
Anti-Inflammatory Effect of Artemisia argyi on Ethanol-Induced Gastric Ulcer: Analytical, In Vitro and In Vivo Studies for the Identification of Action Mechanism and Active Compounds

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extraction
2.3. HPLC-UV/DAD Conditions
2.4. Cell Culture
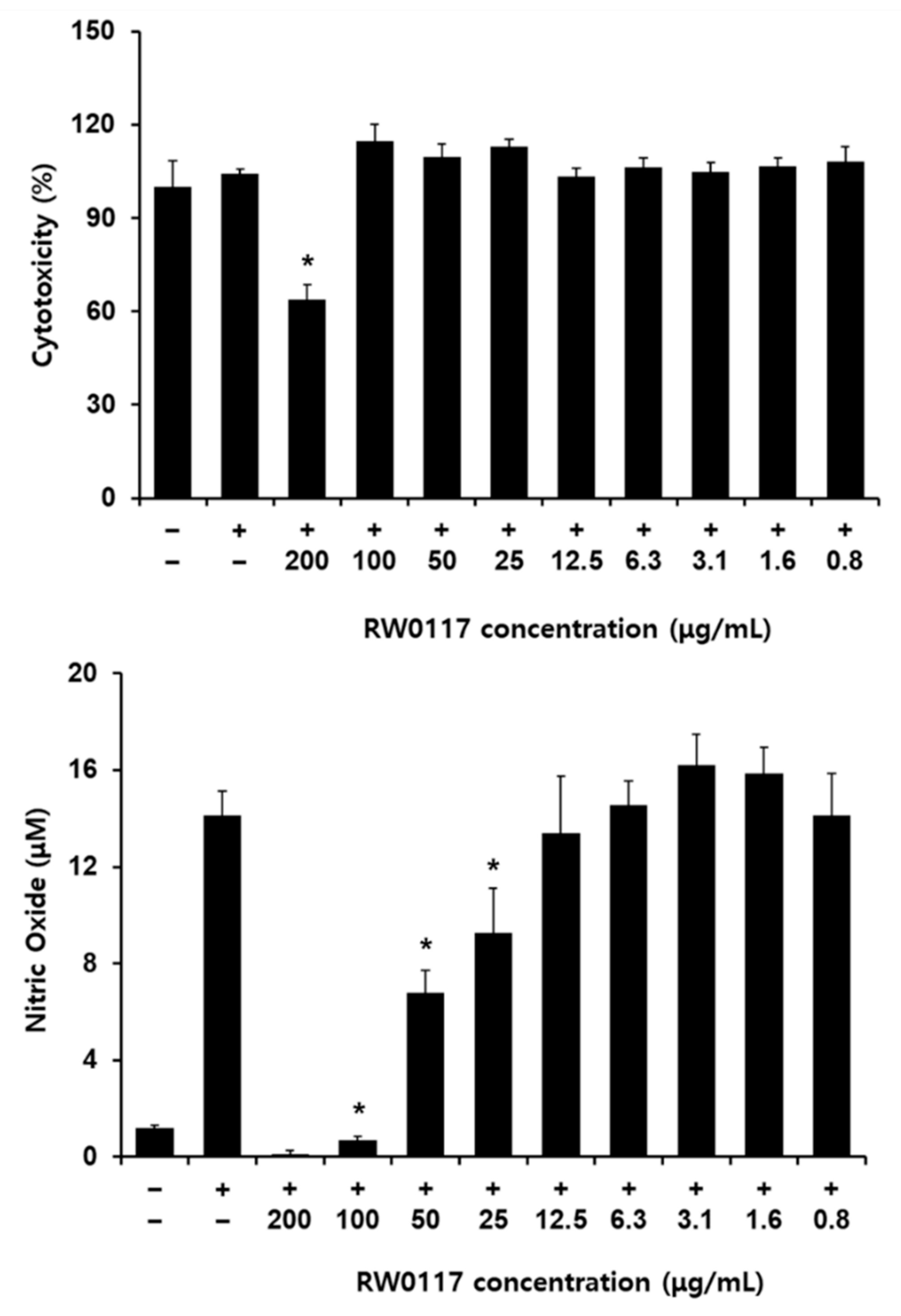
2.5. Measurement of RW0117 Cytotoxicity on RAW264.7 Cells
2.6. Determination of Nitric Oxide, IL-6, and TNF-α Production
2.7. Immunoblotting
2.8. Real-Time Reverse Transcription Polymerase Chain Reaction
2.9. Immunofluorescence Microscopy
2.10. Animals
2.11. Ethanol and HCl-Induced Gastric Ulcer
2.12. Histological Analysis
2.13. Statistical Analysis
3. Results and Discussion
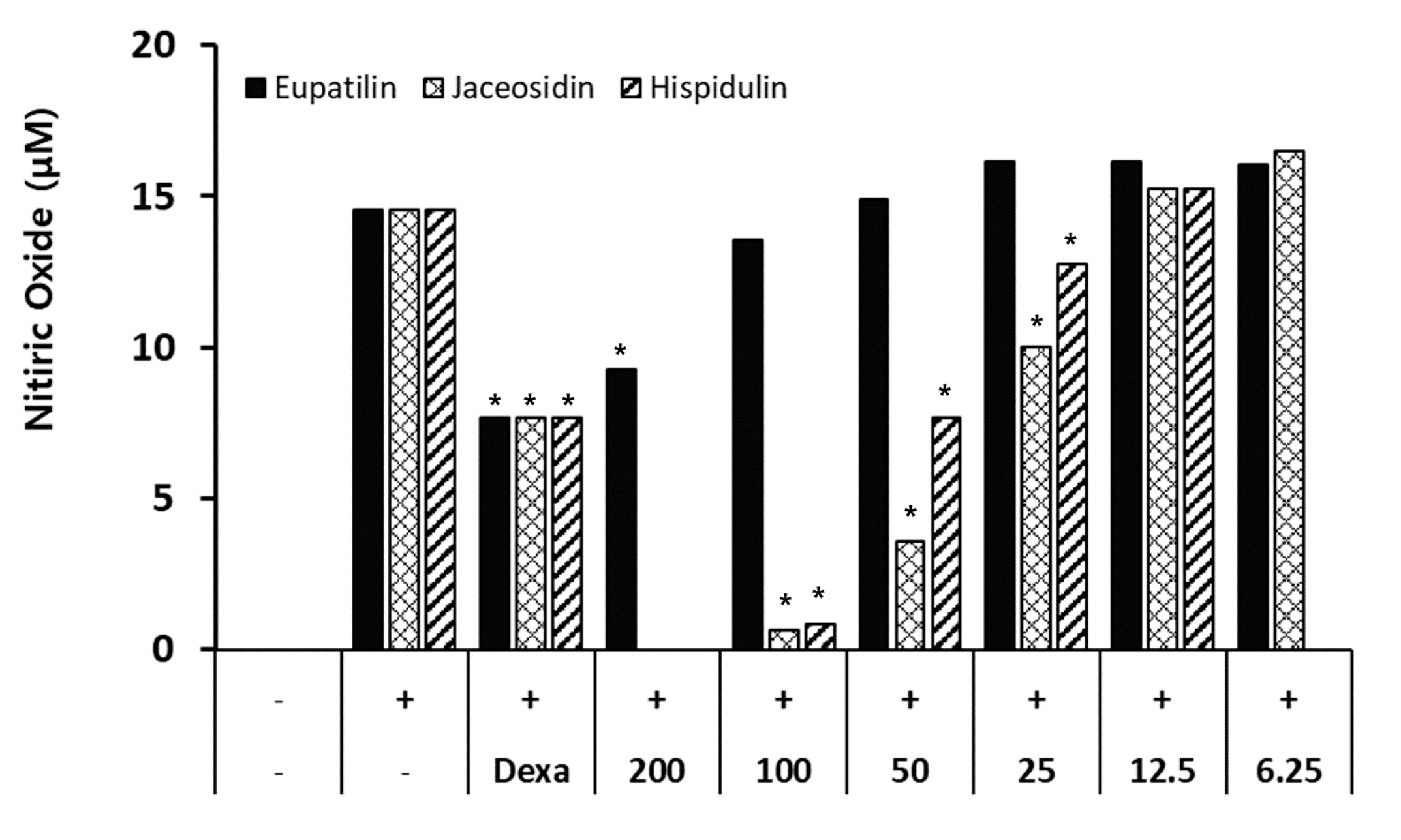
3.1. HPLC Profile of RW0117
3.2. The Effects of RW0117 on Nitric Oxide Production in LPS-Stimulated RAW 264.7 Cells
3.3. Analysis of the Effect of RW0117 on Cytokine Production and mRNA Expression in RAW264.7 Cells
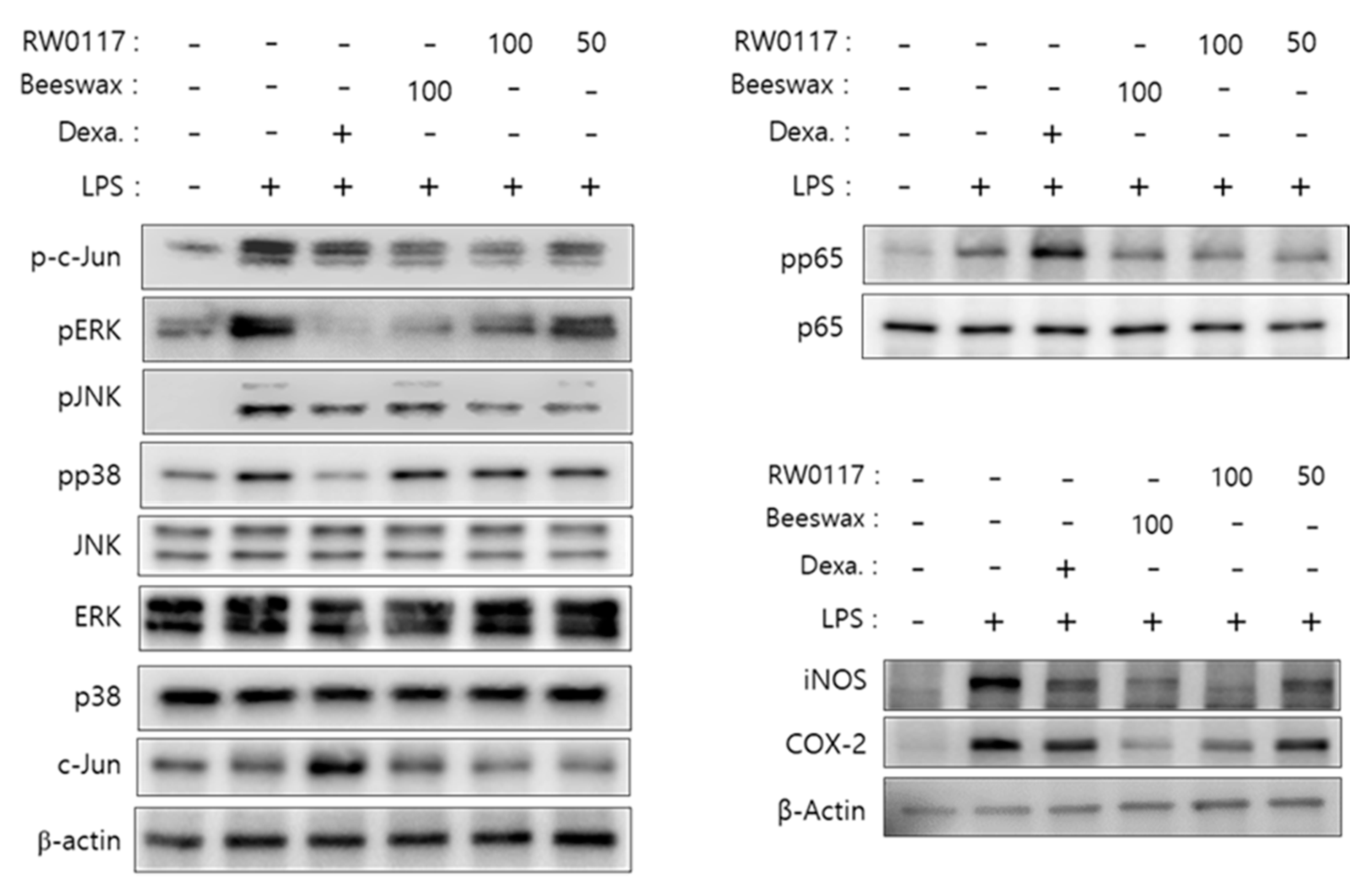
3.4. Analysis of the Effect of RW0117 on MAPK and NF-kB Signaling Pathways
3.5. Analysis of Nuclear Translocation of NF-kB p65 by RW0117 in RAW264.7 Cells
3.6. Protective Effects of RW0117 in Rats with HCl/EtOH-Induced Gastric Ulcers
3.7. Analysis of the Gene Expressions of MUC1, MUC5AC, and MUC6 in Gastric Ulcer in Rats
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zedler, S.; Faist, E. The impact of endogenous triggers on trauma-associated inflammation. Curr. Opin. Crit. Care 2006, 12, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Isomäki, P.; Punnonen, J. Pro- and anti-inflammatory cytokines in rheumatoid arthritis. Ann. Med. 1997, 29, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Mitchell, J.A.; Appleton, I.; Tomlinson, A.; Bishop-Bailey, D.; Croxtall, J.; Willoughby, D.A. Inducible isoforms of cyclooxygenase and nitric-oxide synthase in inflammation. Proc. Natl. Acad. Sci. USA 1994, 91, 2046–2050. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, M.; Higashi, N.; Taki, H.; Osawa, T. Cytolytic mechanisms of activated macrophages. Tumor necrosis factor and Larginine-dependent mechanisms act synergistically as the major cytolytic mechanisms of activated macrophages. J. Immunol. 1990, 144, 1425–1431. [Google Scholar] [PubMed]
- Nathan, C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992, 6, 3051–3064. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar]
- Franke, A.; Teyssen, S.; Singer, M.V. Alcohol-related diseases of the esophagus and stomach. Dig. Dis. 2005, 23, 204–213. [Google Scholar] [CrossRef]
- Marshal, B.J.; Waren, J.R. Unidentified curved baciliin the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 131–1315. [Google Scholar]
- Honda, S.; Fujioka, T.; Tokieda, M.; Satoh, R.; Nishizono, A.; Nasu, M. Development of Helicobacter pylori-induced gastric carcinoma in Mongolian gerbils. Cancer Res. 1998, 58, 425–4259. [Google Scholar]
- Goel, R.K.; Bhattacharya, S.K. Gastroduodenal mucosal defence and mucosal protective agents. Indian J. Exp. Biol. 1991, 29, 701–714. [Google Scholar]
- DeVault, K.R.; Talley, N.J. Insights into the future of gastric acid suppression. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Xu, H.D.; Zhao, B.T.; Chang, H.I.; Rhee, H.I. Gastroprotective effect of cyanidin 3-glucoside on ethanol-induced gastric lesions in rats. Alcohol 2008, 42, 683–687. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, L.; Wang, J.J.; Luo, P.F.; Wang, X.T.; Xia, Z.F. The caspase-1 inhibitor AC-YVAD-CMK attenuates acute gastric injury in mice: Involvement of silencing NLRP3 inflammasome activities. Sci. Rep. 2016, 6, 24166. [Google Scholar] [CrossRef]
- Li, W.; Huang, H.; Niu, X.; Fan, T.; Mu, Q.; Li, H. Protective effect of tetrahydrocoptisine against ethanol-induced gastric ulcer in mice. Toxicol. Appl. Pharmacol. 2013, 272, 21–29. [Google Scholar] [CrossRef]
- Vinay, S.C.; Pushpesh, K.M.; Rakesh, M.; Dharmani, P.; Gautam, P. Allophylus serratus: A plant with potential anti-ulcerogenic activity. J. Ethnopharmacol. 2005, 99, 361–366. [Google Scholar]
- Sai, K.B.; Radha, K.L.; Gowrinath, R.M. Anti-ulcer effect of Aloe vera in non-steroidal anti-inflammatory drug induced peptic ulcers in rats. Afr. J. Pharm. Pharmacol. 2011, 5, 1867–1871. [Google Scholar]
- Li, Q.; Hu, X.; Xuan, Y.; Ying, J.; Fei, Y.; Rong, J.; Liu, Z. Kaempferol protects ethanol-induced gastric ulcers in mice via pro-inflammatory cytokines and NO. Acta. Biochim. Biophys. Sin. 2018, 50, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.; Jung, Y.; Chun, W.; Yang, B.; Ryu, J.; Lim, C.; Cho, S.I. Anti-inflammatory effects of Artemisia leaf extract in mice with contact dermatitis in vitro and in vivo. Mediat. Inflamm. 2016. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.-R.; Ryu, H.-W.; Ko, J.-W.; Park, S.-H.; Yuk, H.-J.; Kim, H.-J. Artemisia argyi attenuates airway inflammation in ovalbumin-induced asthmatic animals. J. Ethnopharmacol. 2017, 209, 108–115. [Google Scholar] [CrossRef]
- Zimmermann-Klemd, A.M.; Reinhardt, J.K.; Morath, A.; Schamel, W.W.; Steinberger, P.; Leitner, J.; Gründemann, C. Immunosuppressive Activity of Artemisia argyi Extract and Isolated Compounds. Front. Pharmacol. 2020, 11, 402. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Lee, S.J.; Venkatarame Gowda Saralamma, V.; Ha, S.E.; Vetrivel, P.; Desta, K.T.; Kim, G.S. Polyphenol mixture of a native Korean variety of Artemisia argyi H. (Seomae mugwort) and its anti-inflammatory effects. Int. J. Mol. Med. 2019, 44, 1741–1752. [Google Scholar] [CrossRef]
- Choi, E.J.; Lee, S.; Chae, J.R.; Lee, H.S.; Jun, C.D.; Kim, S.H. Eupatilin inhibits lipopolysaccharide-induced expression of inflammatory mediators in macrophages. Life Sci. 2011, 88, 1121–1126. [Google Scholar] [CrossRef]
- Min, S.W.; Kim, N.J.; Baek, N.I.; Kim, D.H. Inhibitory effect of eupatilin and jaceosidin isolated from Artemisia princeps on carrageenan-induced inflammation in mice. J. Ethnopharmacol. 2009, 125, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Min, K.J.; Kim, M.J.; Kim, S.H.; Kwon, T.K. Hispidulin inhibits mast cell-mediated allergic inflammation through down-regulation of histamine release and inflammatory cytokines. Molecules 2019, 24, 2131. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.S.; Song, J.H.; Choi, P.; Lee, J.H.; Kim, S.Y.; Shin, K.S.; Kang, K.S. Stimulation of innate immune function by panax ginseng after heat processing. J. Agric. Food Chem. 2018, 66, 4652–4659. [Google Scholar] [CrossRef]
- Shin, M.S.; Kim, S.B.; Lee, J.; Choi, H.S.; Park, J.; Park, J.Y.; Kang, K.S. Beneficial effect of herbal formulation KM1608 on inflammatory bowel diseases: A preliminary experimental study. Molecules 2018, 23, 2068. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.C.; Kim, J.H. Gastroprotective effects of irsogladine maleate on ethanol/hydrochloric acid induced gastric ulcers in mice. Korean J. Intern. Med. 2021, 36, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, I.; Illnait, J.; Terry, H.; Mas, R.; Fernández, L.; Fernández, J.C.; Cruz, Y. Effects of Abexol (beeswax alcohols) on gastrointestinal symptoms in middle-aged and older subjects. Rev. CENIC Cienc. Biol. 2009, 40, 147–154. [Google Scholar]
- Song, X.; Wen, X.; He, J.; Zhao, H.; Li, S.; Wang, M. Phytochemical components and biological activities of Artemisia Argyi. J. Funct. Foods 2019, 52, 648–662. [Google Scholar] [CrossRef]
- Kim, J.K.; Shin, E.C.; Lim, H.J.; Choi, S.J.; Kim, C.R.; Suh, S.H.; Shin, D.H. Characterization of nutritional composition, antioxidative capacity, and sensory attributes of Seomae mugwort, a native Korean variety of Artemisia argyi H. Lev. & Vaniot. J. Anal. Methods Chem. 2015, 2015, 1–9. [Google Scholar]
- Seo, J.M.; Kang, H.M.; Son, K.H.; Kim, J.H.; Lee, C.W.; Kim, H.M.; Kwon, B.M. Antitumor activity of flavones isolated from Artemisia argyi. Planta Med. 2003, 69, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.L.; Li, Z.Z.; Zhang, L.B. Two new flavonoids from Artemisia argyi with their anticoagulation activities. Nat. Prod. Res. 2018, 32, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Shi, B.; Li, T.; Xu, Y.; Jin, X.; Guo, X.; Yan, S. Immunomodulatory effect of Artemisia argyi polysaccharide on peripheral blood leucocyte of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 939–946. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Mouse IL-6 | GAGGATACCACTCCCAACAGACC | AAGTGCATCATCGTTGTTCATACA |
| Mouse iNOS | ACATCGACCCGTCCACAGTAT | CAGAGGGGTAGGCTTGTCTC |
| Mouse β-tubulin | CTCCCAGGTTAAAGTCCTTC | GCAACATAAATACAGAGGTG |
| Rat MUC1 | Rn01462585_m1 (Muc1) | |
| Rat MUC5AC | Rn01451252_m1 (Muc5ac) | |
| Rat MUC6 | Rn01759814_m1 (Muc6) | |
| Rat GAPDH | Rn01775763_g1 (gapdh) | |
| Active Compound | Content (%) |
|---|---|
| Eupatilin | 0.573 ± 0.002 |
| Jaceosidin | 0.223 ± 0.006 |
| Hispidulin | 0.070 ± 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, M.-S.; Lee, J.; Lee, J.W.; Park, S.H.; Lee, I.K.; Choi, J.A.; Lee, J.S.; Kang, K.S. Anti-Inflammatory Effect of Artemisia argyi on Ethanol-Induced Gastric Ulcer: Analytical, In Vitro and In Vivo Studies for the Identification of Action Mechanism and Active Compounds. Plants 2021, 10, 332. https://doi.org/10.3390/plants10020332
Shin M-S, Lee J, Lee JW, Park SH, Lee IK, Choi JA, Lee JS, Kang KS. Anti-Inflammatory Effect of Artemisia argyi on Ethanol-Induced Gastric Ulcer: Analytical, In Vitro and In Vivo Studies for the Identification of Action Mechanism and Active Compounds. Plants. 2021; 10(2):332. https://doi.org/10.3390/plants10020332
Chicago/Turabian StyleShin, Myoung-Sook, Jaemin Lee, Jin Woo Lee, Se Hoon Park, Il Kyun Lee, Jung A. Choi, Jung Suk Lee, and Ki Sung Kang. 2021. "Anti-Inflammatory Effect of Artemisia argyi on Ethanol-Induced Gastric Ulcer: Analytical, In Vitro and In Vivo Studies for the Identification of Action Mechanism and Active Compounds" Plants 10, no. 2: 332. https://doi.org/10.3390/plants10020332
APA StyleShin, M. -S., Lee, J., Lee, J. W., Park, S. H., Lee, I. K., Choi, J. A., Lee, J. S., & Kang, K. S. (2021). Anti-Inflammatory Effect of Artemisia argyi on Ethanol-Induced Gastric Ulcer: Analytical, In Vitro and In Vivo Studies for the Identification of Action Mechanism and Active Compounds. Plants, 10(2), 332. https://doi.org/10.3390/plants10020332