q. Effects of mechanical stretching on the morphology and cytoskeleton of vaginal fibroblasts from women with pelvic organ prolapse. Int. J. Mol. Sci. 2015, 16, 9406–9419. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301. [Google Scholar]
- Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef]
- Grant, C.A.; Twigg, P.C.; Tobin, D.J. Static and dynamic nanomechanical properties of human skin tissue using atomic force microscopy: Effect of scarring in the upper dermis. Acta Biomater. 2012, 8, 4123–4129. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Zapata, A.M.; Feola, A.J.; Heesakkers, J.; de Graaf, P.; Blaganje, M.; Sievert, K.-D. Biomechanical properties of the pelvic floor and its relation to pelvic floor disorders. Eur. Urol. Suppl. 2018, 17, 80–90. [Google Scholar] [CrossRef]
- Hayakawa, T.; Hashimoto, Y.; Myokei, Y.; Aoyama, H.; Izawa, Y. Changes in type of collagen during the development of human post-burn hypertrophic scars. Clin. Chim. Acta 1979, 93, 119–125. [Google Scholar] [CrossRef]
- Meyer, S.; Achtari, C.; Hohlfeld, P.; Juillerat-Jeanneret, L. The contractile properties of vaginal myofibroblasts: Is the myofibroblasts contraction force test a valuable indication of future prolapse development? Int. Urogynecol. J. 2008, 19, 1399–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Zapata, A.M.; Heinz, A.; Kerkhof, M.H.; van de Westerlo-van Rijt, C.; Schmelzer, C.E.; Stoop, R.; Kluivers, K.B.; Oosterwijk, E. Extracellular Matrix Stiffness and Composition Regulate the Myofibroblast Differentiation of Vaginal Fibroblasts. Int. J. Mol. Sci. 2020, 21, 4762. [Google Scholar] [CrossRef] [PubMed]
- Barone, W.R.; Moalli, P.A.; Abramowitch, S.D. Textile properties of synthetic prolapse mesh in response to uniaxial loading. Am. J. Obstet. Gynecol. 2016, 215, 326.e1–326.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobb, W.; Peindl, R.; Zerey, M.; Carbonell, A.; Heniford, B. Mesh terminology 101. Hernia 2009, 13, 1–6. [Google Scholar] [CrossRef]
- Liang, R.; Knight, K.; Abramowitch, S.; Moalli, P.A. Exploring the basic science of prolapse meshes. Curr. Opin. Obstet. Gynecol. 2016, 28, 413. [Google Scholar] [CrossRef]
- Nolfi, A.L.; Brown, B.N.; Liang, R.; Palcsey, S.L.; Bonidie, M.J.; Abramowitch, S.D.; Moalli, P.A. Host response to synthetic mesh in women with mesh complications. Am. J. Obstet. Gynecol. 2016, 215, 206.e1–206.e8. [Google Scholar] [CrossRef] [Green Version]
- Vashaghian, M.; Zandieh-Doulabi, B.; Roovers, J.-P.; Smit, T.H. Electrospun matrices for pelvic floor repair: Effect of fiber diameter on mechanical properties and cell behavior. Tissue Eng. Part A 2016, 22, 1305–1316. [Google Scholar] [CrossRef]
- Hjort, H.; Mathisen, T.; Alves, A.; Clermont, G.; Boutrand, J. Three-year results from a preclinical implantation study of a long-term resorbable surgical mesh with time-dependent mechanical characteristics. Hernia 2012, 16, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangera, A.; Bullock, A.J.; Roman, S.; Chapple, C.R.; MacNeil, S. Comparison of candidate scaffolds for tissue engineering for stress urinary incontinence and pelvic organ prolapse repair. BJU Int. 2013, 112, 674–685. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Sun, X.; Li, Q.; Ding, X.; Liu, H.; Wang, J. Silk fibroin scaffold as a potential choice for female pelvic reconstruction: A study on the biocompatibility in abdominal wall, pelvic, and vagina. Microsc. Res. Tech. 2017, 80, 291–297. [Google Scholar] [CrossRef]
- Vashaghian, M.; Ruiz-Zapata, A.M.; Kerkhof, M.H.; Zandieh-Doulabi, B.; Werner, A.; Roovers, J.P.; Smit, T.H. Toward a new generation of pelvic floor implants with electrospun nanofibrous matrices: A feasibility study. Neurourol. Urodyn. 2017, 36, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Y.; Zhu, C.; Tong, X.; Yang, M.; Yang, L.; Liu, Z.; Huang, W.; Wu, F.; Zong, H. Preclinical animal study and human clinical trial data of co-electrospun poly (L-lactide-co-caprolactone) and fibrinogen mesh for anterior pelvic floor reconstruction. Int. J. Nanomed. 2016, 11, 389. [Google Scholar]
- Barone, W.R.; Amini, R.; Maiti, S.; Moalli, P.A.; Abramowitch, S.D. The impact of boundary conditions on surface curvature of polypropylene mesh in response to uniaxial loading. J. Biomech. 2015, 48, 1566–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, J.; Kaldenhoff, E.; Kirschner-Hermanns, R.; Mühl, T.; Klinge, U. Elongation of textile pelvic floor implants under load is related to complete loss of effective porosity, thereby favoring incorporation in scar plates. J. Biomed. Mater. Res. Part A 2014, 102, 1079–1084. [Google Scholar] [CrossRef]
- Vashaghian, M.; Diedrich, C.M.; Zandieh-Doulabi, B.; Werner, A.; Smit, T.H.; Roovers, J.-P. Gentle cyclic straining of human fibroblasts on electrospun scaffolds enhances their regenerative potential. Acta Biomater. 2019, 84, 159–168. [Google Scholar] [CrossRef]
- Jia, Y.; Zhou, J.; Chang, Y.; An, F. The effect of optimized concentrations of bFGF and EGF on the proliferation of fibroblasts and the expression of collagen: Related to pelvic floor tissue regeneration. Chin. Med. J. 2018, 131, 2089–2096. [Google Scholar] [CrossRef]
- Wu, X.; Jia, Y.; Sun, X.; Wang, J. Tissue engineering in female pelvic floor reconstruction. Eng. Life Sci. 2020, 20, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Caruso, S.; Cianci, S.; Vitale, S.G.; Matarazzo, M.G.; Amore, F.F.; Cianci, A. Effects of ultralow topical estriol dose on vaginal health and quality of life in postmenopausal women who underwent surgical treatment for pelvic organ prolapse. Menopause 2017, 24, 900–907. [Google Scholar] [CrossRef]
- Guo, S.A.; DiPietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef]
- Martin, P. Wound healing—Aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Klingberg, F.; Hinz, B.; White, E.S. The myofibroblast matrix: Implications for tissue repair and fibrosis. J. Pathol. 2013, 229, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Vasin, R.; Filimonov, V.; Mnikhovich, M.; Kaprin, A.; Kostin, A.; Vasina, I. Morphologic structure and immunohistochemical analysis of vaginal wall in women with pelvic organ prolapse. Urologiia 2019, 6, 12–20. [Google Scholar] [CrossRef]
- Kulasekaran, P.; Scavone, C.A.; Rogers, D.S.; Arenberg, D.A.; Thannickal, V.J.; Horowitz, J.C. Endothelin-1 and transforming growth factor-β1 independently induce fibroblast resistance to apoptosis via AKT activation. Am. J. Respir. Cell Mol. Biol. 2009, 41, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Arpino, V.; Brock, M.; Gill, S.E. The role of TIMPs in regulation of extracellular matrix proteolysis. Matrix Biol. 2015, 44, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Bochaton-Piallat, M.-L.; Gabbiani, G.; Hinz, B. The myofibroblast in wound healing and fibrosis: Answered and unanswered questions. F1000Research 2016, 5, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szauter, K.M.; Cao, T.; Boyd, C.D.; Csiszar, K. Lysyl oxidase in development, aging and pathologies of the skin. Pathol. Biol. 2005, 53, 448–456. [Google Scholar] [CrossRef]
- Alarab, M.; Bortolini, M.A.; Drutz, H.; Lye, S.; Shynlova, O. LOX family enzymes expression in vaginal tissue of premenopausal women with severe pelvic organ prolapse. Int. Urogynecol. J. 2010, 21, 1397–1404. [Google Scholar] [CrossRef]
- Kufaishi, H. Vaginal Fibroblasts Derived from Premenopausal Women with and without Severe Pelvic Organ Prolapse: Differential Characteristics and Effect of Mechanical Stretch; ProQuest LLC: Ann Arbor, MI, USA, 2015. [Google Scholar]
- Annes, J.P.; Chen, Y.; Munger, J.S.; Rifkin, D.B. Integrin αVβ6-mediated activation of latent TGF-β requires the latent TGF-β binding protein-1. J. Cell Biol. 2004, 165, 723–734. [Google Scholar] [CrossRef]
- Froese, A.R.; Shimbori, C.; Bellaye, P.-S.; Inman, M.; Obex, S.; Fatima, S.; Jenkins, G.; Gauldie, J.; Ask, K.; Kolb, M. Stretch-induced activation of transforming growth factor-β1 in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2016, 194, 84–96. [Google Scholar] [CrossRef]
- Cook, H.; Stephens, P.; Davies, K.J.; Thomas, D.W.; Harding, K.G. Defective extracellular matrix reorganization by chronic wound fibroblasts is associated with alterations in TIMP-1, TIMP-2, and MMP-2 activity. J. Investig. Dermatol. 2000, 115, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Des Jardins-Park, H.E.; Foster, D.S.; Longaker, M.T. Fibroblasts and wound healing: An update. Regen. Med. 2018, 13, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Darzi, S.; Werkmeister, J.A.; Gargett, C.E.; Mukherjee, S. Emerging nano/micro-structured degradable polymeric meshes for pelvic floor reconstruction. Nanomaterials 2020, 10, 1120. [Google Scholar] [CrossRef] [PubMed]
- Feola, A.; Endo, M.; Urbankova, I.; Vlacil, J.; Deprest, T.; Bettin, S.; Klosterhalfen, B.; Deprest, J. Host reaction to vaginally inserted collagen containing polypropylene implants in sheep. Am. J. Obstet. Gynecol. 2015, 212, 474.e1–474.e8. [Google Scholar] [CrossRef] [PubMed]
- Todros, S.; Pavan, P.; Pachera, P.; Natali, A. Synthetic surgical meshes used in abdominal wall surgery: Part II—Biomechanical aspects. J. Biomed. Mater. Res. Part B Appl. Biomater. 2017, 105, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Diegelmann, R.F.; Evans, M.C. Wound healing: An overview of acute, fibrotic and delayed healing. Front. Biosci. 2004, 9, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Feola, A.; Abramowitch, S.; Jallah, Z.; Stein, S.; Barone, W.; Palcsey, S.; Moalli, P. Deterioration in biomechanical properties of the vagina following implantation of a high-stiffness prolapse mesh. BJOG Int. J. Obstet. Gynaecol. 2013, 120, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.; Macdougall, K.; Olabisi, O.; McGuire, N. In vivo response to polypropylene following implantation in animal models: A review of biocompatibility. Int. Urogynecol. J. 2017, 28, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.A.; Shepherd, J.P.; Oliphant, S.S.; Wang, L.; Bunker, C.H.; Lowder, J.L. Trends in inpatient prolapse procedures in the United States, 1979–2006. Am. J. Obstet. Gynecol. 2010, 202, 501.e1–501.e7. [Google Scholar] [CrossRef] [Green Version]
- Orenstein, S.B.; Saberski, E.R.; Kreutzer, D.L.; Novitsky, Y.W. Comparative analysis of histopathologic effects of synthetic meshes based on material, weight, and pore size in mice. J. Surg. Res. 2012, 176, 423–429. [Google Scholar] [CrossRef]
- Couri, B.M.; Lenis, A.T.; Borazjani, A.; Paraiso, M.F.R.; Damaser, M.S. Animal models of female pelvic organ prolapse: Lessons learned. Expert Rev. Obstet. Gynecol. 2012, 7, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, N.; Rosamilia, A.; Arkwright, J.; Lee, J.; Davies-Tuck, M.; Melendez, J.; Werkmeister, J.; Gargett, C.E. Vaginal wall weakness in parous ewes: A potential preclinical model of pelvic organ prolapse. Int. Urogynecol. J. 2017, 28, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Hympánová, L.; Rynkevic, R.; Román, S.; da Cunha, M.G.M.; Mazza, E.; Zündel, M.; Urbánková, I.; Gallego, M.R.; Vange, J.; Callewaert, G. Assessment of electrospun and ultra-lightweight polypropylene meshes in the sheep model for vaginal surgery. Eur. Urol. Focus 2020, 6, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Tayrac, R.; Chentouf, S.; Garreau, H.; Braud, C.; Guiraud, I.; Boudeville, P.; Vert, M. In vitro degradation and in vivo biocompatibility of poly (lactic acid) mesh for soft tissue reinforcement in vaginal surgery. J. Biomed. Mater. Res. Part B Appl. Biomater. 2008, 85, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Mangir, N.; Dikici, B.A.; Chapple, C.R.; MacNeil, S. Landmarks in vaginal mesh development: Polypropylene mesh for treatment of SUI and POP. Nat. Rev. Urol. 2019, 16, 675–689. [Google Scholar] [CrossRef] [Green Version]
- Grinnell, F.; Ho, C.-H.; Lin, Y.-C.; Skuta, G. Differences in the regulation of fibroblast contraction of floating versus stressed collagen matrices. J. Biol. Chem. 1999, 274, 918–923. [Google Scholar] [CrossRef] [Green Version]
- Wipff, P.-J.; Rifkin, D.B.; Meister, J.-J.; Hinz, B. Myofibroblast contraction activates latent TGF-β1 from the extracellular matrix. J. Cell Biol. 2007, 179, 1311–1323. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guler, Z.; Roovers, J.P. Role of Fibroblasts and Myofibroblasts on the Pathogenesis and Treatment of Pelvic Organ Prolapse. Biomolecules 2022, 12, 94. https://doi.org/10.3390/biom12010094
Guler Z, Roovers JP. Role of Fibroblasts and Myofibroblasts on the Pathogenesis and Treatment of Pelvic Organ Prolapse. Biomolecules. 2022; 12(1):94. https://doi.org/10.3390/biom12010094
Chicago/Turabian StyleGuler, Zeliha, and Jan Paul Roovers. 2022. "Role of Fibroblasts and Myofibroblasts on the Pathogenesis and Treatment of Pelvic Organ Prolapse" Biomolecules 12, no. 1: 94. https://doi.org/10.3390/biom12010094
APA StyleGuler, Z., & Roovers, J. P. (2022). Role of Fibroblasts and Myofibroblasts on the Pathogenesis and Treatment of Pelvic Organ Prolapse. Biomolecules, 12(1), 94. https://doi.org/10.3390/biom12010094