
Metamitron, a Photosynthetic Electron Transport Chain Inhibitor, Modulates the Photoprotective Mechanism of Apple Trees

 ,
, 
Abstract
:1. Introduction
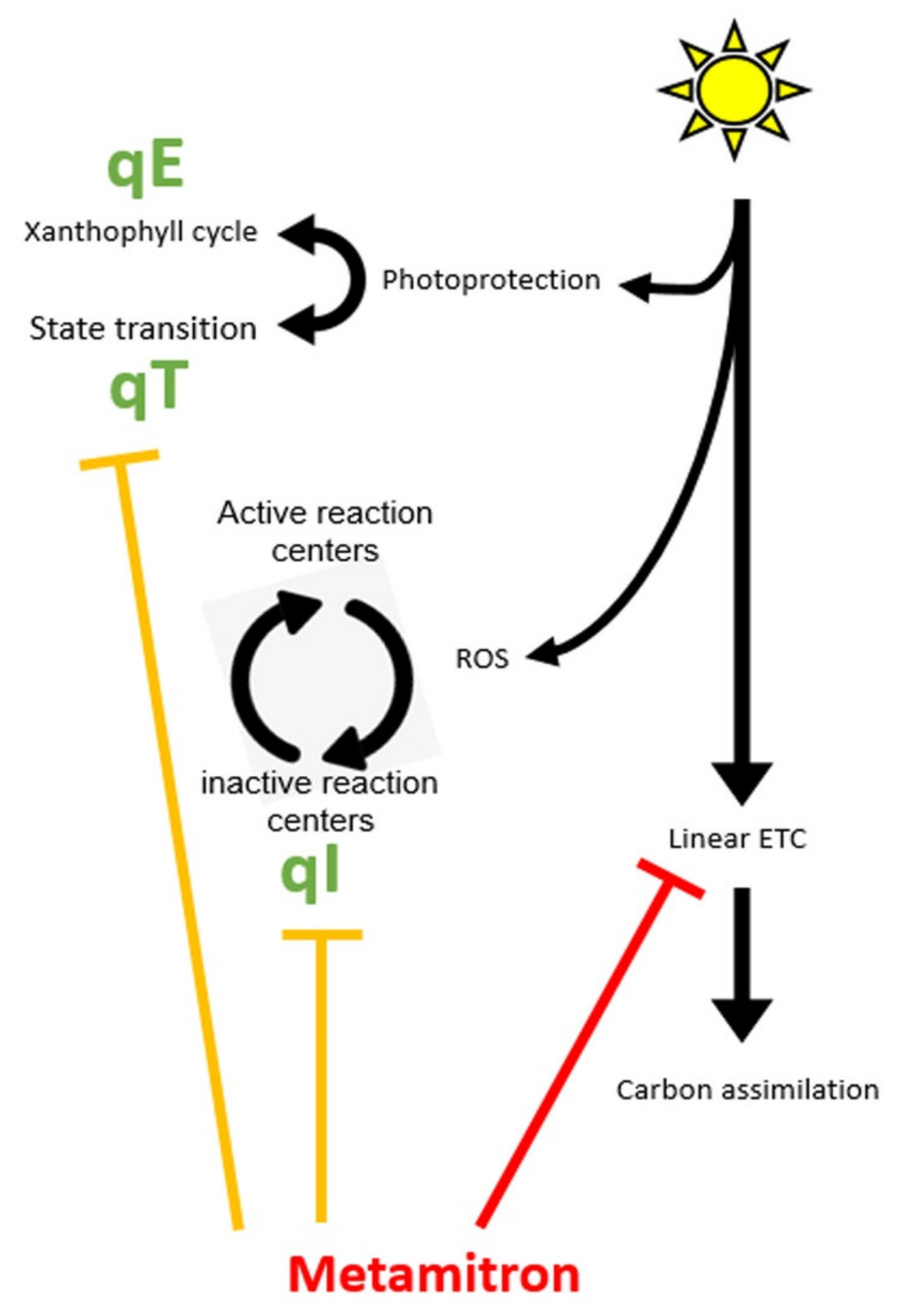
- Energy related (qE, several seconds) [10], responsible for the epoxidation of special carotenoids (Xanthophylls) located on the Light Harvesting Complexes (LHC, photosynthetic antenna). Upon epoxidation, these Xanthophylls change their chemical activity from light harvesting to energy dissipation and thus prevent light energy from reaching the reaction centers of PhotoSystem II (PSII).
- State transition related (qT, minutes) [11], responsible for transition of the LHCII between the two central photosynthetic complexes—PSII and Photosystem I (PSI). This results in a different allocation of absorbed energy between the two photosystems and reduces the damage to PSII.
- Photoinhibition dependent (qI, hours) [12], with prolonged stress, ROS will eventually accumulate in the vicinity of PSII and destroy the D1 subunit an important component of the reaction center [13]. The destruction of D1 results in a non-active PSII. The damaged D1 are marked for replacement via a regulated process that replaces the damaged unit with a de novo synthesized copy [14].
2. Results
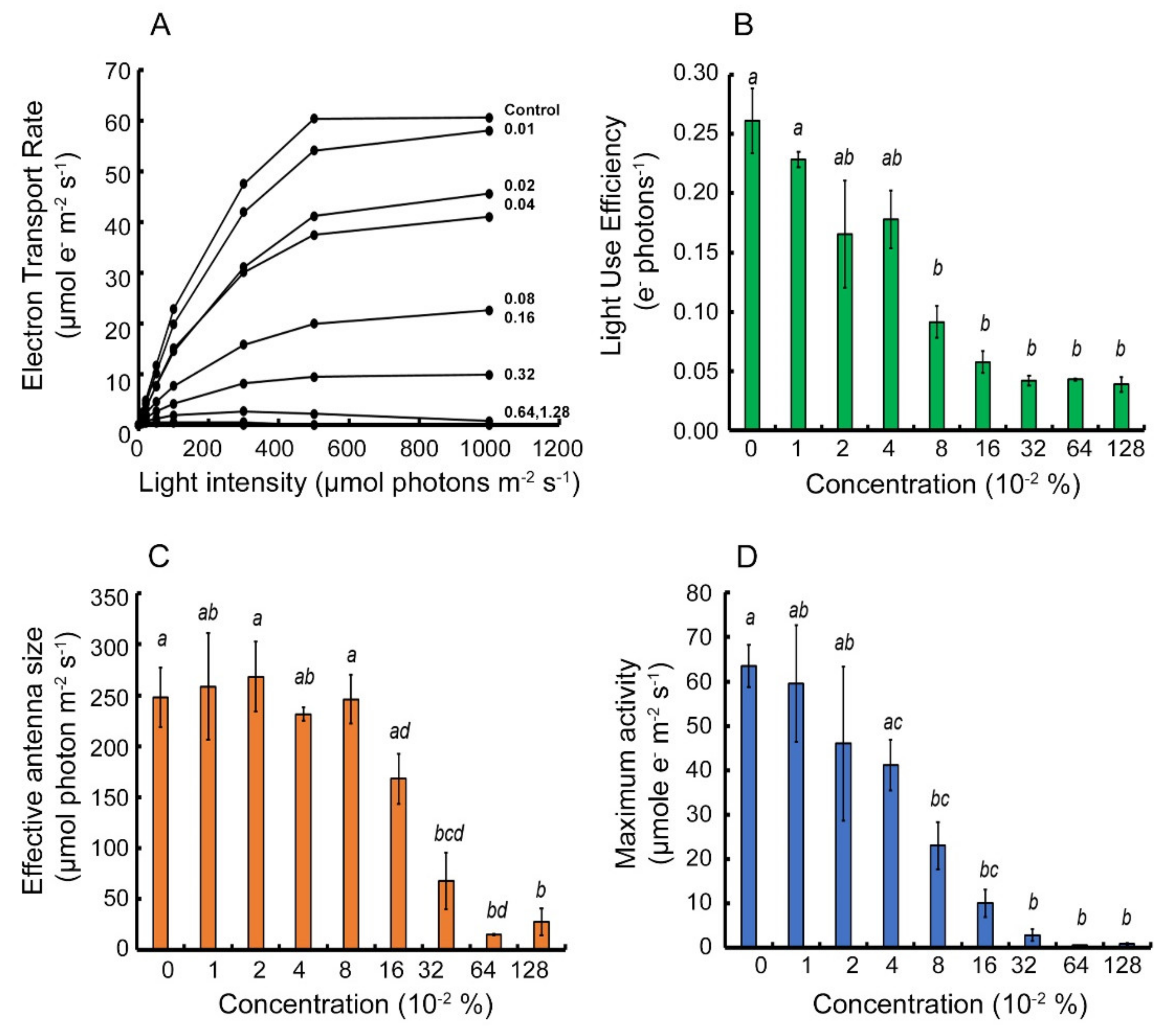
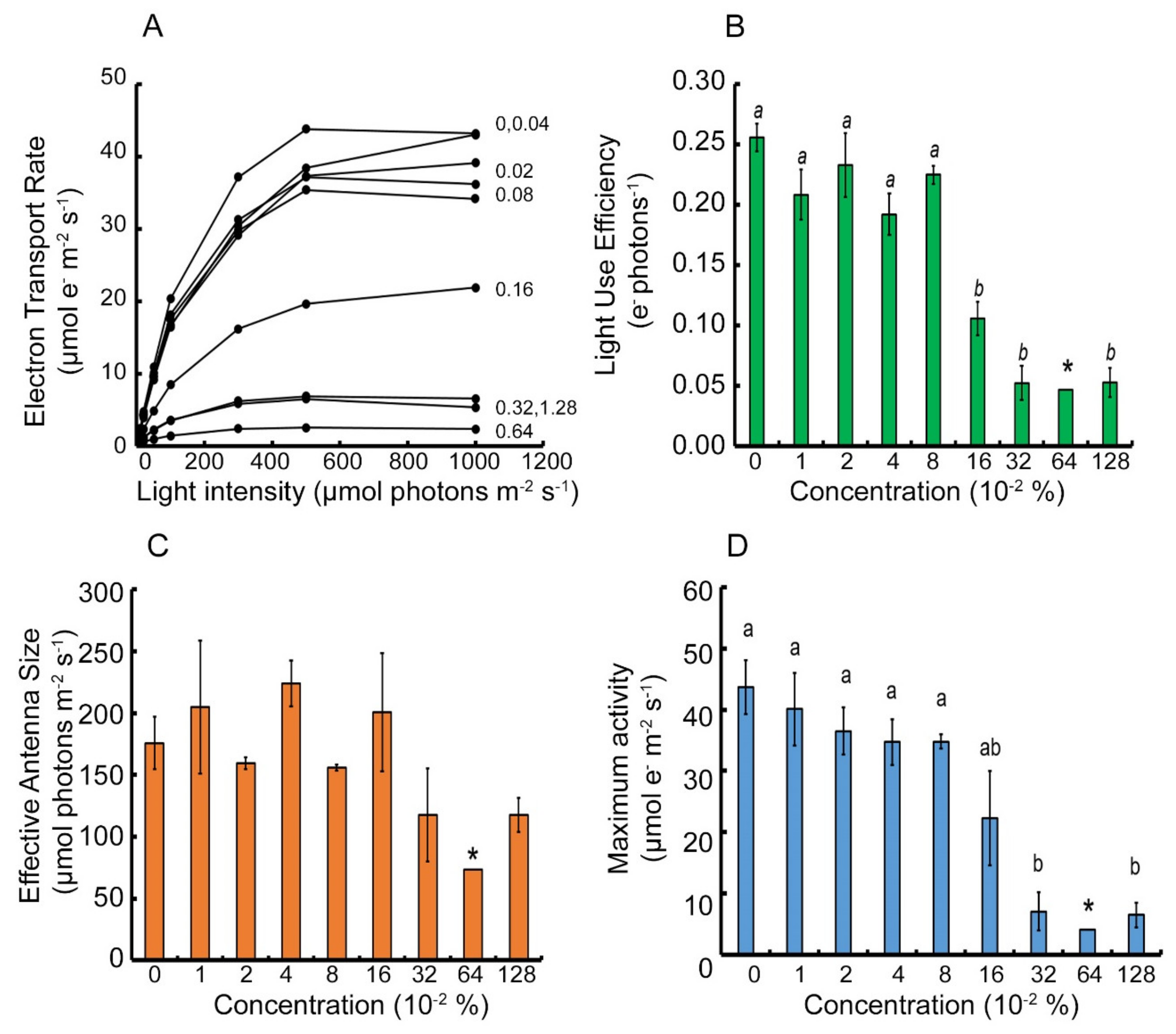
2.1. Parameterization of Light Response Curves of Two Cultivars of Apple Trees Leaves
2.2. Analysis of the Photoprotective Mechanism’s Response to the Metamitron Inhibitor
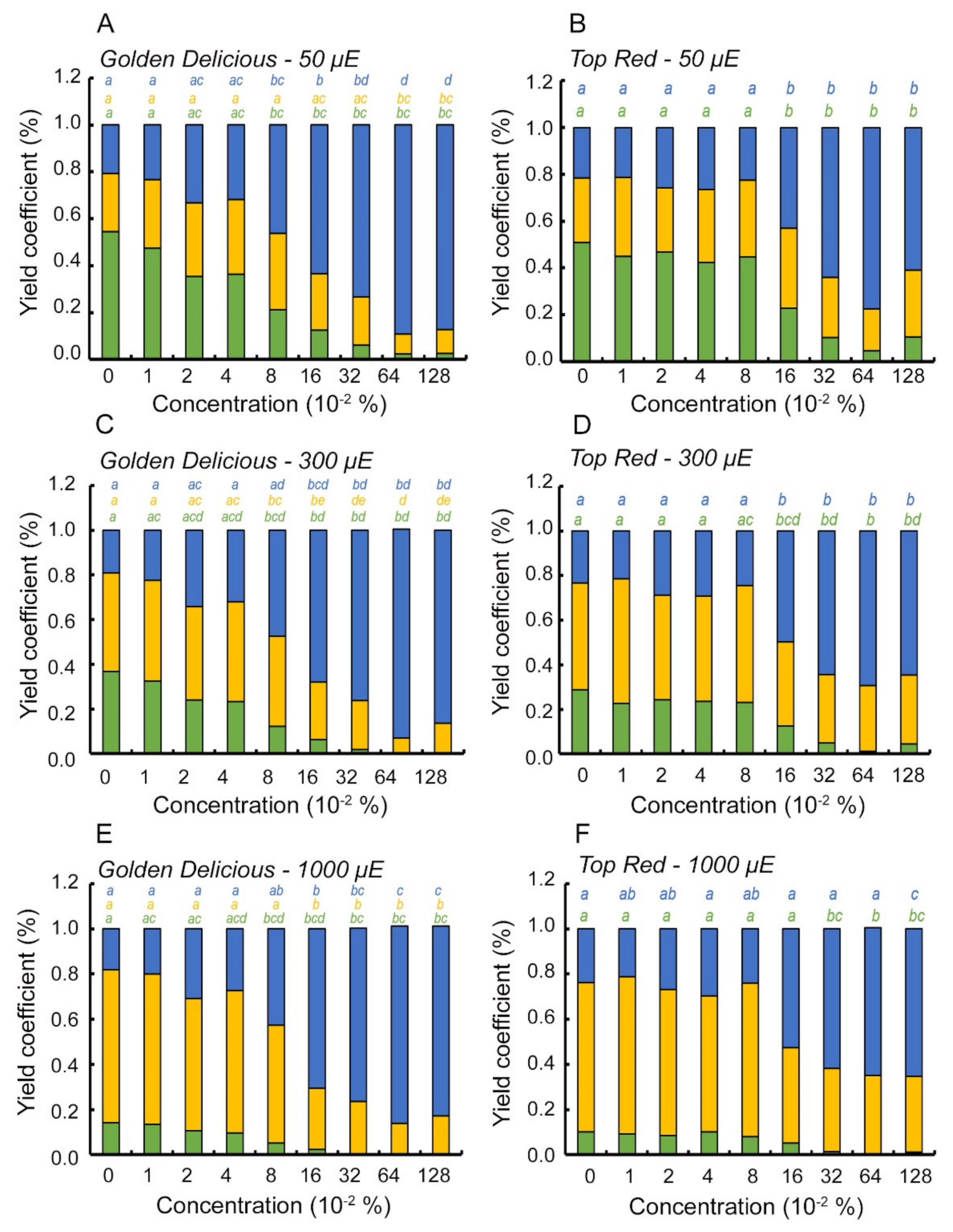
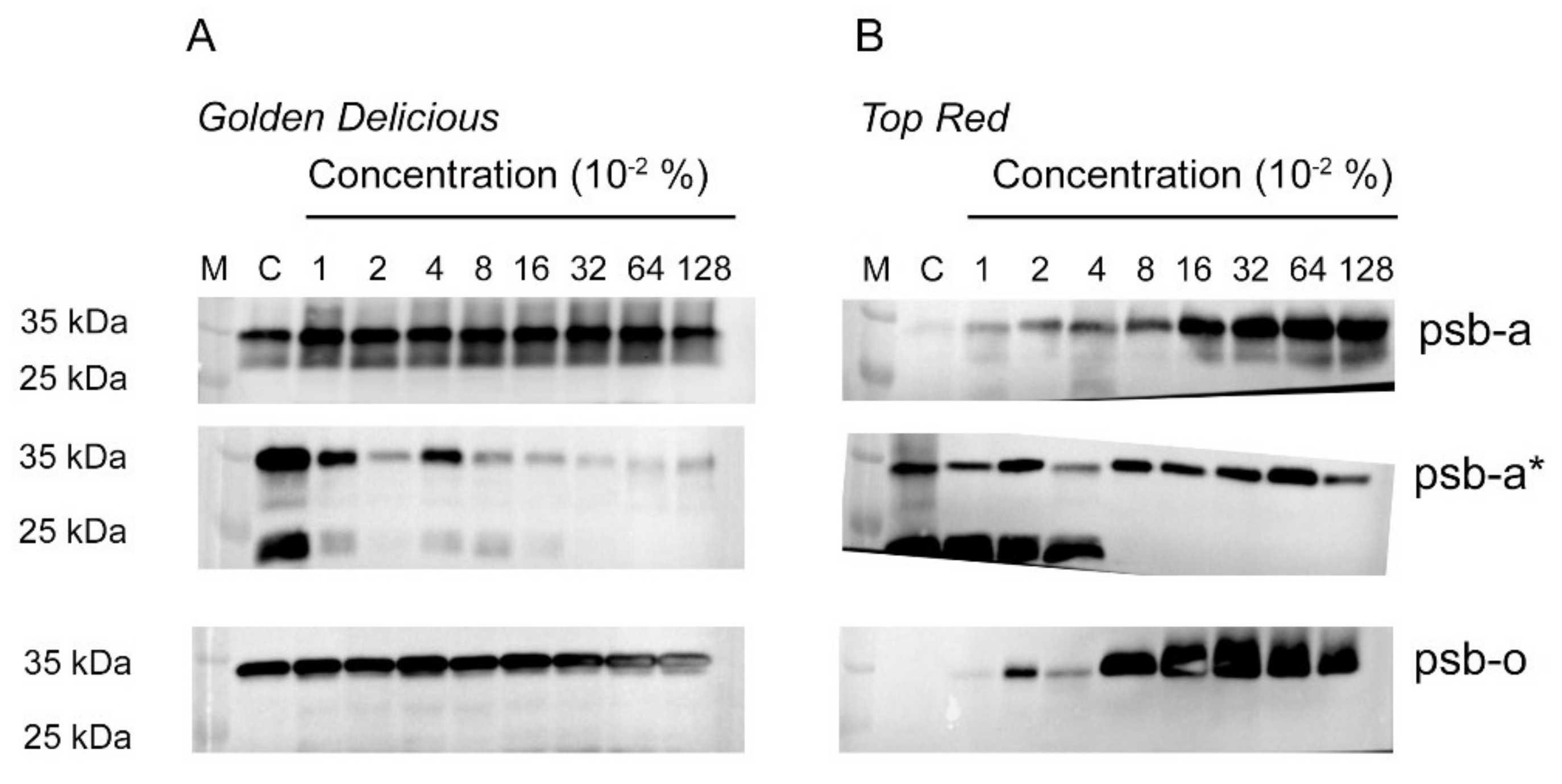
2.3. Metamitron Interferes with the Photoprotective Mechanism in Apple Trees
3. Discussion
4. Materials and Methods
4.1. Plant Material & Chemicals Application
4.2. Chlorophyll a Fluorescence Measurements
- OJIP [28] (Supplementary Figure S1): The OJIP transient is a fast recording of the initial increase in fluorescence upon illumination of dark-adapted leaves. Leaves were adapted to the dark for 20 min. The OJIP initials stand for: O (F0) measured 50 µs after illumination starts, J—first intermediate step measured after 2 ms, I—second intermediate step measured after 30 ms, and P—Fm (maximum fluorescence) [42];
- Light response curve [23,43]: This protocol records maximum quantum yield in the dark and then effective quantum yield in series, each after an illumination period. In such a way, by gradually increasing the light, information regarding light use efficiency, characteristic light intensity and maximum activity of PSII can be extracted from the data. Maximum quantum yield is calculated as:where Fm and F0 are maximum and minimum fluorescence measured after dark adaptation time, respectively. Effective quantum yield is calculated in the same manner as equation (1), where it is considered effective because it is recorded during or at the end of a light period just before closing or changing the light intensity. LC3 pre-configured measurement was used with 7 steps increase in actinic light intensity within the PAM-fluorometer: 10, 20, 50, 100, 300, 500, 1000 µmol photons m−2 s−1. Parameterization of the light response curve followed Eilers and Peeters (1988) formulation [43]: A linear regression curve was fitted to the logarithmic data with the following fit equation:where p and I are the photosynthetic activity and light intensity, respectively. The coefficients a, b and c are used to minimize the fit over the logarithmic data in order to extract the required information. Their initial values at the start of the fitting procedure are calculated directly from the acquired data:where s, Im and Pm are the initial linear slope, the light intensity at maximum photosynthetic activity, and the maximum photosynthetic activity reached. Then, after the fitting procedure is finished, reverse equations are used to extract the information needed:
- Light energy utilization distribution was calculated as suggested by Kramer et al. (2004) [29] where the puddle model [44] was taken into account during calculation:where ΦII is the effective quantum yield of PSII, F′m and F′s are the maximum and steady state fluorescence measured during light, respectively; NPQ initials stand for Non-Photochemical Quenching, ΦNO represents the quantum yield of non-regulated loss in PSII, and ΦNPQ is the quantum yield of regulated non-photochemical energy loss in PSII.
- Relaxation kinetics [45] were calculated during an induction-relaxation pre-configured measurement (NPQ1) in the portable fluorometer. The induction period was given at 60 s at an intensity of 50 µmol photons m−2 s−1 (comparable to the ambient light intensity the leaves experienced during acclimation in the growth chamber). Then, the relaxation period after illumination in the dark was given at 88 s, during which three saturating pulses are fired at a pre-set intervals of 26 s each in the dark. The calculations for each of the coefficients are:where F′#m is the number of the saturating pulse during the relaxation part of the induction- relaxation protocol.
4.3. The 77K Fluorescence Assay
4.4. SDS-PAGE and Western Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dennis, F.G., Jr. The History of Fruit Thinning. Plant Growth Regul. 2000, 31, 1–16. [Google Scholar] [CrossRef]
- Greene, D.; Costa, G. Fruit Thinning in Pome- and Stone-Fruit: State of the Art. Acta Hortic. 2013, 998, 93–102. [Google Scholar] [CrossRef]
- Lakso, A.N.; Johnson, R.S. A Simplified Dry Matter Production Model for Apple Using Automatic Programming Simulation Software. Acta Hortic. 1990, 276, 141–148. [Google Scholar] [CrossRef]
- Rosa, N.; Lidon, F.C.; Rodrigues, A.P.; Pais, I.P.; Scotti-Campos, P.; Asín, L.; Oliveira, C.M.; Ramalho, J.C. Implications of Nighttime Temperature on Metamitron Impacts on the Photosynthetic Machinery Functioning of Malus x Domestica Borkh. J. Plant Physiol. 2021, 261, 153427. [Google Scholar] [CrossRef]
- Stern, R.A. The Photosynthesis Inhibitor Metamitron Is a Highly Effective Thinner for ‘Golden Delicious’ Apple in a Warm Climate. Fruits 2015, 70, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-Photochemical Quenching. A Response to Excess Light Energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Pinnola, A.; Bassi, R. Molecular Mechanisms Involved in Plant Photoprotection. Biochem. Soc. Trans. 2018, 46, 467–482. [Google Scholar] [CrossRef]
- Messant, M.; Krieger-Liszkay, A.; Shimakawa, G. Dynamic Changes in Protein-Membrane Association for Regulating Photosynthetic Electron Transport. Cells 2021, 10, 1216. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schansker, G.; Tóth, S.Z.; Strasser, R.J. Dark Recovery of the Chl a Fluorescence Transient (OJIP) after Light Adaptation: The QT-Component of Non-Photochemical Quenching Is Related to an Activated Photosystem I Acceptor Side. Biochim. Biophys. Acta (BBA)-Bioenerget. 2006, 1757, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H. Reactive Oxygen Species, Oxidative Signaling and the Regulation of Photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Aro, E.-M.; McCaffery, S.; Anderson, J.M. Photoinhibition and D1 Protein Degradation in Peas Acclimated to Different Growth Irradiances. Plant Physiol. 1993, 103, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Melis, A. Photosystem-II Damage and Repair Cycle in Chloroplasts: What Modulates the Rate of Photodamage in Vivo? Trends Plant Sci. 1999, 4, 130–135. [Google Scholar] [CrossRef]
- Rochaix, J.-D.; Lemeille, S.; Shapiguzov, A.; Samol, I.; Fucile, G.; Willig, A.; Goldschmidt-Clermont, M. Protein Kinases and Phosphatases Involved in the Acclimation of the Photosynthetic Apparatus to a Changing Light Environment. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3466–3474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonardi, V.; Pesaresi, P.; Becker, T.; Schleiff, E.; Wagner, R.; Pfannschmidt, T.; Jahns, P.; Leister, D. Photosystem II Core Phosphorylation and Photosynthetic Acclimation Require Two Different Protein Kinases. Nature 2005, 437, 1179–1182. [Google Scholar] [CrossRef]
- Bellafiore, S.; Barneche, F.; Peltier, G.; Rochaix, J.-D. State Transitions and Light Adaptation Require Chloroplast Thylakoid Protein Kinase STN7. Nature 2005, 433, 892–895. [Google Scholar] [CrossRef]
- Theis, J.; Schroda, M. Revisiting the Photosystem II Repair Cycle. Plant Signal Behav. 2016, 11, e1218587. [Google Scholar] [CrossRef] [Green Version]
- Mattoo, A.K.; Pick, U.; Hoffman-Falk, H.; Edelman, M. The Rapidly Metabolized 32,000-Dalton Polypeptide of the Chloroplast Is the “Proteinaceous Shield” Regulating Photosystem II Electron Transport and Mediating Diuron Herbicide Sensitivity. Proc. Natl. Acad. Sci. USA 1981, 78, 1572–1576. [Google Scholar] [CrossRef] [Green Version]
- Allakhverdiev, S.I.; Nishiyama, Y.; Takahashi, S.; Miyairi, S.; Suzuki, I.; Murata, N. Systematic Analysis of the Relation of Electron Transport and ATP Synthesis to the Photodamage and Repair of Photosystem II in Synechocystis. Plant Physiol. 2005, 137, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous Recording of Photochemical and Non-Photochemical Chlorophyll Fluorescence Quenching with a New Type of Modulation Fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The Relationship between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Henley, W.J. Measurement and Interpretation of Photosynthetic Light-Response Curves in Algae in the Context of Photoinhibition and Diel Changes. J. Phycol. 1993, 29, 729–739. [Google Scholar] [CrossRef]
- Long, S.P.; Humphries, S.; Falkowski, P.G. Photoinhibition of Photosynthesis in Nature. Annu. Rev. Plant Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Kroon, B.; Prézelin, B.B.; Schofield, O. Chormatic Regulation of Quantum Yields for Photosystem II Charge Separation, Oxygen Evolution, and Carbon Fixation in Heterocapsa Pygmaea (Pyrrophyta). J. Phycol. 1993, 29, 453–462. [Google Scholar] [CrossRef]
- Jassby, A.D.; Platt, T. Mathematical Formulation of the Relationship between Photosynthesis and Light for Phytoplankton. Limnol. Oceanogr. 1976, 21, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence; Springer: Berlin, Germany, 2004; pp. 321–362. [Google Scholar]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of QA Redox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209. [Google Scholar] [CrossRef]
- Henmi, T.; Miyao, M.; Yamamoto, Y. Release and Reactive-Oxygen-Mediated Damage of the Oxygen-Evolving Complex Subunits of PSII during Photoinhibition. Plant Cell Physiol. 2004, 45, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Klughammer, C.; Schreiber, U. Complementary PS II Quantum Yields Calculated from Simple Fluorescence Parameters Measured by PAM Fluorometry and the Saturation Pulse Method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- Wen, X.; Yang, Z.; Ding, S.; Yang, H.; Zhang, L.; Lu, C.; Lu, Q. Analysis of the Changes of Electron Transfer and Heterogeneity of Photosystem II in Deg1-Reduced Arabidopsis Plants. Photosynth. Res. 2021, 150, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Greer, D.H. Photon Flux Density and Temperature-Dependent Responses of Photosynthesis and Photosystem II Performance of Apple Leaves Grown in Field Conditions. Funct. Plant Biol. 2015, 42, 782–791. [Google Scholar] [CrossRef]
- Campbell, R.J.; Marini, R.P.; Birch, J.B. Canopy Position Affects Light Response Curves for Gas Exchange Characteristics of Apple Spur Leaves. J. Am. Soc. Hortic. Sci. 1992, 117, 467–472. [Google Scholar] [CrossRef] [Green Version]
- McArtney, S.J.; Obermiller, J.D.; Arellano, C. Comparison of the Effects of Metamitron on Chlorophyll Fluorescence and Fruit Set in Apple and Peach. HortScience 2012, 47, 6. [Google Scholar] [CrossRef] [Green Version]
- Pfündel, E.; Klughammer, C.; Schreiber, U. Monitoring the Effects of Reduced PS II Antenna Size on Quantum Yields of Photosystems I and II Using the Dual-PAM-100 Measuring System. PAM Appl. Notes 2008, 1, 21–24. [Google Scholar]
- Robinson, T.L.; Lakso, A.N. Advances in Predicting Chemical Thinner Response of Apple Using a Carbon Balance Model. N. Y. Fruit Q. 2011, 19, 15–20. [Google Scholar] [CrossRef]
- Lakso, A.N.; Robinson, T.L. Decision Support for Apple Thinning Based on Carbon Balance Modeling. In Proceedings of the IX International Symposium on Modelling in Fruit Research and Orchard Management 1068, Saint-Jean-sur-Richelieu, QC, Canada, 19–23 June 2011; pp. 235–242. [Google Scholar]
- Stander, O.P.J.; Botes, J.; Krogscheepers, C. The Potential Use of Metamitron as a Chemical Fruit-Thinning Agent in Mandarin. HortTechnology 2018, 28, 28–34. [Google Scholar] [CrossRef] [Green Version]
- De Mello Farias, R.; Barreto, C.F.; Schiavon, A.V.; Hellwig, C.G.; Martins, C.R.; Malgarim, M.B.; Mello-Farias, P.C. Effect of Chemical Thinning Season Using Metamitron on Peaches ‘Sensação’Quality. J. Exp. Agric. Int. 2017, 19, 1–7. [Google Scholar]
- Elsysy, M.A.; Hubbard, A.; Einhorn, T.C. Postbloom Thinning of ‘Bartlett’Pear with Metamitron. HortScience 2020, 55, 174–180. [Google Scholar] [CrossRef]
- Ariyarathna, R.A.I.S.; Weerasena, S.L.; Beneragama, C. Application of Polyphasic OJIP Chlorophyll Fluorescent Transient Analysis as an Indicator for Testing of Seedling Vigour of Common Bean (Phaseolus Vulgaris L.). Trop. Agric. Res. 2020, 31, 106. [Google Scholar] [CrossRef]
- Eilers, P.H.C.; Peeters, J.C.H. A Model for the Relationship between Light Intensity and the Rate of Photosynthesis in Phytoplankton. Ecol. Model. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Lavergne, J.; Trissl, H.-W. Theory of Fluorescence Induction in Photosystem II: Derivation of Analytical Expressions in a Model Including Exciton-Radical-Pair Equilibrium and Restricted Energy Transfer between Photosynthetic Units. Biophys. J. 1995, 68, 2474–2492. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to Correctly Determine the Different Chlorophyll Fluorescence Parameters and the Chlorophyll Fluorescence Decrease Ratio R Fd of Leaves with the PAM Fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Liran, O.; Shir, O.M.; Levy, S.; Grunfeld, A.; Shelly, Y. Novel Remote Sensing Index of Electron Transport Rate Predicts Primary Production and Crop Health in L. Sativa and Z. Mays. Remote Sens. 2020, 12, 1718. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days | Concentration (%) | Quantum Yield of Process ‡ | ||
|---|---|---|---|---|
| qE | qT | qI | ||
| 2 | 0 | 0.156 ± 0.090 ab | 0.096 ± 0.012 a | 0.536 ± 0.120 ab |
| 0.01 | 0.143 ± 0.032 a | 0.136 ± 0.011 ab | 0.473 ± 0.049 a | |
| 0.05 | 0.336 ± 0.016 b | 0.166 ± 0.005 b | 0.257 ± 0.023 b | |
| 0.5 | 0.227 ± 0.027 ab | 0.154 ± 0.011 b | 0.411± 0.039 ab | |
| 5 | 0 | 0.278 ± 0.081 | 0.149 ± 0.045 | 0.520 ± 0.186 |
| 0.01 | 0.134 ± 0.076 | 0.105 ± 0.023 | 0.527 ± 0.132 | |
| 0.05 | 0.223 ± 0.022 | 0.171 ± 0.024 | 0.439 ± 0.076 | |
| 0.5 | 0.255 ± 0.045 | 0.189 ± 0.010 | 0.389 ± 0.068 | |
| 7 | 0 | 0.114 ± 0.026 | 0.132 ± 0.027 | 0.710 ± 0.245 |
| 0.01 | 0.063 ± 0.033 | 0.094 ± 0.034 | 0.521 ± 0.080 | |
| 0.05 | 0.168 ± 0.046 | 0.176 ± 0.033 | 0.524 ± 0.093 | |
| 0.5 | 0.152 ± 0.028 | 0.176 ± 0.014 | 0.498 ± 0.008 | |
| 10 | 0 | 0.087 ± 0.012 | 0.129 ± 0.017 | 0.471 ± 0.030 |
| 0.01 | 0.121 ± 0.020 | 0.127 ± 0.024 | 0.484 ± 0.038 | |
| 0.05 | 0.161 ± 0.020 | 0.173 ± 0.007 | 0.439 ± 0.022 | |
| 0.5 | 0.166 ± 0.033 | 0.170 ± 0.08 | 0.466 ± 0.042 | |
| Days | Concentration (%) | Quantum Yield of Process ‡ | ||
|---|---|---|---|---|
| qE | qT | qI | ||
| 2 | 0 | 0.165 ± 0.003 | 0.140 ± 0.009 | 0.465 ± 0.028 a |
| 0.01 | 0.194 ± 0.030 | 0.135 ± 0.014 | 0.430 ± 0.048 a | |
| 0.05 | 0.198 ± 0.022 | 0.138 ± 0.004 | 0.437 ± 0.028 a | |
| 0.5 | 0.241 ± 0.061 | 0.099 ± 0.048 | 0.285 ± 0.069 b | |
| 5 | 0 | 0.149 ± 0.006 | 0.148 ± 0.017 a | 0.471 ± 0.044 a |
| 0.01 | 0.123 ± 0.006 | 0.133 ± 0.004 a | 0.500 ± 0.002 a | |
| 0.05 | 0.143 ± 0.015 | 0.130 ± 0.012 a | 0.535 ± 0.036 a | |
| 0.5 | 0.199 ± 0.060 | 0.067 ± 0.020 b | 0.243 ± 0.015 b | |
| 7 | 0 | 0.145 ± 0.018 | 0.154 ± 0.014 | 0.455 ± 0.032 |
| 0.01 | 0.116 ± 0.040 | 0.122 ± 0.017 | 0.528 ± 0.073 | |
| 0.05 | 0.151 ± 0.025 | 0.160 ± 0.022 | 0.439 ± 0.060 | |
| 0.5 | 0.263 ± 0.087 | 0.133 ± 0.042 | 0.509 ± 0.133 | |
| 10 | 0 | 0.040 ± 0.018 | 0.059 ± 0.019 | 0.534 ± 0.029 a |
| 0.01 | 0.062 ± 0.018 | 0.089 ± 0.008 | 0.581 ± 0.005 a | |
| 0.05 | 0.045 ± 0.041 | 0.078 ± 0.033 | 0.626 ± 0.038 a | |
| 0.5 | 0.039 ± 0.022 | 0.033 ± 0.011 | 1.046 ± 0.121 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tadmor, Y.; Raz, A.; Reikin-Barak, S.; Ambastha, V.; Shemesh, E.; Leshem, Y.; Crane, O.; Stern, R.A.; Goldway, M.; Tchernov, D.; et al. Metamitron, a Photosynthetic Electron Transport Chain Inhibitor, Modulates the Photoprotective Mechanism of Apple Trees. Plants 2021, 10, 2803. https://doi.org/10.3390/plants10122803
Tadmor Y, Raz A, Reikin-Barak S, Ambastha V, Shemesh E, Leshem Y, Crane O, Stern RA, Goldway M, Tchernov D, et al. Metamitron, a Photosynthetic Electron Transport Chain Inhibitor, Modulates the Photoprotective Mechanism of Apple Trees. Plants. 2021; 10(12):2803. https://doi.org/10.3390/plants10122803
Chicago/Turabian StyleTadmor, Yuval, Amir Raz, Shira Reikin-Barak, Vivek Ambastha, Eli Shemesh, Yehoram Leshem, Omer Crane, Raphael A. Stern, Martin Goldway, Dan Tchernov, and et al. 2021. "Metamitron, a Photosynthetic Electron Transport Chain Inhibitor, Modulates the Photoprotective Mechanism of Apple Trees" Plants 10, no. 12: 2803. https://doi.org/10.3390/plants10122803
APA StyleTadmor, Y., Raz, A., Reikin-Barak, S., Ambastha, V., Shemesh, E., Leshem, Y., Crane, O., Stern, R. A., Goldway, M., Tchernov, D., & Liran, O. (2021). Metamitron, a Photosynthetic Electron Transport Chain Inhibitor, Modulates the Photoprotective Mechanism of Apple Trees. Plants, 10(12), 2803. https://doi.org/10.3390/plants10122803