First Detection of Meloidogyne luci (Nematoda: Meloidogynidae) Parasitizing Potato in the Azores, Portugal



Abstract
:1. Introduction
2. Results and Discussion
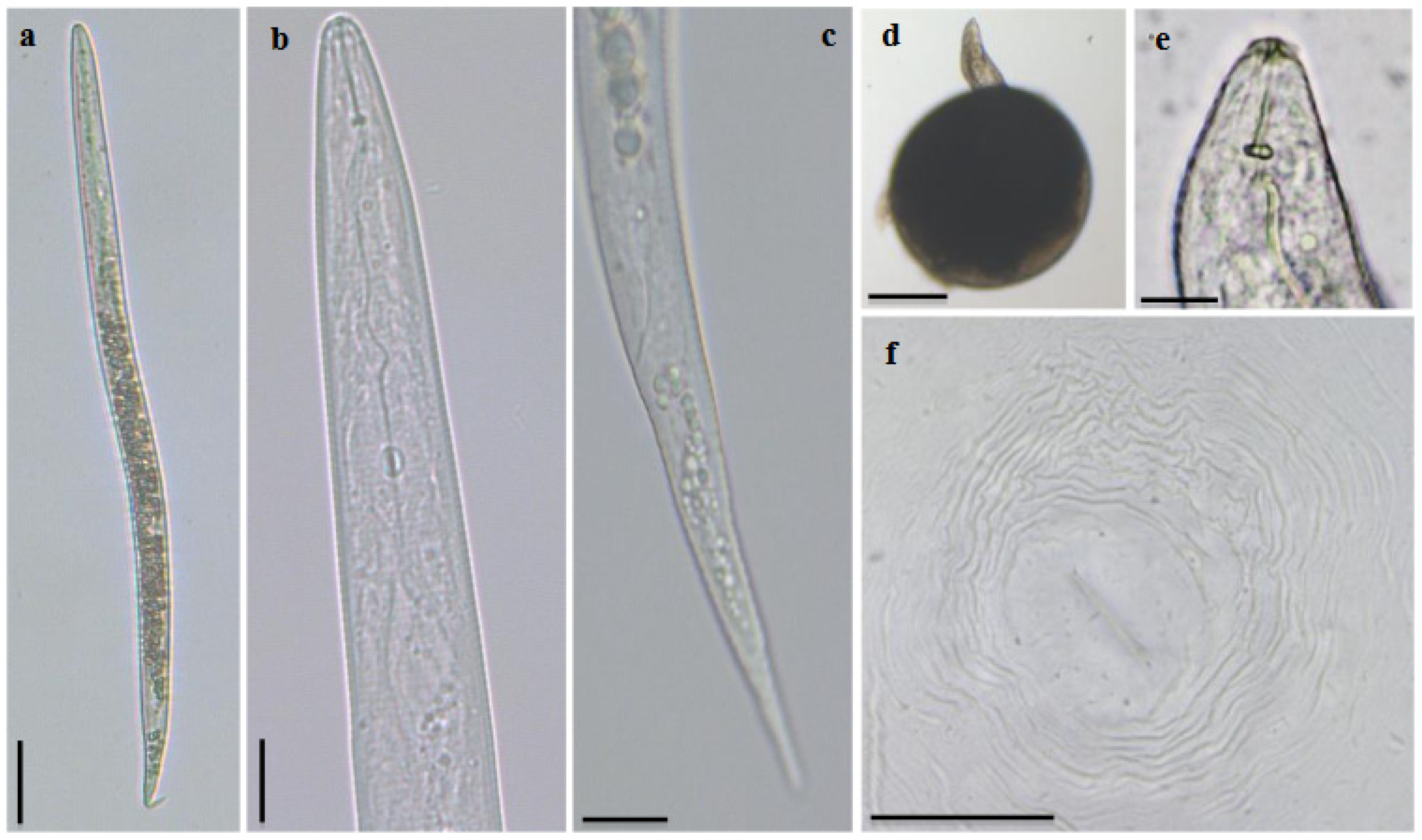
2.1. Morphological and Morphometric Characterization
2.2. Biochemical Characterization
2.3. Molecular Characterization
3. Materials and Methods
3.1. Nematode Isolates
3.2. Morphological and Morphometric Characterization
3.3. Biochemical Characterization
3.4. Molecular Characterization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT. 2020. Available online: http://www.fao.org/faostat/en/#data (accessed on 16 October 2020).
- Camacho, M.J.; Nóbrega, F.; Lima, A.; Mota, M.; Inácio, M.L. Morphological and molecular identification of the potato cyst nematodes Globodera rostochiensis and G. pallida in Portuguese potato fields. Nematology 2017, 8, 883–889. [Google Scholar] [CrossRef]
- Lima, F.S.O.; Mattos, S.V.; Silva, S.E.; Carvalho, M.A.S.; Teixeira, R.A.; Silva, J.C.; Correa, V.R. Nematodes Affecting Potato and Sustainable Practices for Their Management. In Potato—From Incas to All Over the World, 1st ed.; Yildiz, M., Ed.; IntechOpen: London, UK, 2018; Volume 1, pp. 107–121. [Google Scholar]
- Dong, K.; Dean, R.A.; Fortnum, B.A.; Lewis, S.A. Development of PCR primers to identify species of root knot nematodes: Meloidogyne arenaria, M. hapla, M. incognta and M. Javanica. Nematropica 2001, 31, 271–280. [Google Scholar]
- Trudgill, D.L.; Blok, V.C. Apomictic, polyphagous root-knot nematodes: Exceptionally successful and damaging biotrophic root pathogens. Annu. Rev. Phytopathol. 2001, 39, 53–77. [Google Scholar] [CrossRef] [PubMed]
- Hunt, D.; Handoo, Z. Taxonomy, identification and principal species. In Root—Knot Nematodes; Perry, R.N., Moens, M., Starr, J.L., Eds.; CABI: London, UK, 2009; pp. 55–88. [Google Scholar]
- Abrantes, I.D.; dos Santos, M.V.; da Conceição, I.L.; Santos, M.D.; Vovlas, N. Root-knot and other plant-parasitic nematodes associated with fig trees in Portugal. Nema. Mediterr. 2008, 36, 131–136. [Google Scholar]
- Abrantes, I.D.; Santos, M.S.; Correia, M.; da Conceição, I.L.; da Cunha, M.J.; Feio, G.; dos Santos, M.C. Root-knot nematodes, Meloidogyne spp., on potato in Portugal. Nematology 2009, 11, 311–313. [Google Scholar] [CrossRef] [Green Version]
- Maleita, C.; Esteves, I.; Cardoso, J.M.S.; Cunha, M.J.; Carneiro, R.M.D.G.; Abrantes, I. Meloidogyne luci, a new root-knot nematode parasitizing potato in Portugal. Plant Pathol. 2018, 67, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Santos, D.; Correia, A.; Abrantes, I.; Maleita, C. The quarantine root knot nematode Meloidogyne enterolobii—A potential threat to Portugal and Europe. Plant Pathol. 2019, 68, 1607–1615. [Google Scholar] [CrossRef]
- Viera dos Santos, M.; Almeida, M.T.M.; Costa, S.R. First report of Meloidogyne naasi parasitizing turfgrass in Portugal. J. Nematol. 2020, 52, 1–4. [Google Scholar] [CrossRef]
- Carneiro, R.M.D.G.; Correa, V.R.; Almeida, M.R.A.; Gomes, A.C.M.M.; Deimi, A.M.; Castagnone-Sereno, P.; Karssen, G. Meloidogyne luci n. sp. (Nematoda: Meloidogynidae), a root-knot nematode parasitising different crops in Brazil, Chile and Iran. Nematology 2014, 16, 289–301. [Google Scholar] [CrossRef]
- Santos, D.; Correia, A.; Abrantes, I.; Maleita, C. New Hosts and Records in Portugal for the Root-Knot Nematode Meloidogyne luci. J. Nematol. 2019, 51, 1–4. [Google Scholar] [CrossRef] [Green Version]
- EPPO Alert List: Addition of Meloidogyne luci together with Meloidogyne ethiopica. Available online: https://gd.eppo.int/reporting/article-6186 (accessed on 23 November 2017).
- Esbenshade, P.R.; Triantaphyllou, A.C. Identification of major Meloidogyne species employing enzyme phenotypes as differentiating characters. In An Advanced Treatise on Meloidogyne; Sasser, J.N., Carter, C.C., Eds.; North Carolina State University Graphics: Raleigh, NC, USA, 1985; Volume I, pp. 135–140. [Google Scholar]
- Carneiro, R.M.D.G.; Almeida, M.R.A.; Quénéhervé, P. Enzyme phenotypes of Meloidogyne spp. isolates. Nematology 2000, 2, 645–654. [Google Scholar] [CrossRef]
- Stare, B.G.; Strajnar, P.; Susič, N.; Urek, G.; Širca, S. Reported populations of Meloidogyne ethiopica in Europe identified as Meloidogyne luci. Plan. Dis. 2017, 101, 1627–1632. [Google Scholar] [CrossRef] [Green Version]
- Standard Protocol PM 7/119 (1). Nematode Extraction; EPPO Bulletin 43:471-95; EPPO: Paris, France, 2013.
- Pais, C.S.; Abrantes, I.D.; Fernandes, M.F.; Santos, M.D. Técnica de electroforese aplicada ao estudo das enzimas dos nematodes-das-galhas-radiculares, Meloidogyne spp. Ciência Biol. Ecol. Syst. 1986, 6, 19–34. [Google Scholar]
- Powers, T.O.; Harris, T.S. A polymerase chain reaction method for identification of five major Meloidogyne species. J. Nematol. 1993, 25, 1–6. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Character/Ratio | M. luci J2 (Azores) (n = 10) | Carneiro et al., 2014 (n = 30) |
|---|---|---|
| Length | 404.99 ± 23 (376.3–446) | 383 ± 85 (300–470) |
| Stylet length | 14.05 ± 1 (11.4–15.6) | 12.5 ± 0.2 (12.0–13.5) |
| Dorsal oesophageal gland | 3.04 ± 0.41 (2.2–3.4) | 2.9 ± 0.5 (2.3–3.3) |
| Tail length | 46.64± 5.92 (39.7–54.1) | 44 ± 4.5 (40.0–48.5) |
| Hyaline terminus | 11.23 ± 1.97 (9.02–14.06) | 11.7 ± 3.0 (9–15) |
| Max. body width | 15.93 ± 2.26 (14.6–20.9) | 16 ± 1.5 (13–20) |
| a * | 25.76 ± 2.87 (22.2–31.9) | 25.6 ± 10.5 (15.0–36.1) |
| c ** | 8.77 ± 0.86 (7.5–9.6) | 8.7 ± 2.6 (6.2–11.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusinque, L.; Nóbrega, F.; Cordeiro, L.; Serra, C.; Inácio, M.L. First Detection of Meloidogyne luci (Nematoda: Meloidogynidae) Parasitizing Potato in the Azores, Portugal. Plants 2021, 10, 99. https://doi.org/10.3390/plants10010099
Rusinque L, Nóbrega F, Cordeiro L, Serra C, Inácio ML. First Detection of Meloidogyne luci (Nematoda: Meloidogynidae) Parasitizing Potato in the Azores, Portugal. Plants. 2021; 10(1):99. https://doi.org/10.3390/plants10010099
Chicago/Turabian StyleRusinque, Leidy, Filomena Nóbrega, Laura Cordeiro, Clara Serra, and Maria L. Inácio. 2021. "First Detection of Meloidogyne luci (Nematoda: Meloidogynidae) Parasitizing Potato in the Azores, Portugal" Plants 10, no. 1: 99. https://doi.org/10.3390/plants10010099
APA StyleRusinque, L., Nóbrega, F., Cordeiro, L., Serra, C., & Inácio, M. L. (2021). First Detection of Meloidogyne luci (Nematoda: Meloidogynidae) Parasitizing Potato in the Azores, Portugal. Plants, 10(1), 99. https://doi.org/10.3390/plants10010099