
Sub-Tissue Localization of Phytochemicals in Cinnamomum camphora (L.) J. Presl. Growing in Northern Italy

 , ,
, ,  ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Results
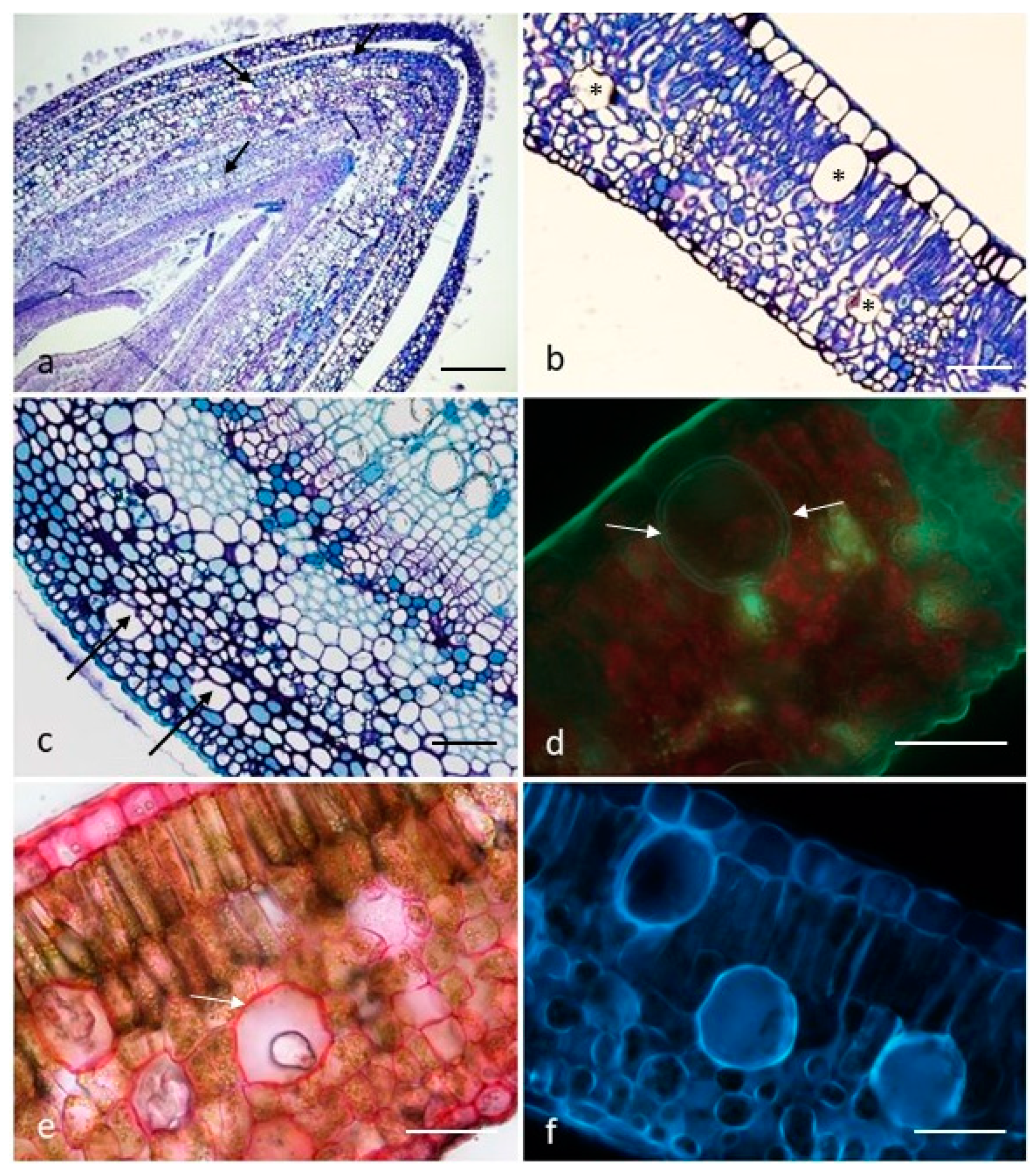
2.1. Micro-Morphological Investigation
2.2. Phytochemical Investigation
2.3. Antibacterial Activity
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chemicals
4.3. Morphological Investigation
Light Microscopy (LM)
4.4. Phytochemical Investigation
4.4.1. Essential oilIsolation
4.4.2. GC-MS Analysis
4.5. Antibacterial Activity
4.5.1. Bacterial Strains Employed
4.5.2. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Determinations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ravindran, P.N.; Nirmal Babu, K.; Shylaja, M. (Eds.) Cinnamon and Cassia, the Genus Cinnamomum; CRC Press: Boca Raton, FL, USA, 2004; ISBN 978-0415317559. [Google Scholar]
- Windadri, I.F.; Rahayu, B.S.S. Cinnamomum camphora (L) J.S. Presl. In Plant Resources of South-East Asia, No. 19, Essential Oil Plants; Oyen, I.P.A., Dung, N.X., Eds.; Backhuys Pub.: Leiden, The Netherlands, 1999; pp. 74–78. ISBN 978-9057820106. [Google Scholar]
- Shanshan, G.; Zhufeng, G.; Wenjuan, Z.; Junyu, L.; Chengfang, W.; Zhiwei, D.; Shushan, D. The Chemical Composition of Essential Oils from Cinnamomum camphora and Their Insecticidal Activity against the Stored Product Pests. Int. J. Mol. Sci. 2016, 17, 1836. [Google Scholar] [CrossRef]
- Rangari, P. Screening of antibacterial sensitivity of essential oils of Camphor and Cinnamon. Shodh Anusandhan Samachar. 2011, 1, 22–26. [Google Scholar]
- Pragadheesh, V.S.; Saroj, A.; Yadav, A.; Chanotiya, C.S.; Alam, M.; Samad, A. Chemical characterization and antifungal activity of Cinnamomum camphora essential oil. Ind. Crop. Prod. 2013, 49, 628–633. [Google Scholar] [CrossRef]
- Satyal, P.; Paudel, P.; Poudel, A.; Dosoky, N.S.; Pokharel, K.K.; Setzer, W.N. Bioactivities an compositional analyses of Cinnamomum essential oil from Nepal: C. camphora, C. tamala, and C. glaucescens. Nat. Prod. Commun. 2013, 8, 1777–1784. [Google Scholar] [CrossRef] [Green Version]
- Shujiang, S.; Qiduan, W.; Jiyan, S.; Chuwen, L.; Xiaoning, Z.; Jianhui, X.; Shuhua, G.; Ziren, S.; Huifang, Z. Composition analysis of volatile oils from flowers, leaves and branches of Cinnamomum camphora chvar. Borneol in China. J. Essent. Oil Res. 2013, 25, 395–401. [Google Scholar] [CrossRef]
- Lee, H.J.; Hyun, E.A.; Yoon, W.J.; Kim, B.H.; Rhee, M.H.; Kang, H.K.; Cho, J.Y.; Yoo, E.S. In vitro anti-inflammatory and anti-oxidative effects of Cinnamomum camphora extracts. J. Ethnopharmacol. 2006, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Jawaid, T. Cinnamomum camphora (Kapur): Review. J. Pharm. 2012, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, X.; Lin, J.; Liu, J.; Jiang, M.; Chu, L. Chemical Composition and Antifungal Activity of Extracts from the Xylem of Cinnamomum camphora. BioResources 2014, 9, 2560–2571. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yan, W. Conservation and applications of camphor tree (Cinnamomum camphora) in China: Ethnobotany and genetic resources. Gen. Resour. Crop Evol. 2015, 63, 1049–1061. [Google Scholar] [CrossRef]
- Grbić, G.; Ćulić, M.; Martać, L.; Soković, M.; Spasić, S.; Doković, D. Effect of Camphor essential oil on rat cerebral cortex activity as manifested by fractal dimension changes. Arch. Biol. Sci. 2008, 60, 547–553. [Google Scholar] [CrossRef]
- Jharna, M.; Mahapatra, N.R. Evaluation of the clinical efficacy and safety of the herbal cold balm in the treatment of the common cold associated with headache. Antiseptic 2009, 106, 87–90. [Google Scholar]
- Geng, S.-L.; Zhao-Xue, C.M.; Bin, S.; Sheng, Z.; Hin-Hua, Y. Histochemistry and cell wall specialization of oil cells related to the essential oil accumulation in the bark of Cinnamomum cassia Presl. (Lauraceae). Plant Product. Sci. 2012, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pino, J.A.; Fuentes, V. Leaf Oil of Cinnamomum camphora (L.) J. Presl. from Cuba. J. Essent. Oil Res. 1998, 10, 531–532. [Google Scholar] [CrossRef]
- Frizzo, C.D.; Santos, A.C.; Paroul, N.; Serafini, L.A.; Dellacassa, E.; Lorenzo, D.; Moyna, P. Essential oils of camphor tree (Cinnamomum camphora Nees & Eberm) Cultivated in Southern Brazil. Braz. Arch. Biol. Technol. 1999, 43, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Wang, J.; Song, L.; Cao, X.; Yao, X.; Tang, F.; Yue, Y. GCxGC-TOFMS Analysis of essential oils composition from leaves, twigs and seeds of Cinnamomum camphora L. Presl and their insecticidal and repellent activities. Molecules 2016, 21, 423. [Google Scholar] [CrossRef] [Green Version]
- Chalchat, J.-C.; Valade, I. Chemical composition of leaf oils of Cinnamomum from Madagascar: C. zeylanicum Blume, C. camphora L., C. fragrans Baillon and C. angustifolium. J. Essent. Oil Res. 2000, 12, 537–540. [Google Scholar] [CrossRef]
- Wanyang, S.; Wei, H.; Guangyu, W. Study on chemical constituents of the essential oil and classification of types from Cinnamomum camphora. Acta Bot. Sin. (China) 1989, 31, 209–214. [Google Scholar]
- Piechulla, B.; Bartelt, R.; Brosemann, A.; Effmert, U.; Bouwmeester, H.; Hippauf, F.; Brandt, W. The α-Terpineol to 1,8-Cineole Cyclization Reaction of Tobacco Terpene Synthases. Plant Physiol. 2016, 172, 2120–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, M.E.; Gerritsen, A.F.; Van Der Schaaf, P.J. Development of oil and mucilage cells in Cinnamomum burmanni. An ultrastructural study. Acta Bot. Neerl. 1991, 40, 339–356. [Google Scholar] [CrossRef]
- Bakker, M.E.; Gerritsen, A.F.; Van Der Schaaf, P.J. Leaf anatomy of Cinnamomum Schaeffer (Lauraceae) with special reference to oil and mucilage cells. Blumea 1992, 37, 1–30. [Google Scholar]
- Stubbs, B.J.; Specht, A.; Brushett, D. The Essential Oil of Cinnamomum camphora (L.) Nees and Eberm.-Variation in Oil Composition Throughout the Tree in Two Chemotypes from Eastern Australia. J. Essent. Oil Res. 2004, 16, 200–205. [Google Scholar] [CrossRef]
- Khosravi Dehaghi, N.; Ostad, S.N.; Maafi, N.; Pedram, S.; Ajani, Y.; Hadjiakhoondi, A.; Khanavi, M. Cytotoxic activity of the essential oil of Salvia verticillata. Res. J. Pharmacogn. 2014, 1, 27–33. [Google Scholar]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowsky, L.F.; Calixto, J.B. Anti-inflammatory effects of compounds alpha-humulene and (-)-trans-caryophyllene isolated from the essential oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef]
- Chaves, J.S.; Leal, P.C.; Pianowisky, L.; Calixto, L.B. Pharmacokinetics and tissue distribution of the sesquiterpene α-humulene in mice. Planta Med. 2008, 74, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Myslivečková, Z.; Szotáková, B.; Špičáková, A.; Lněničková, K.; Ambrož, M.; Kubíček, V.; Krasulová, K.; Anzenbacher, P.; Skálová, L. The inhibitory effects of β-caryophyllene, β-caryophyllene oxide and α-humulene on the activities of the main drug-metabolizing enzymes in rat and human liver in vitro. Chem. Biol. Interact. 2017, 278, 123–128. [Google Scholar] [CrossRef]
- Miller, J.A.; Miller, E.C. The metabolic activation and nucleic acid adducts of naturally-occurring carcinogens: Recent results with ethyl carbamate and the spice flavors safrole and estragole. Br. J. Cancer 1983, 48, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nelson, X.J.; Jackson, R.R. Hunger-driven response by a nectar-eating jumping spider to specific phytochemicals. Chemoecology 2013, 23, 149–153. [Google Scholar] [CrossRef]
- Tabari, M.A.; Khodashenas, A.; Jafari, M.; Petrelli, R.; Cappellacci, L.; Nabissi, M.; Maggi, F.; Pavela, R.; Youssefi, M.R. Acaricidal properties of hemp (Cannabis sativa L.) essential oil against Dermanyssus gallinae and Hyalomma dromedarii. Ind. Crop. Prod. 2020, 147, 112238. [Google Scholar] [CrossRef]
- Neal, J.J. Myristicin, safrole, and fagaramide as phytosynergists of xanthotoxin. J. Chem. Ecol. 1989, 15, 309–315. [Google Scholar] [CrossRef]
- Worth, H.; Schacher, C.; Dethlefsen, U. Concomitant therapy with Cineole (Eucalyptol) reduces exacerbations in COPD: A placebo-controlled double-blind trial. Respir. Res. 2009, 10, 69. [Google Scholar] [CrossRef] [Green Version]
- Juergens, U.R.; Dethlefsen, U.; Steinkamp, G.; Gillissen, A.; Repges, R.; Vetter, H. Anti-inflammatory activity of 1.8-cineol (eucalyptol) in bronchial asthma: A double-blind placebo-controlled trial. Respir. Med. 2003, 97, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.A.; Rao, V.S. Antiinflammatory and antinociceptive effects of 1,8-cineole a terpenoid oxide present in many plant essential oils. Phytother. Res. 2000, 14, 240–244. [Google Scholar] [CrossRef]
- Santos, F.A.; Rao, V.S. 1,8-cineol, a food flavoring agent, prevents ethanol-induced gastric injury in rats. Dig. Dis. Sci. 2001, 46, 331–337. [Google Scholar] [CrossRef]
- Lampronti, I.; Saab, A.M.; Gambari, A. Antiproliferative activity of essential oils derived from plants belonging to the Magnoliophyta division. Int. J. Oncol. 2006, 29, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Sabino, C.K.; Ferreria-Filho, E.S.; Mendes, M.B.; Da Silva Filho, J.C. Cardiovascular effects induced by α-terpineol in hypertensive rats. Flavour Frag. J. 2013, 28, 333–339. [Google Scholar] [CrossRef]
- Khaleel, C.; Tabanca, N.; Buchbauer, G. α-Terpineol, a natural monoterpene: A review of its biological properties. Open Chem. 2018, 16, 349–361. [Google Scholar] [CrossRef]
- De Sousa, D.P.; Quintans, L., Jr.; De Almeida, R.N. Evolution of the Anticonvulsant Activity of α-Terpineol. Pharm. Biol. 2007, 45, 69–70. [Google Scholar] [CrossRef] [Green Version]
- Quintans, L., Jr.; Oliveira, M.G.B.; Santana, M.F.; Santana, M.T.; Guimarães, A.G.; Siqueira, J.S. α-Terpineol reduces nociceptive behavior in mice. Pharm. Biol. 2011, 49, 583–586. [Google Scholar] [CrossRef]
- Souza, R.H.L.; Cardoso, M.S.P.; Menezes, C.T.; Silva, J.P.; De Sousa, D.P.; Batista, J.S. Gastroprotective activity of α-terpineol in two experimental models of gastric ulcer in rats. DARU J. Pharm. Sci. 2011, 19, 277–281. [Google Scholar]
- Held, S.; Schieberle, P.; Somoza, V. Characterization of alpha-terpineol as an anti-inflammatory component of orange juice by in vitro studies using oral buccal cells. J. Agric. Food Chem. 2007, 55, 8040–8046. [Google Scholar] [CrossRef] [PubMed]
- Valente, J.; Zuzarte, M.; Gonçalves, M.J.; Lopes, M.C.; Cavaleiro, C.; Salgueiro, L.; Cruz, M.T. Antifungal, antioxidant and anti-inflammatory activities of Oenanthe crocata L. essential oil. Food Chem. Toxicol. 2013, 2, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Matias, E.F.F.; Alves, E.F.; Silva, M.K.N.; Carvalho, V.R.A.; Figueredo, F.G.; Ferreira, J.V.A.; Coutinho, H.D.M.; Silva, J.M.F.L.; Ribeiro-Filho, J.; Costa, J.G.M. Seasonal variation, chemical composition and biological activity of the essential oil of Cordia verbenacea DC (Boraginaceae) and the sabinene. Ind. Crop. Prod. 2016, 87, 45–53. [Google Scholar] [CrossRef]
- Williams, N.H.; Whitten, W.M. Orchid floral fragrances and male Euglossine bees: Methods and advances in the last sesquidecade. Biol. Bull. 1983, 164, 355–395. [Google Scholar] [CrossRef]
- Borg-Karlson, A.K.; Unelius, C.R.; Valterova, I.; Nilsson, A. Floral fragrance chemistry in the early flowering shrub Daphne mezereum. Phytochemistry 1996, 41, 1477–1483. [Google Scholar] [CrossRef]
- Granero, A.M.; Guerra Sanz, J.M.G.; Gonzalez, F.J.E.; Martinez Vidal, J.L.; Dornhaus, A.; Ghani, J.; Serrano, A.R.; Chittka, L. Chemical compounds of the foraging recruitment pheromone in bumblebees. Naturwissenschaften 2005, 92, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Mitko, L.; Weber, M.G.; Ramirez, S.R.; Hedenström, E.; Wcislo, W.T.; Eltz, T. Olfactory specialization for perfume collection in male orchid bees. J. Exper. Biol. 2016, 219, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Sfara, V.; Zerba, E.N.; Alzogaray, R.A. Fumigant insecticidal activity and repellent effect of five essential oils and seven monoterpenes on first-instar nymphs of Rhodnius prolixus. J. Med. Entomol. 2009, 46, 511–515. [Google Scholar] [CrossRef]
- Mulyaningsih, S.; Sporer, F.; Reichling, J.; Wink, M. Antibacterial activity of essential oils from Eucalyptus and of selected components against multidrug-resistant bacterial pathogens. Pharm. Biol. 2011, 49, 893–899. [Google Scholar] [CrossRef]
- Szmigielski, R.; Cieslak, M.; Rudziński, K.J.; Maciejewska, B. Identification of volatiles from Pinus silvestris attractive for Monochamus galloprovincialis using a SPME-GC/MS platform. Environ. Sci. Pollut. Res. 2012, 19, 2860–2869. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, Y.; Kunoh, H.; Yamamoto, H.; Akimitsu, K. Biological roles of monoterpene volatiles derived from rough lemon (Citrus jambhiri Lush) in citrus defense. J. Gen. Plant. Pathol. 2007, 73, 168–179. [Google Scholar] [CrossRef]
- Singh, S.K.; Strobel, G.A.; Knighton, B.; Geary, B.; Sears, J.; Ezra, D. An endophytic Phomopsis sp. possessing bioactivity and fuel potential with its volatile organic compounds. Microb. Ecol. 2011, 61, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, D.; Huang, X.; Yang, H.; Qiu, Z.; Zou, L.; Yang, C. Study on Antibacterial and Quorum-Sensing Inhibition Activities of Cinnamomum camphora Leaf Essential Oil. Molecules 2019, 24, 3792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merghni, A.; Noumi, E.; Hadded, O.; Dridi, N.; Panwar, H.; Ceylan, O.; Mastouri, M.; Snoussi, M. Assessment of the antibiofilm and antiquorum sensing activities of Eucalyptus globulus essential oil and its main component 1,8-cineole against methicillin-resistant Staphylococcus aureus strains. Microb. Pathog. 2018, 118, 74–80. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Meile, J.C.; Lebrun, M.; Caruso, D.; Chu-Ky, S.; Sarter, S. Litsea cubeba leaf essential oil from Vietnam: Chemical diversity and its impacts on antibacterial activity. Lett. Appl. Microbiol. 2018, 66, 207–214. [Google Scholar] [CrossRef]
- Sayout, A.; Ouarhach, A.; Dilagui, I.; Soraa, N.; Romane, A. Antibacterial activity and chemical composition of essential oil from Lavandula tenuisecta Coss. ex Ball. an endemic species from Morocco. Eur. J. Integr. Med. 2020, 33, 101017. [Google Scholar] [CrossRef]
- Kumar, S.; Kumari, R. Cinnamomum: Review article of essential oil compounds, ethnobotany, antifungal and antibacterial effects. Open Access J. Sci. 2019, 20193, 13–16. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan Red 7B or Fluoral Yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile Red: A selective fluorescent stain for intracellular lipids droplets. J. Cell. Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef] [Green Version]
- David, R.; Carde, J.P. Coloration différentielle des inclusions lipidiques et terpéniques des pseudophylles du Pin maritime au moyen du reactif NADI. C. R. Biol. 1964, 258, 1338–1340. [Google Scholar]
- Jensen, W.A. Botanical Histochemistry: Principles and Practice; WH Freeman & Co: San Francisco, CA, USA, 1962. [Google Scholar]
- Beccari, N.; Mazzi, V. Manual of Microscopic Technique (Manuale di tecnica microscopica); Società Editrice Libraria: Como, Italy, 1966. [Google Scholar]
- Fratini, F.; Mancini, S.; Turchi, B.; Friscia, E.; Pistelli, L.; Giusti, G.; Cerri, D. A novel interpretation of the Fractional Inhibitory Concentration Index: The case of Origanum vulgare L. and Leptospermum scoparium J. R. et G. Forst essential oils against Staphylococcus aureus strains. Microbiol. Res. 2017, 195, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 63–175. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Test | Target Compound Class | Oil Cells | Mucilage Cells | ||
|---|---|---|---|---|---|
| Leaf | Stem | Leaf | Stem | ||
| Fluoral Yellow-088 | Total lipids | ++ | ++ | − | − |
| Nile Red | Neutral lipids | ++ | ++ | − | − |
| NADI reagent | Terpenes | ++ | ++ | − | − |
| Alcian Blue | Acid mucopolysaccharides | − | − | ++ | ++ |
| Naturstoff Reagentz-A | Flavonoids | − | − | − | − |
| Compounds Description | 2017 | 2018 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n. | LRI a | Name | Class | Aerial Parts | FL | FL20 | FL80 | DL | Young Stems | Bark | Fruits |
| 1 | 921 | α-thujene | HM | 0.7 | 0.6 | 0.8 | 0.8 | 0.6 | 0.7 | 0.5 | 0.5 |
| 2 | 927 | α-pinene | HM | 4.3 | 5.0 | 5.4 | 5.4 | 3.7 | 5.0 | 2.6 | 3.1 |
| 3 | 940 | camphene | HM | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | - |
| 4 | 966 | sabinene | HM | 15.6 | 21.0 | 20.0 | 19.7 | 23.3 | 6.7 | 2.8 | 11.7 |
| 5 | 969 | β-pinene | HM | 3.7 | 4.5 | 4.8 | 4.7 | 4.2 | 4.9 | 2.6 | 2.9 |
| 6 | 990 | myrcene | HM | 1.1 | 1.1 | 1.2 | 1.1 | 0.8 | 0.9 | 0.4 | 0.9 |
| 7 | 1004 | α-phellandrene | HM | 0.8 | 0.7 | 0.8 | 0.9 | 0.5 | 2.1 | - | 2.6 |
| 8 | 1014 | α-terpinene | HM | 0.8 | 0.4 | 0.9 | 0.9 | 0.3 | 2.8 | - | 1.0 |
| 9 | 1022 | p-cymene | HM | - | 0.4 | 0.4 | 0.4 | 0.3 | 1.5 | 5.6 | 0.3 |
| 10 | 1023 | o-cymene | PP | 0.4 | - | - | - | - | - | - | - |
| 12 | 1025 | sylvestrene | HM | 0.8 | - | - | - | - | - | - | - |
| 11 | 1026 | limonene | HM | - | 1.3 | 1.3 | 1.3 | 0.8 | 2.3 | 1.3 | 1.6 |
| 13 | 1027 | 1,8-cineole | OM | 38.5 | 46.6 | 48.1 | 46.4 | 63.6 | 53.7 | 68.6 | 40.2 |
| 14 | 1038 | (Z)-β -ocimene | HM | - | - | - | - | - | - | - | 0.1 |
| 15 | 1048 | (E)-β -ocimene | HM | - | - | - | - | - | 0.1 | - | 0.4 |
| 16 | 1056 | γ-terpinene | HM | 1.3 | 0.9 | 1.5 | 1.5 | 0.4 | 4.0 | - | 1.6 |
| 17 | 1065 | cis-sabinene hydrate | HM | 0.6 | 0.1 | - | - | - | - | - | - |
| 18 | 1085 | terpinolene | HM | 0.3 | 0.2 | 0.3 | 0.3 | 0.2 | 0.8 | - | 0.4 |
| 19 | 1118 | cis-p-menth-2-en-1-ol | OM | 0.1 | - | - | - | - | - | - | - |
| 20 | 1165 | δ-terpineol | OM | 0.5 | 0.3 | 0.2 | 0.3 | 0.1 | 0.2 | 0.1 | 0.3 |
| 21 | 1174 | terpinen-4-ol | OM | 2.7 | 1.7 | 2.5 | 2.7 | 0.1 | 7.1 | 6.2 | 4.2 |
| 22 | 1187 | α-terpineol | OM | 17.8 | 13.6 | 10.7 | 12.3 | 1.1 | 5.7 | 6.1 | 22.0 |
| 23 | 1192 | cis-piperitol | OM | 0.1 | - | - | - | - | - | - | - |
| 24 | 1239 | carvone | OM | 0.6 | - | - | - | - | - | - | 0.1 |
| 25 | 1283 | safrole | PP | 1.4 | - | - | -- | - | - | - | 5.1 |
| 26 | 1343 | α-cubebene | HS | - | - | - | - | - | - | 0.1 | - |
| 27 | 1346 | α-terpinyl acetate | OM | 2.3 | 1.2 | 0.8 | 1.1 | - | 0.9 | 0.4 | 0.9 |
| 28 | 1405 | methyl-eugenol | PP | 0.2 | - | - | - | - | - | - | - |
| 29 | 1407 | (E)-caryophyllene | HS | - | - | - | - | - | - | 0.1 | - |
| 30 | 1440 | α-humulene | HS | - | - | - | - | - | - | 1.1 | - |
| 31 | 1475 | α-selinene | HS | 0.2 | - | - | - | - | - | - | - |
| 32 | 1486 | bicyclogermacrene | HS | 0.2 | - | - | - | - | - | - | - |
| 33 | 1492 | α-muurolene | HS | 0.1 | - | - | - | - | - | - | - |
| 34 | 1504 | γ-cadinene | HS | 0.1 | - | - | - | - | - | - | - |
| 35 | 1510 | trans-calamenene | HS | - | - | - | - | - | - | 0.1 | - |
| 36 | 1511 | δ-cadinene | HS | 0.6 | 0.3 | 0.1 | 0.2 | - | 0.2 | 0.1 | 0.2 |
| 37 | 1562 | 8-acetoxy-carvotan acetone | OS | 1.5 | - | - | - | - | - | - | - |
| 38 | 1563 | spathulenol | OS | - | - | - | - | - | - | 0.3 | - |
| 39 | 1565 | germacrene D-4-ol | OS | 0.4 | - | - | - | - | - | - | |
| 40 | 1593 | α-humulene epoxide II | OS | - | - | - | - | - | - | 0.3 | - |
| 41 | 1630 | epi-α-cadinol | OS | - | - | - | - | - | - | 0.2 | - |
| 42 | 1632 | α-muurolol | OS | 0.4 | - | - | - | - | - | - | - |
| 43 | 1638 | β-eudesmol | OS | 0.2 | - | - | - | - | - | - | - |
| 44 | 1642 | α-cadinol | OS | 0.6 | - | - | - | - | - | 0.2 | 0.1 |
| EO yield (%) | 2.39 | 1.17 | 1.38 | 1.40 | 0.62 | 0.37 | 0.09 | 3.35 | |||
| Monoterpenes (HM) | 29.5 | 36.2 | 37.5 | 37.1 | 35.1 | 32.1 | 15.9 | 26.9 | |||
| Oxygenated monoterpenes (OM) | 63.4 | 63.4 | 62.4 | 62.7 | 64.9 | 67.6 | 81.5 | 67.7 | |||
| Sesquiterpenes (HS) | 1.2 | 0.3 | 0.1 | 0.2 | - | 0.2 | 1.5 | 0.2 | |||
| Oxygenated sesquiterpenes (OS) | 3.0 | - | - | - | - | - | 0.1 | 0.1 | |||
| Phenyl propanoids (PP) | 1.9 | - | - | - | - | - | - | 5.1 | |||
| Total | 99.0 | 99.9 | 100.0 | 100.0 | 100.0 | 100.0 | 99.8 | 100.0 | |||
| Activity | Staphylococcus aureus | Listeria monocytogenes | Enterococcus faecalis | Salmonella tiphymurium | Escherichia coli | Pseudomonas aeruginosa |
|---|---|---|---|---|---|---|
| MIC values | 25.000 | 12.500 | 25.000 | 25.000 | 25.000 | 25.000 |
| MBC values | 50.000 | 50.000 | 50.000 | 25.000 | 50.000 | 50.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bottoni, M.; Milani, F.; Mozzo, M.; Radice Kolloffel, D.A.; Papini, A.; Fratini, F.; Maggi, F.; Santagostini, L. Sub-Tissue Localization of Phytochemicals in Cinnamomum camphora (L.) J. Presl. Growing in Northern Italy. Plants 2021, 10, 1008. https://doi.org/10.3390/plants10051008
Bottoni M, Milani F, Mozzo M, Radice Kolloffel DA, Papini A, Fratini F, Maggi F, Santagostini L. Sub-Tissue Localization of Phytochemicals in Cinnamomum camphora (L.) J. Presl. Growing in Northern Italy. Plants. 2021; 10(5):1008. https://doi.org/10.3390/plants10051008
Chicago/Turabian StyleBottoni, Martina, Fabrizia Milani, Marta Mozzo, Daniele Armando Radice Kolloffel, Alessio Papini, Filippo Fratini, Filippo Maggi, and Laura Santagostini. 2021. "Sub-Tissue Localization of Phytochemicals in Cinnamomum camphora (L.) J. Presl. Growing in Northern Italy" Plants 10, no. 5: 1008. https://doi.org/10.3390/plants10051008
APA StyleBottoni, M., Milani, F., Mozzo, M., Radice Kolloffel, D. A., Papini, A., Fratini, F., Maggi, F., & Santagostini, L. (2021). Sub-Tissue Localization of Phytochemicals in Cinnamomum camphora (L.) J. Presl. Growing in Northern Italy. Plants, 10(5), 1008. https://doi.org/10.3390/plants10051008