
Salinity Stress Affects Photosynthesis, Malondialdehyde Formation, and Proline Content in Portulaca oleracea L.



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
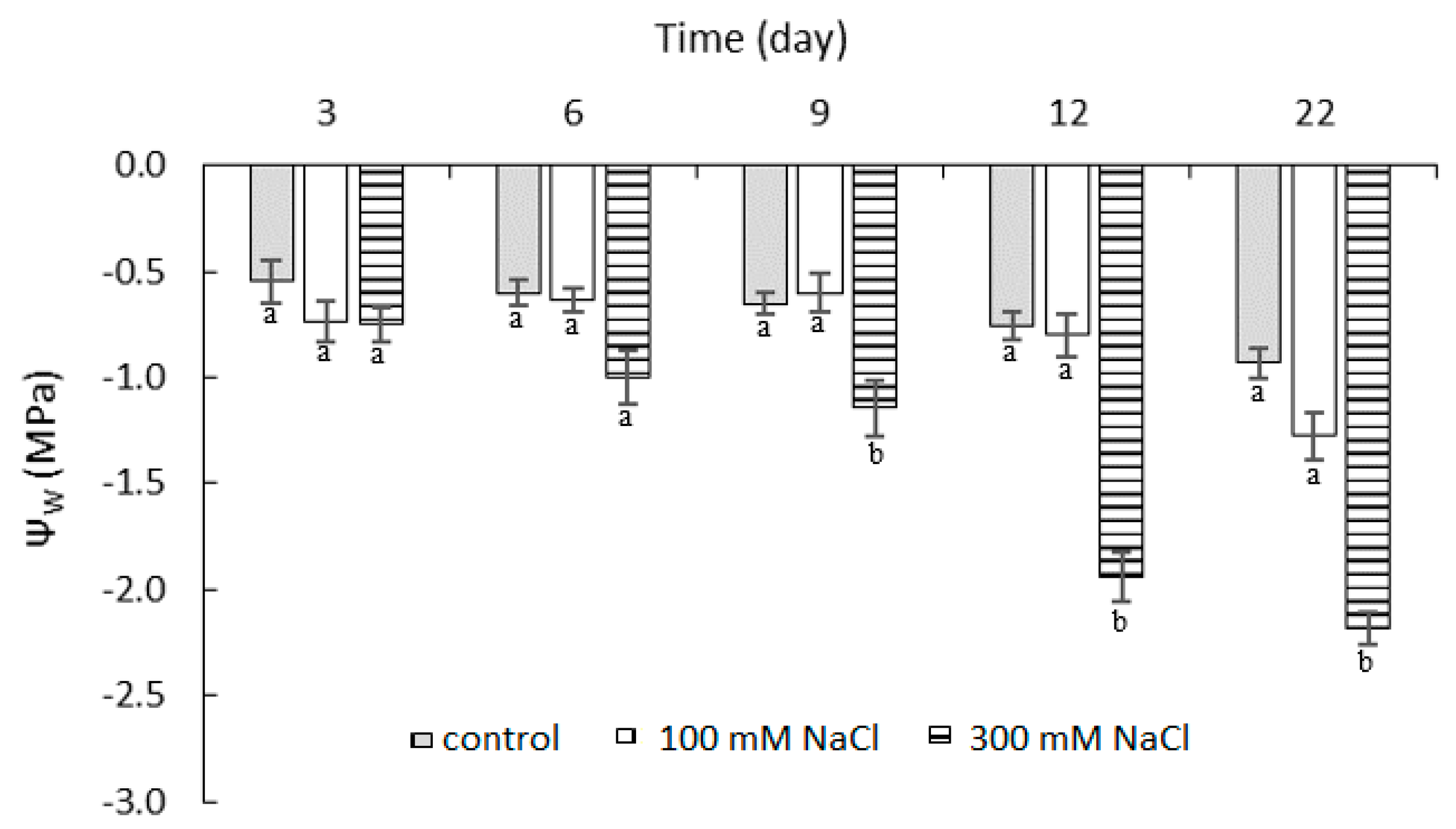
2.1. Effect of NaCl Concentration and Salt Stress Exposure Time on Leaf Water Potential
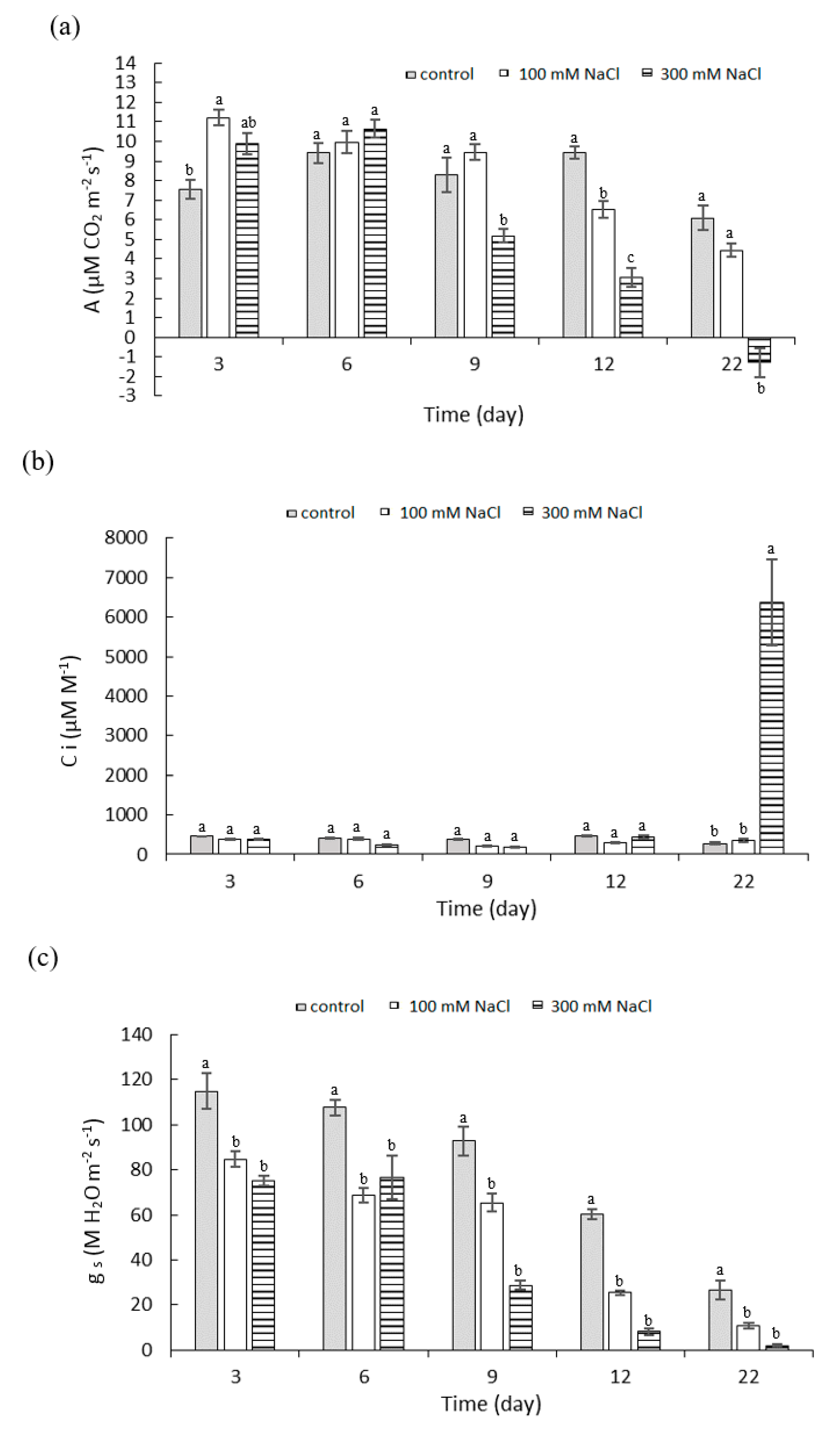
2.2. Effect of NaCl Concentration and Salt Stress Exposure Time on Leaf Gas Exchange Parameters
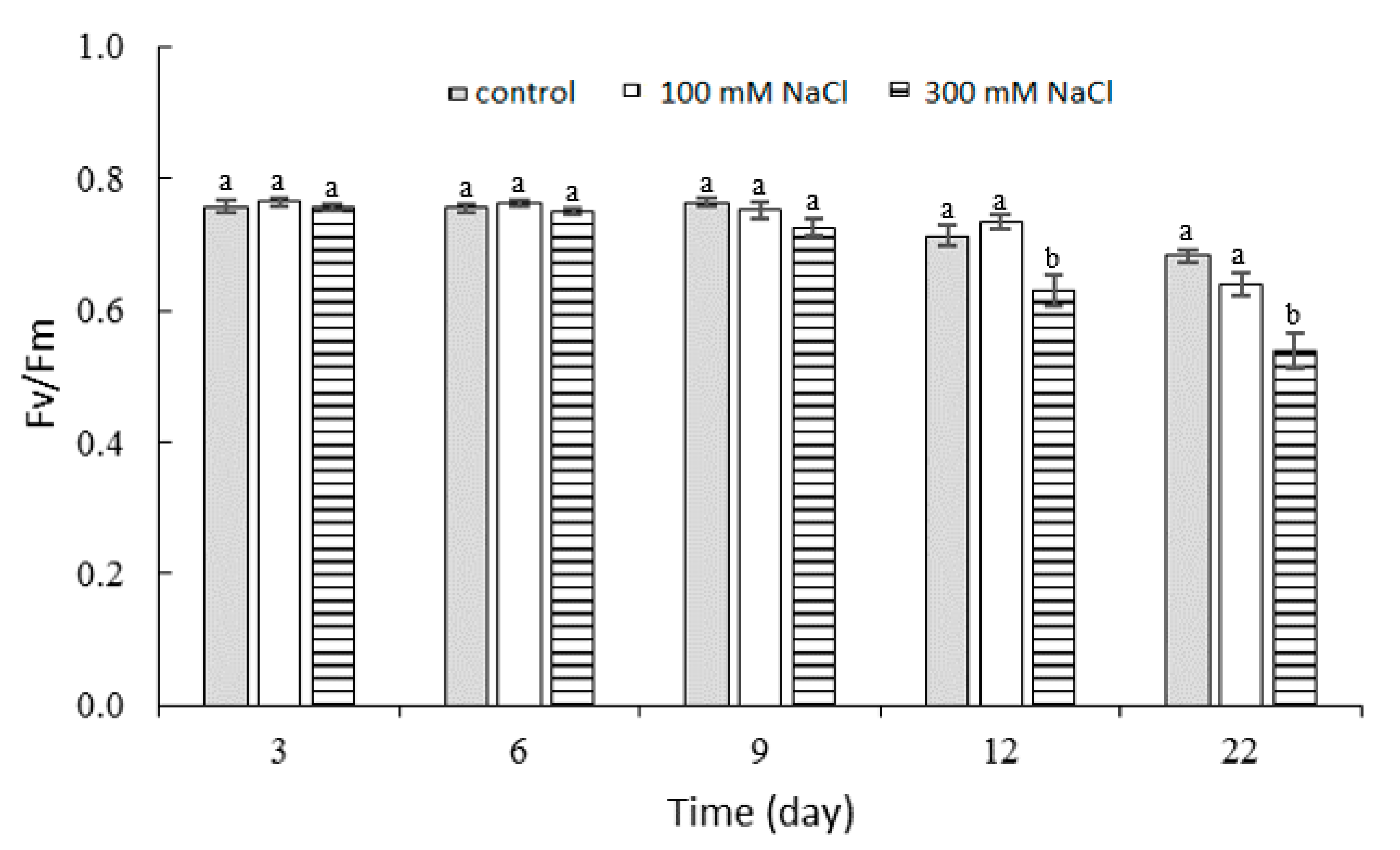
2.3. Effect of NaCl Concentration and Salt Stress Exposure Time on Chlorophyll Fluorescence
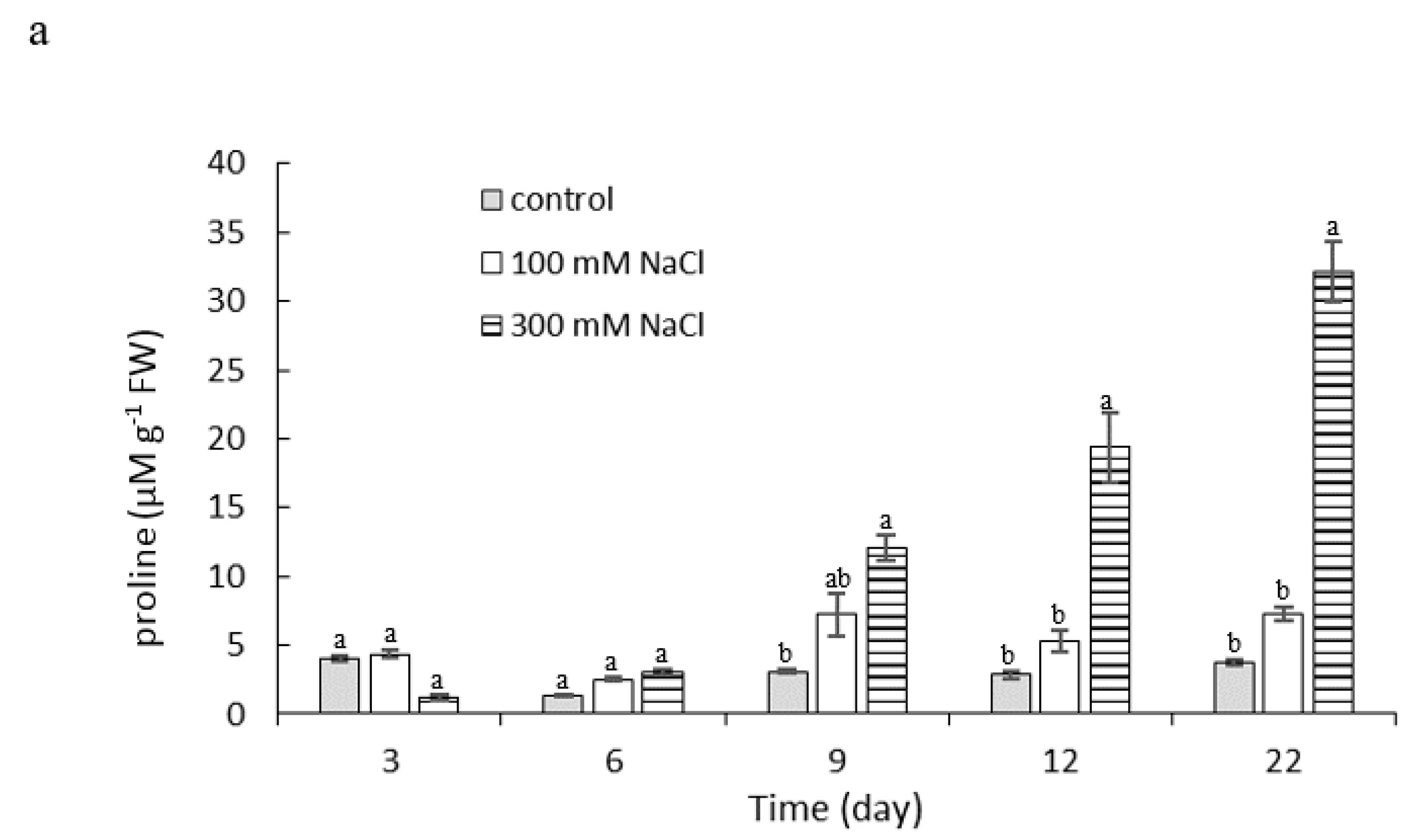
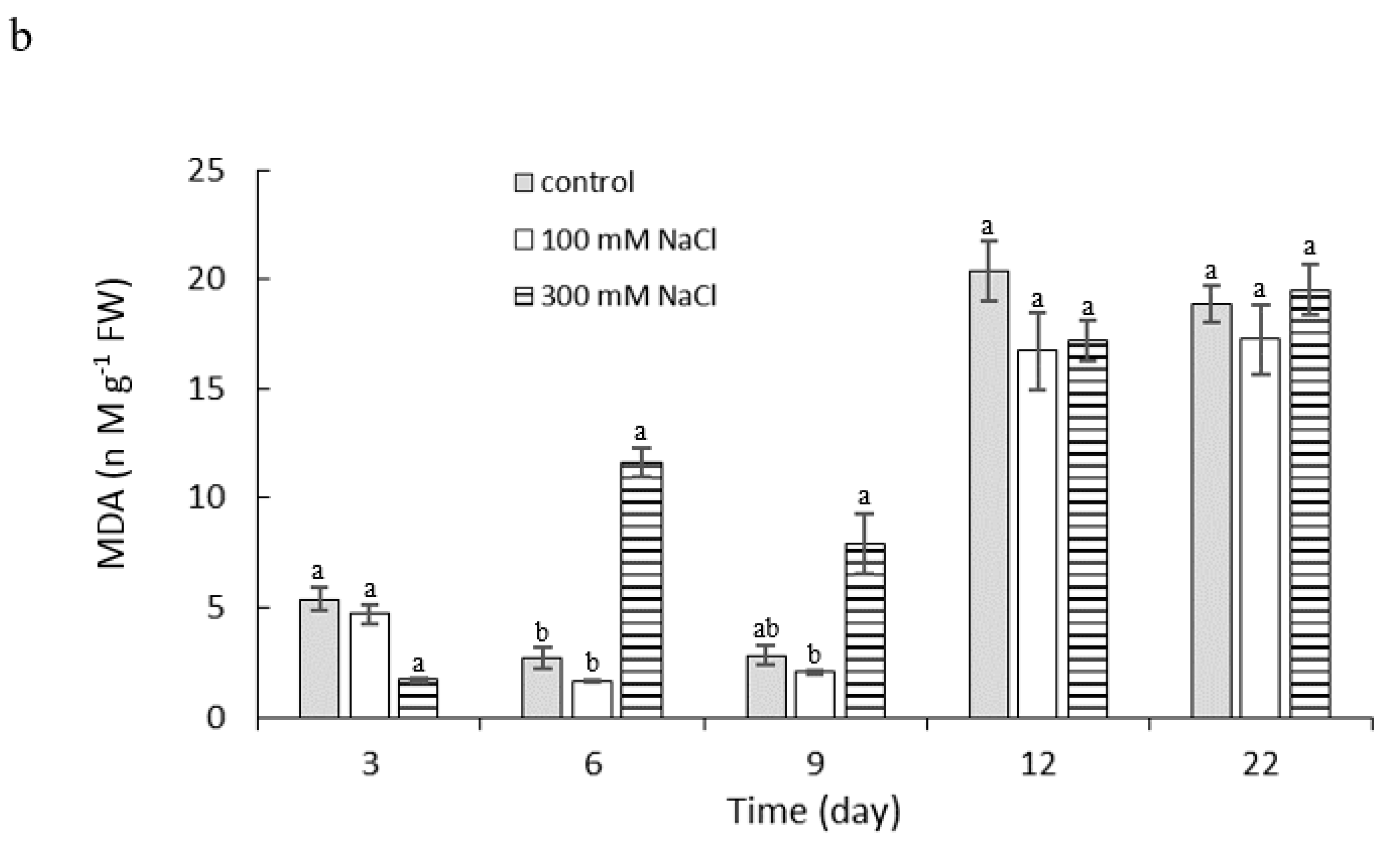
2.4. Effect of NaCl Concentration and Salt Stress Exposure Time on Proline and Malondialdehyde Content
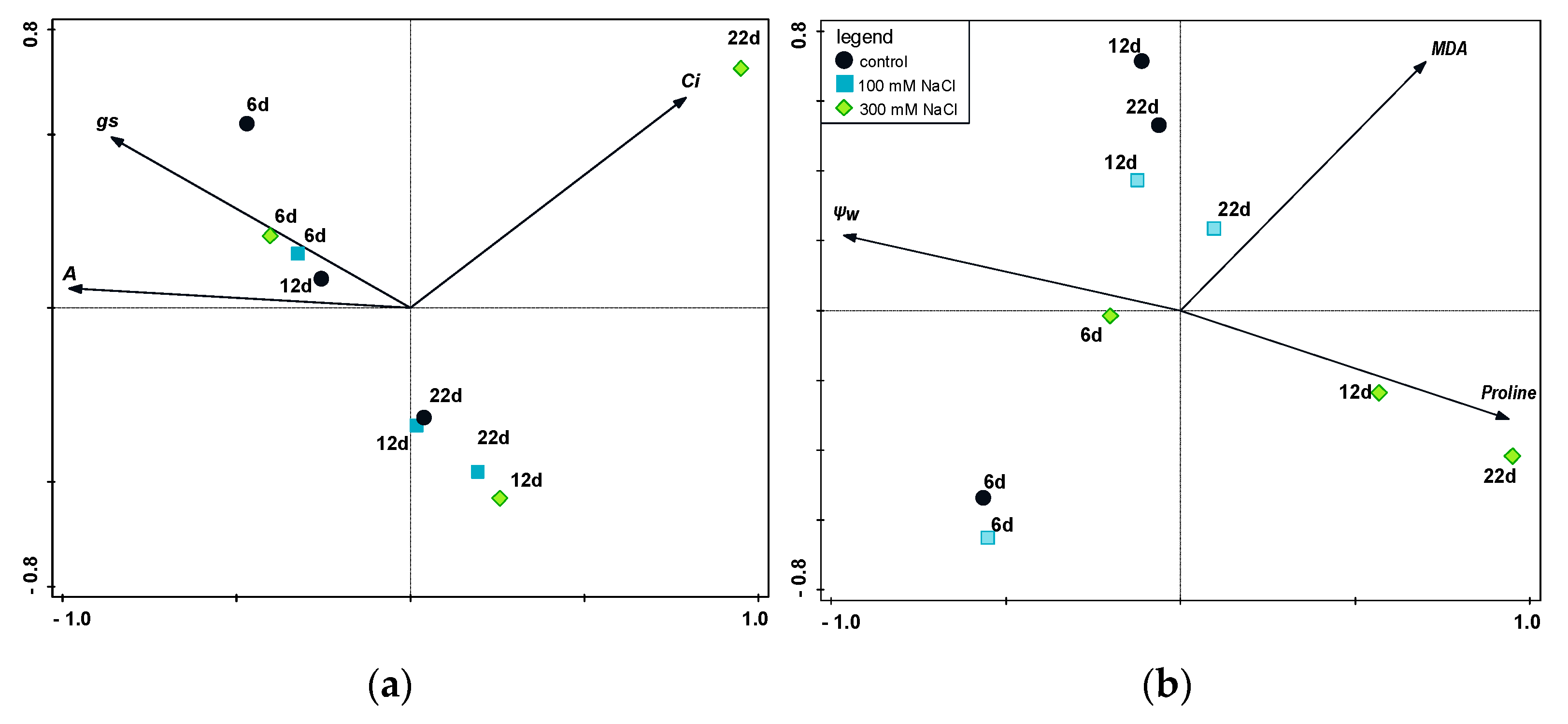
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Salinity Treatments
4.2. Leaf Water Potential
4.3. Leaf Gas Exchange
4.4. Chlorophyll Fluorescence
4.5. Proline Content
4.6. Malondialdehyde (MDA)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shahbaz, M.; Ashraf, M. Improving Salinity Tolerance in Cereals. Crit. Rev. Plant Sci. 2013, 32, 237–249. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil Salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as One of the Tools for Its Alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops—What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Plant Salt Tolerance: Adaptations in Halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Akbarimoghaddam, H.; Galavi, M.; Ghanbari, A.; Panjehkeh, N. Salinity Effects on Seed Germination and Seedling Growth of Bread Wheat Cultivars. Trakia J. Sci. 2011, 9, 43–50. [Google Scholar]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating Physiological Responses of Plants to Salinity Stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the Molecular Mechanisms Mediating Plant Salt-Stress Responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Hnilickova, H.; Hnilicka, F.; Martinkova, J.; Kraus, K. Effects of Salt Stress on Water Status, Photosynthesis and Chlorophyll Fluorescence of Rocket. Plant Soil Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhou, G. Responses of Leaf Stomatal Density to Water Status and Its Relationship with Photosynthesis in a Grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a Fluorescence Study Revealing Effects of High Salt Stress on Photosystem II in Wheat Leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Rastogi, A.; Zivcak, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt Chlorophyll Fluorescence as a Tool for Crop Phenotyping: An Example of Barley Landraces Exposed to Various Abiotic Stress Factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Biswal, B.; Joshi, P.N.; Raval, M.K.; Biswal, U.C. Photosynthesis, a Global Sensor of Environmental Stress in Green Plants: Stress Signalling and Adaptation. Curr. Sci. 2011, 101, 47–56. [Google Scholar]
- Hossain, M.S.; Dietz, K.-J. Tuning of Redox Regulatory Mechanisms, Reactive Oxygen Species and Redox Homeostasis under Salinity Stress. Front. Plant Sci. 2016, 7, 548. [Google Scholar] [CrossRef] [Green Version]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen Under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Saibi, W.; Feki, K.; Ben Mahmoud, R.; Brini, F. Durum Wheat Dehydrin (DHN-5) Confers Salinity Tolerance to Transgenic Arabidopsis Plants through the Regulation of Proline Metabolism and ROS Scavenging System. Planta 2015, 242, 1187–1194. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Salinas, R.; Sánchez, E.; Ruíz, J.M.; Lao, M.T.; Romero, L. Proline, Betaine, and Choline Responses to Different Phosphorus Levels in Green Bean. Commun. Soil Sci. Plant Anal. 2013, 44, 465–472. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Vries, R.S.M.; de Boer, A.H. Effect of Salt Stress on Growth, Na+ Accumulation and Proline Metabolism in Potato (Solanum Tuberosum) Cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef]
- Sarabi, B.; Bolandnazar, S.; Ghaderi, N.; Ghashghaie, J. Genotypic Differences in Physiological and Biochemical Responses to Salinity Stress in Melon (Cucumis Melo L.) Plants: Prospects for Selection of Salt Tolerant Landraces. Plant Physiol. Biochem. 2017, 119, 294–311. [Google Scholar] [CrossRef] [PubMed]
- De la Torre-González, A.; Montesinos-Pereira, D.; Blasco, B.; Ruiz, J.M. Influence of the Proline Metabolism and Glycine Betaine on Tolerance to Salt Stress in Tomato (Solanum Lycopersicum L.) Commercial Genotypes. J. Plant Physiol. 2018, 231, 329–336. [Google Scholar] [CrossRef]
- Hannachi, S.; Van Labeke, M.-C. Salt Stress Affects Germination, Seedling Growth and Physiological Responses Differentially in Eggplant Cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Anower, M.R.; Peel, M.D.; Mott, I.W.; Wu, Y. Physiological Processes Associated with Salinity Tolerance in an Alfalfa Half-Sib Family. J. Agron. Crop Sci. 2017, 203, 506–518. [Google Scholar] [CrossRef]
- Zhou, Y.-X.; Xin, H.-L.; Rahman, K.; Wang, S.-J.; Peng, C.; Zhang, H. Portulaca Oleracea L.: A Review of Phytochemistry and Pharmacological Effects. Biomed. Res. Int. 2015, 2015, 925631. [Google Scholar] [CrossRef] [Green Version]
- Kafi, M.; Rahimi, Z. Effect of Salinity and Silicon on Root Characteristics, Growth, Water Status, Proline Content and Ion Accumulation of Purslane (Portulaca oleracea L.). Soil Sci. Plant Nutr. 2011, 57, 341–347. [Google Scholar] [CrossRef]
- Alam, M.A.; Juraimi, A.S.; Rafii, M.Y.; Abdul Hamid, A.; Aslani, F. Screening of Purslane (Portulaca Oleracea L.) Accessions for High Salt Tolerance. Sci. World J. 2014, 2014, e627916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, R.; Wang, Y.; Liu, R.; Gou, J.; Chan, Z. Physiological and Metabolic Changes of Purslane (Portulaca Oleracea L.) in Response to Drought, Heat, and Combined Stresses. Front Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Hnilickova, H.; Hnilicka, F.; Orsak, M.; Hejnak, V. Effect of Salt Stress on Growth, Electrolyte Leakage, Na+ and K+ Content in Selected Plant Species. Plant Soil Environ. 2019, 65, 90–96. [Google Scholar] [CrossRef] [Green Version]
- FAO. High Level Expert Forum—How to Feed the World in 2050; Economic and Social Development, Food and Agricultural Organization of the United Nations: Rome, Italy, 2009. [Google Scholar]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an Inflorescence Meristem-Specific Cytokinin Oxidase—OsCKX2 in Rice Reduces Yield Penalty under Salinity Stress Condition. Plant Cell Environ. 2018, 41, 936–946. [Google Scholar] [CrossRef]
- Gupta, B.K.; Sahoo, K.K.; Ghosh, A.; Tripathi, A.K.; Anwar, K.; Das, P.; Singh, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Manipulation of Glyoxalase Pathway Confers Tolerance to Multiple Stresses in Rice. Plant Cell Environ. 2018, 41, 1186–1200. [Google Scholar] [CrossRef]
- Bunce, J.A. How Do Leaf Hydraulics Limit Stomatal Conductance at High Water Vapour Pressure Deficits? Plant Cell Environ. 2006, 29, 1644–1650. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Figueroa, M.E.; Davy, A.J. Salt Stimulation of Growth and Photosynthesis in an Extreme Halophyte, Arthrocnemum Macrostachyum. Plant Biol. 2010, 12, 79–87. [Google Scholar] [CrossRef]
- Xing, J.-C.; Dong, J.; Wang, M.-W.; Liu, C.; Zhao, B.-Q.; Wen, Z.-G.; Zhu, X.-M.; Ding, H.-R.; Zhao, X.-H.; Hong, L.-Z. Effects of NaCl Stress on Growth of Portulaca Oleracea and Underlying Mechanisms. Braz. J. Bot. 2019, 2, 217–226. [Google Scholar] [CrossRef]
- Tang, N.; Zhang, B.; Chen, Q.; Yang, P.; Wang, L.; Qian, B. Effect of Salt Stress on Photosynthetic and Antioxidant Characteristics in Purslane (Portulaca oleracea). Int. J. Agric. Biol. 2020, 24, 1309–1314. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Ocampo, G.; Koteyeva, N.K.; Voznesenskaya, E.V.; Edwards, G.E.; Sage, T.L.; Sage, R.F.; Columbus, J.T. Evolution of Leaf Anatomy and Photosynthetic Pathways in Portulacaceae. Am. J. Bot. 2013, 100, 2388–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voznesenskaya, E.V.; Koteyeva, N.K.; Edwards, G.E.; Ocampo, G. Revealing Diversity in Structural and Biochemical Forms of C4 Photosynthesis and a C3-C4 Intermediate in Genus portulaca L. (Portulacaceae). J. Exp. Bot. 2010, 61, 3647–3662. [Google Scholar] [CrossRef] [PubMed]
- Winter, K.; Holtum, J.A.M. Facultative Crassulacean Acid Metabolism (CAM) Plants: Powerful Tools for Unravelling the Functional Elements of CAM Photosynthesis. J. Exp. Bot. 2014, 65, 3425–3441. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, R.M.; Andreo, C.S.; Lara, M.V. Deciphering the Mechanisms Involved in Portulaca oleracea (C4) Response to Drought: Metabolic Changes Including Crassulacean Acid-like Metabolism Induction and Reversal upon Re-Watering. Physiol. Plant 2014, 152, 414–430. [Google Scholar] [CrossRef]
- Montero, E.; Francisco, A.M.; Montes, E.; Herrera, A. Salinity Induction of Recycling Crassulacean Acid Metabolism and Salt Tolerance in Plants of Talinum Triangulare. Ann. Bot. 2018, 121, 1333–1342. [Google Scholar] [CrossRef]
- Osmond, C.B. Crassulacean Acid Metabolism: A Curiosity in Context. Ann. Rev. Plant Physiol. 1978, 29, 379–414. [Google Scholar] [CrossRef]
- Ferrari, R.C.; Bittencourt, P.P.; Rodrigues, M.A.; Moreno-Villena, J.J.; Alves, F.R.R.; Gastaldi, V.D.; Boxall, S.F.; Dever, L.V.; Demarco, D.; Andrade, S.C.S.; et al. C4 and Crassulacean Acid Metabolism within a Single Leaf: Deciphering Key Components behind a Rare Photosynthetic Adaptation. New Phytol. 2020, 225, 1699–1714. [Google Scholar] [CrossRef]
- Cockburn, W. Stomatal Mechanism as the Basis of the Evolution of CAM and C4 Photosynthesis. Plant Cell Environ. 1983, 6, 275–279. [Google Scholar] [CrossRef]
- Borland, A.M.; Zambrano, V.A.B.; Ceusters, J.; Shorrock, K. The Photosynthetic Plasticity of Crassulacean Acid Metabolism: An Evolutionary Innovation for Sustainable Productivity in a Changing World. New Phytol. 2011, 191, 619–633. [Google Scholar] [CrossRef]
- Franco, A.; Herzog, B.; Hübner, C.; de Mattos, E.; Scarano, F.; Ball, E.; Lüttge, U. Diurnal Changes in Chlorophyll a Fluorescence, CO2-Exchange and Organic Acid Decarboxylation in the Tropical CAM Tree Clusia Hilariana. Tree Physiol. 1999, 19, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Herrera, A. Crassulacean Acid Metabolism and Fitness under Water Deficit Stress: If Not for Carbon Gain, What Is Facultative CAM Good For? Ann. Bot. 2009, 103, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, M.V.; Drincovich, M.F.; Andreo, C.S. Induction of a Crassulacean Acid-like Metabolism in the C4 Succulent Plant, Portulaca Oleracea L.: Study of Enzymes Involved in Carbon Fixation and Carbohydrate Metabolism. Plant Cell Physiol. 2004, 45, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, D.E.; Costa, M.L.; Guiamet, J.J. Senescence-Associated Degradation of Chloroplast Proteins inside and Outside the Organelle. Plant Biol. 2008, 10, 15–22. [Google Scholar] [CrossRef]
- Karkanis, A.C.; Petropoulos, S.A. Physiological and Growth Responses of Several Genotypes of Common Purslane (Portulaca Oleracea L.) under Mediterranean Semi-Arid Conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of Chlorophyll Fluorescence Can Improve Crop Production Strategies: An Examination of Future Possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.; Guo, S.-R.; Sun, J.; Yuan, L.-Y. Effects of Salt Stress on the Structure and Function of the Photosynthetic Apparatus in Cucumis Sativus and Its Protection by Exogenous Putrescine. Physiol. Plant 2012, 146, 285–296. [Google Scholar] [CrossRef]
- Jamil, M.; Rehman, S.-U.; Lee, K.J.; Kim, J.M.; Kim, H.-S.; Rha, E.S. Salinity Reduced Growth PS2 Photochemistry and Chlorophyll Content in Radish. Sci. Agric. 2007, 64, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Naeem, M.S.; Jin, Z.L.; Wan, G.L.; Liu, D.; Liu, H.B.; Yoneyama, K.; Zhou, W.J. 5-Aminolevulinic Acid Improves Photosynthetic Gas Exchange Capacity and Ion Uptake under Salinity Stress in Oilseed Rape (Brassica napus L.). Plant Soil 2010, 332, 405–415. [Google Scholar] [CrossRef]
- Zaman, S.; Shah, S.B.; Jiang, Y.T.; Che, S.Q. Saline Conditions Alter Morpho-Physiological Intensification in Purslane (Portulaca oleracea L.). J. Biol. Regul. Homeost. Agents 2018, 32, 635–639. [Google Scholar]
- Reddy, A.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Liu, H.; Sultan, M.A.R.F.; Liu, X.-l.; Zhang, J.; Yu, F.; Zhao, H.-x. Physiological and Comparative Proteomic Analysis Reveals Different Drought Responses in Roots and Leaves of Drought-Tolerant Wild Wheat (Triticum boeoticum). PLoS ONE 2015, 10, e0121852. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and Temporal Profile of Glycine Betaine Accumulation in Plants Under Abiotic Stresses. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kumar, J.; Singh, S.; Singh, D.V.; Prasad, S. Roles of Osmoprotectants in Improving Salinity and Drought Tolerance in Plants: A Review. Rev. Environ. Sci. Bio/Technol. 2015, 14. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Mulry, K.R.; Hanson, B.A.; Dudle, D.A. Alternative Strategies in Response to Saline Stress in Two Varieties of Portulaca Oleracea (Purslane). PLoS ONE 2015, 10, e0138723. [Google Scholar] [CrossRef]
- Yazici, I.; Türkan, I.; Sekmen, A.H.; Demiral, T. Salinity Tolerance of Purslane (Portulaca oleracea L.) Is Achieved by Enhanced Antioxidative System, Lower Level of Lipid Peroxidation and Proline Accumulation. Environ. Exp. Bot. 2007, 61, 49–57. [Google Scholar] [CrossRef]
- Ma, J.; Du, G.; Li, X.; Zhang, C.; Guo, J. A Major Locus Controlling Malondialdehyde Content under Water Stress Is Associated with Fusarium Crown Rot Resistance in Wheat. Mol. Genet. Genom. 2015, 290, 1955–1962. [Google Scholar] [CrossRef]
- Jakhar, S.; Mukherjee, D. Chloroplast Pigments, Proteins, Lipid Peroxidation and Activities of Antioxidative Enzymes during Maturation and Senescence of Leaves and Reproductive Organs of Cajanus Cajan L. Physiol. Mol. Biol. Plants 2014, 20, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Borsai, O.; Hassan, M.A.; Negrușier, C.; Raigón, M.D.; Boscaiu, M.; Sestraș, R.E.; Vicente, O. Responses to Salt Stress in Portulaca: Insight into Its Tolerance Mechanisms. Plants. 2020, 9, 1660. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified Thiobarbituric Acid Assay for Measuring Lipid Oxidation in Sugar-Rich Plant Tissue Extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 978-1-107-69440-8. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity Stress Affects Photosynthesis, Malondialdehyde Formation, and Proline Content in Portulaca oleracea L. Plants 2021, 10, 845. https://doi.org/10.3390/plants10050845
Hnilickova H, Kraus K, Vachova P, Hnilicka F. Salinity Stress Affects Photosynthesis, Malondialdehyde Formation, and Proline Content in Portulaca oleracea L. Plants. 2021; 10(5):845. https://doi.org/10.3390/plants10050845
Chicago/Turabian StyleHnilickova, Helena, Kamil Kraus, Pavla Vachova, and Frantisek Hnilicka. 2021. "Salinity Stress Affects Photosynthesis, Malondialdehyde Formation, and Proline Content in Portulaca oleracea L." Plants 10, no. 5: 845. https://doi.org/10.3390/plants10050845
APA StyleHnilickova, H., Kraus, K., Vachova, P., & Hnilicka, F. (2021). Salinity Stress Affects Photosynthesis, Malondialdehyde Formation, and Proline Content in Portulaca oleracea L. Plants, 10(5), 845. https://doi.org/10.3390/plants10050845