
A Review of Unreduced Gametes and Neopolyploids in Alfalfa: How to Fill the Gap between Well-Established Meiotic Mutants and Next-Generation Genomic Resources





Abstract
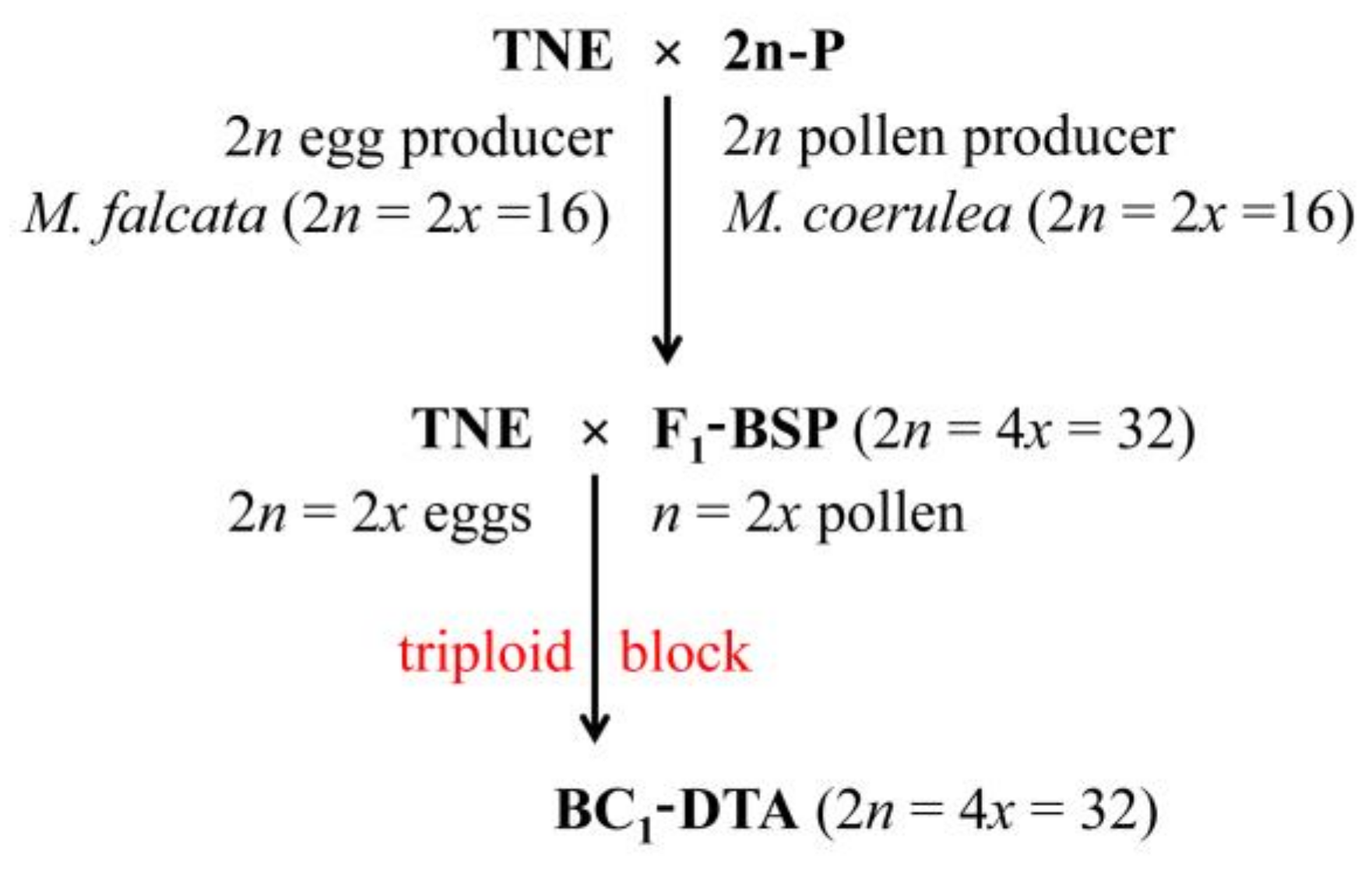
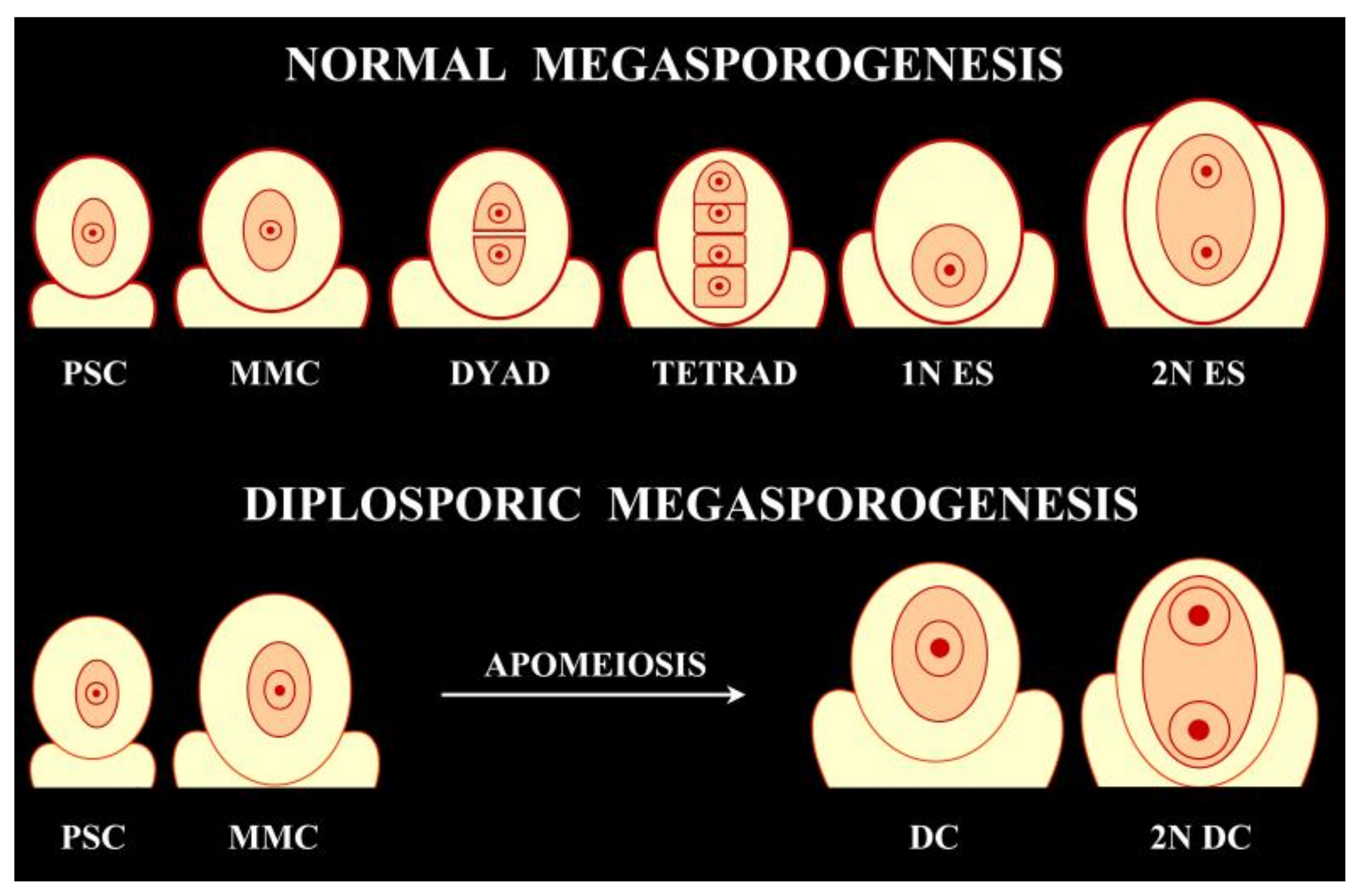
:1. Overview of the Occurrence of 2n Gametes and Their Use for Sexual Polyploidization in the Medicago sativa-coerulea-falcata Complex
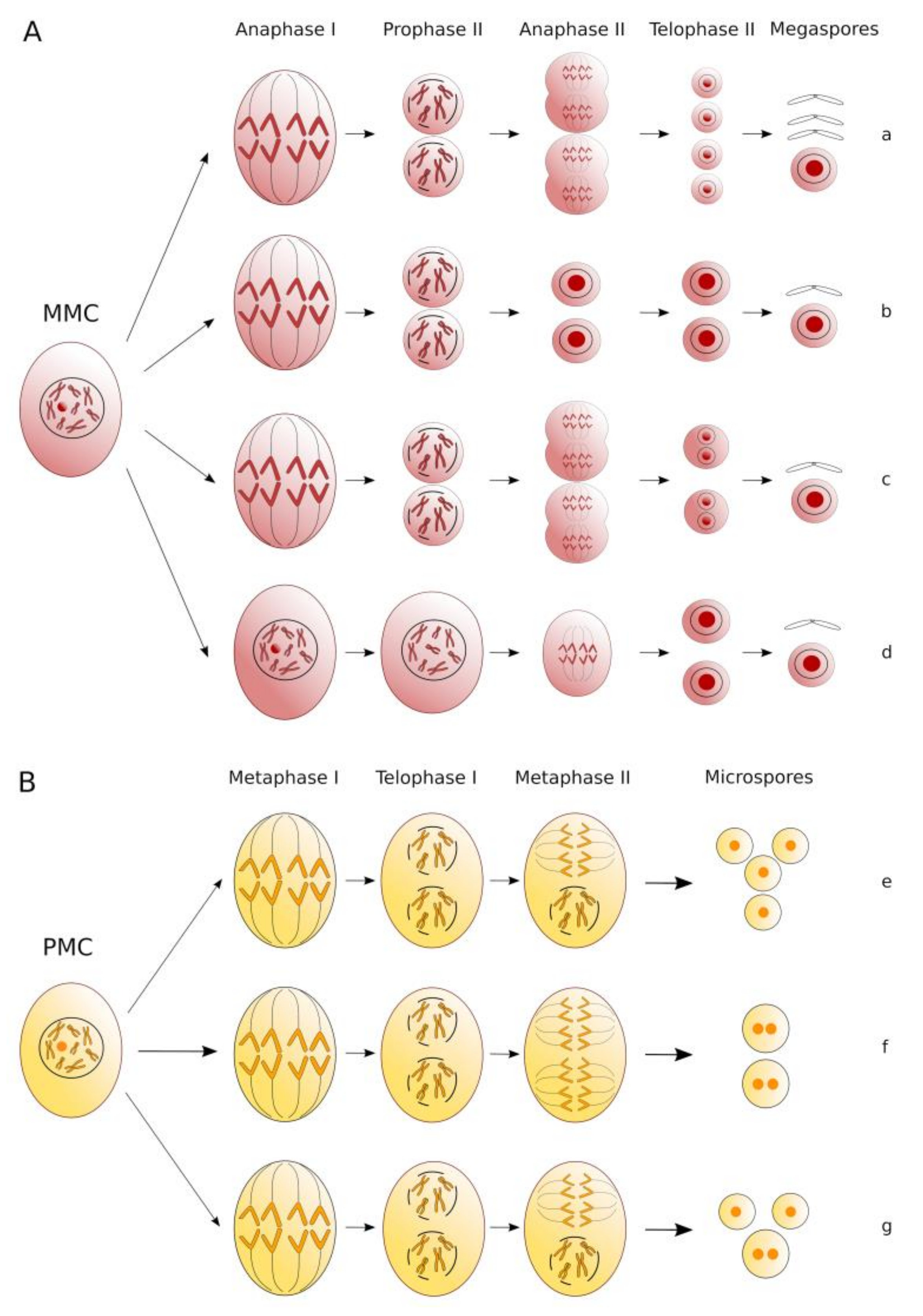
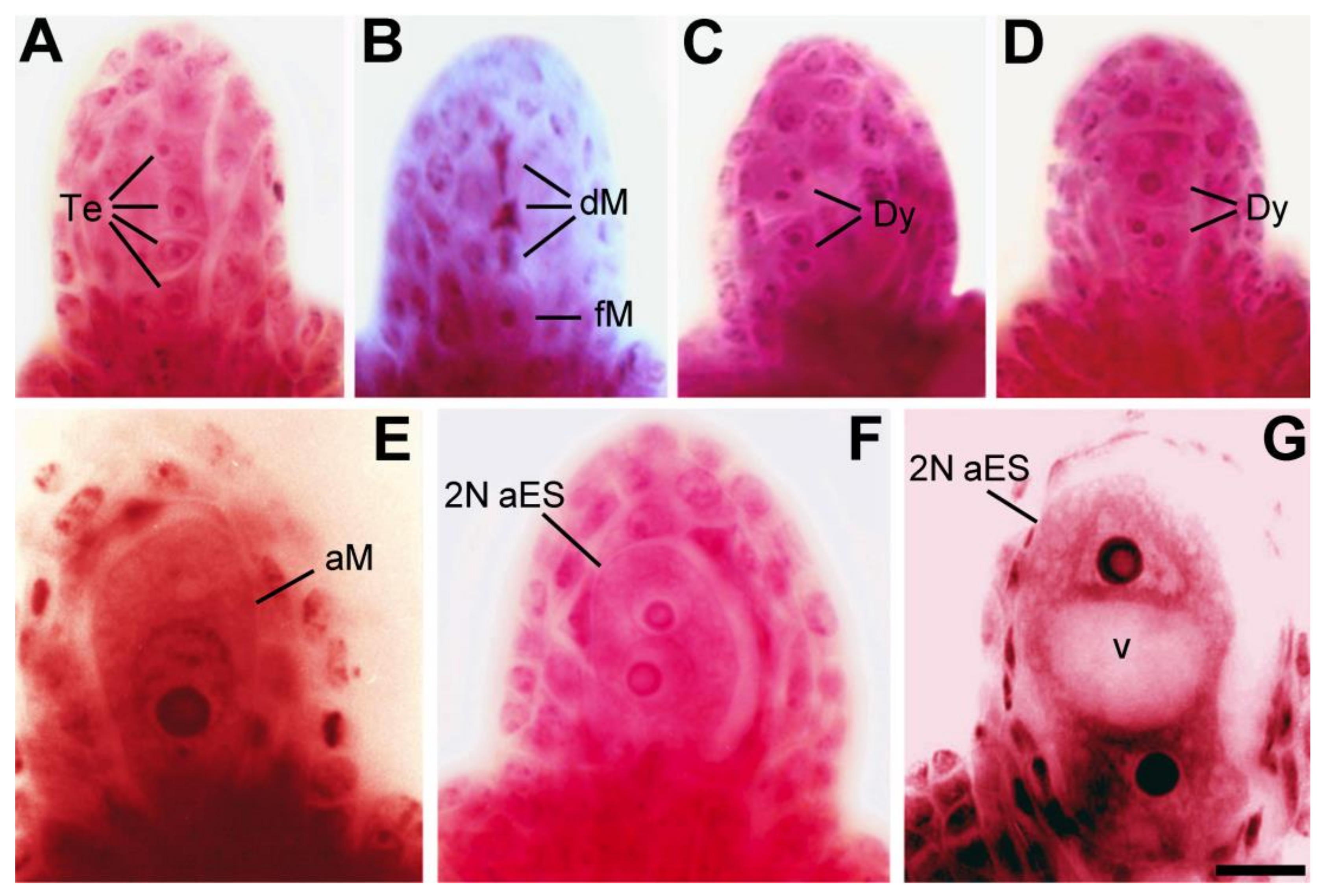
2. Elements of Apomixis in Alfalfa: Sexually Tetraploidized Plants Maintain Apomeiosis Developmentally Independent of Parthenogenesis
3. Candidate Genes for Unreduced Gamete Production and Exploitation of Genomic Resources for the Analysis of Alfalfa Meiotic Mutants
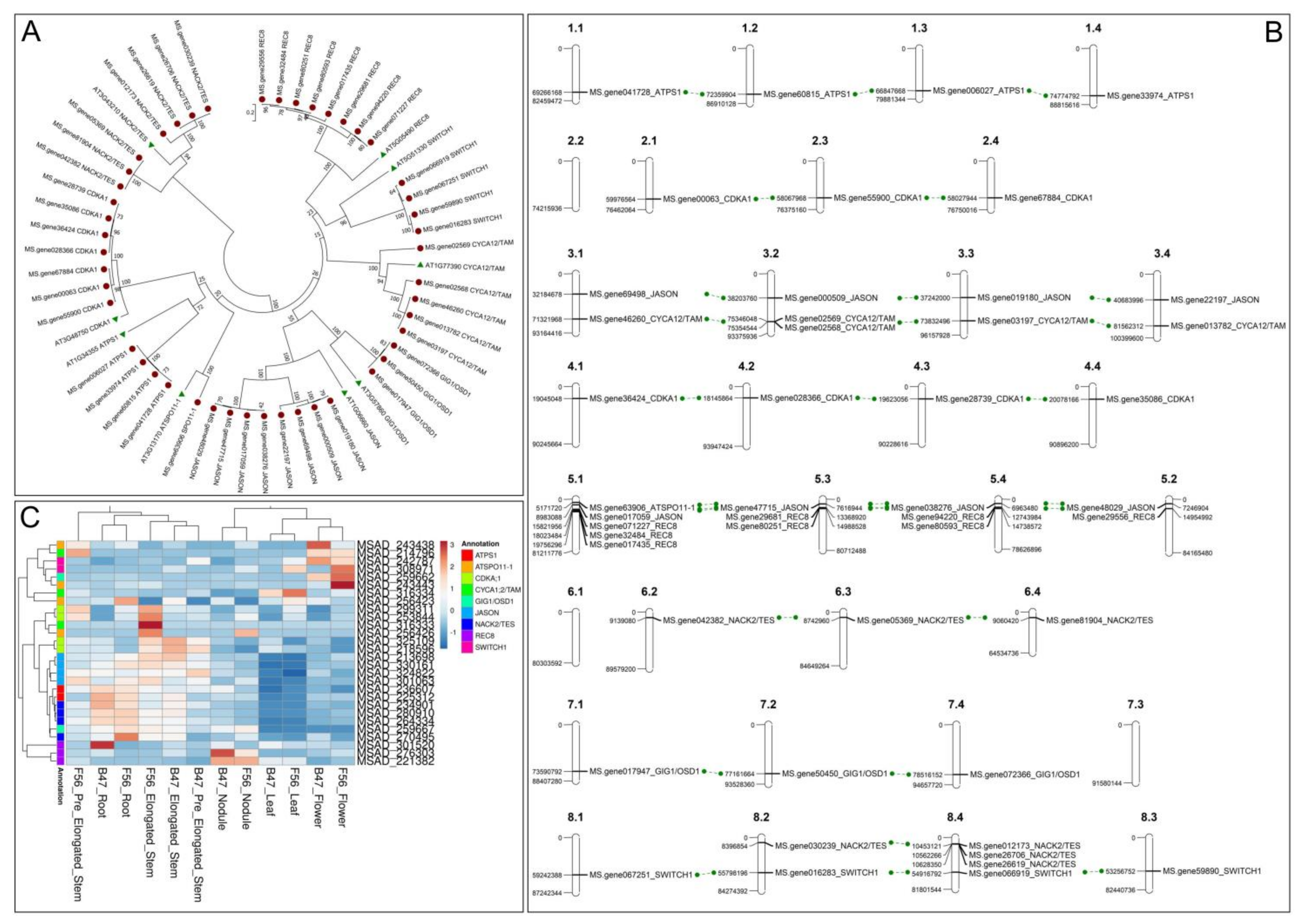
3.1. Forty Years of Genetic Resources and Genomic Studies in Medicago sativa
3.2. Identification of Candidate Genes through a Reverse Genetics Approach
4. Concluding Remarks and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Mendiburu, A.O.; Peloquin, S.J. Sexual polyploidization and depolyploidization: Some terminology and definitions. Theor. Appl. Genet. 1976, 48, 137–143. [Google Scholar]
- De Wet, J.M. Origins of polyploids. Basic Life Sci. 1980, 13, 3–15. [Google Scholar]
- Darlington, C. Chromosome Botany and the Origins of Cultivated Plants; Allen & Unwin: London, UK, 1973. [Google Scholar]
- Harlan, J.R.; de Wet, J.M.J. On Ö. Winge and a Prayer: The origins of polyploidy. Bot. Rev. 1975, 41, 361–390. [Google Scholar] [CrossRef]
- Lesins, K.A.; Lesins, I. Genus Medicago a Taxogenetic Study; Springer: Dordrecht, The Netherlands, 1979. [Google Scholar]
- Calderini, O.; Pupilli, F.; Cluster, P.D.; Mariani, A.; Arcioni, S. Cytological studies of the nucleolus organizing regions in the Medicago complex: Sativa-coerulea-falcata. Genome 1996, 39, 914–920. [Google Scholar] [CrossRef]
- Stanford, E.H.; Clement, W.M.; Bingham, E.T. Cytology and Evolution of the Medicago sativa-falcata Complex; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2015; pp. 87–101. [Google Scholar]
- Bingham, E.T. Maximizing heterozygosity in autopolyploids. Basic Life Sci. 1979, 13, 471–489. [Google Scholar]
- Hermsen, J.G.T. The potential of meiotic polyploidization in breading allogamous crops. Iowa State J. Res. 1984, 58, 435–448. [Google Scholar]
- Veronesi, F.; Mariani, A.; Bingham, E.T. Unreduced gametes in diploid Medicago and their importance in alfalfa breeding. Theor. Appl. Genet. 1986, 72, 37–41. [Google Scholar] [CrossRef]
- McCoy, T.J.; Bingham, E.T. Cytology and Cytogenetics of Alfalfa; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 1988; pp. 737–776. [Google Scholar]
- McCoy, T.J. Genome manipulation and molecular genetic analysis of alfalfa (Medicago sativa). In Gametes with Somatic Chromosome Number in the Evolution and Breeding of Polyploid Polysomic Species: Achievements and Perspectives; Tavoletti, S., Mariani, A., Eds.; University of Perugia: Perugia, Italy, 1992; pp. 55–59. [Google Scholar]
- Barcaccia, G.; Tavoletti, S.; Mariani, A.; Veronesi, F. Occurrence, inheritance and use of reproductive mutants in alfalfa improvement. Euphytica 2003, 133, 37–56. [Google Scholar] [CrossRef]
- McCoy, T.J.; Rowe, D.E. Single cross alfalfa (Medicago sativa L.) hybrids produced via 2n gametes and somatic chromosome doubling: Experimental and theoretical comparisons. Theor. Appl. Genet. 1986, 72, 80–83. [Google Scholar] [CrossRef]
- Barcaccia, G.; Albertini, E.; Luchin, M.; Veronesi, F. Progress in assembling a functional system of apomictic seed production in alfalfa. In Herbage Seed as a Key Factor for Improving Production and Environmental Quality; Falcinelli, M., Rosellini, D., Eds.; University of Perugia: Perugia, Italy, 1999; pp. 198–202. [Google Scholar]
- Vorsa, N.; Bingham, E.T. Cytology of 2n pollen formation in diploid alfalfa, Medicago sativa. Can. J. Genet. Cytol. 1979, 21, 525–530. [Google Scholar] [CrossRef]
- Tavoletti, S. Cytological mechanisms of 2n egg formation in a diploid genotype of Medicago sativa subsp. falcata. Euphytica 1994, 75, 1–8. [Google Scholar] [CrossRef]
- McCoy, T.J. The inheritance of 2n pollen formation in diploid alfalfa Medicago sativa. Can. J. Genet. Cytol. 1982, 24, 315–323. [Google Scholar] [CrossRef]
- Barcaccia, G.; Tavoletti, S.; Falcinelli, M.; Veronesi, F. Environmental influences on the frequency and viability of meiotic and apomeiotic cells of a diploid mutant of alfalfa. Crop Sci. 1997, 37, 70–76. [Google Scholar] [CrossRef]
- Peloquin, S.J. GChromosome Engineering with Meiotic Mutants. In Biotechnology and Ecology of Pollen; Mulcahy, D.L., Mulcahy, G.B., Ottaviano, E., Eds.; University of Massachusetts: Amherst, MA, USA, 1985; pp. 47–52. [Google Scholar]
- Kaul, M.L.H.; Murthy, T.G.K. Mutant genes affecting higher plant meiosis. Theor. Appl. Genet. 1985, 70, 449–466. [Google Scholar] [CrossRef]
- Clement, W.M.; Stanford, E.H. A mechanism for the production of tetraploid and pentaploid progeny from diploid × tetraploid crosses of alfalfa. Crop Sci. 1961, 1, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Tavoletti, S.; Mariani, A.; Veronesi, F. Cytological analysis of macro- and microsporogenesis of a diploid alfalfa clone producing male and female 2n gametes. Crop Sci. 1991, 31, 1258–1263. [Google Scholar] [CrossRef]
- McCoy, T.J.; Smith, L.Y. Genetics, cytology, and crossing behavior of an alfalfa (Medicago sativa) mutant resulting in failure of the postmeiotic cytokinesis. Can. J. Genet. Cytol. 1983, 25, 390–397. [Google Scholar] [CrossRef]
- Pfeiffer, T.W.; Bingham, E.T. Abnormal meiosis in alfalfa, Medicago sativa: Cytology of 2 N egg and 4 N pollen formation. Can. J. Genet. Cytol. 1983, 25, 107–112. [Google Scholar] [CrossRef]
- Barcaccia, G.; Mazzuccato, A.; Falcinelli, M.; Veronesi, F. Callose localization during meiotic and apomeiotic megasporo- genesis in alfalfa (Medicago ssp.). Caryologia 1996, 49, 45–56. [Google Scholar] [CrossRef]
- Tavoletti, S.; Bingham, E.T.; Yandell, B.S.; Veronesit, F.; Osborn, T.C. Half tetrad analysis in alfalfa using multiple restriction fragment length polymorphism markers. Proc. Natl. Acad. Sci. USA 1996, 93, 10918–10922. [Google Scholar] [CrossRef] [Green Version]
- Johnston, S.A.; Den Nijs, T.P.M.; Peloquin, S.J.; Hanneman, R.E. The significance of genic balance to endosperm development in interspecific crosses. Theor. Appl. Genet. 1980, 57, 5–9. [Google Scholar] [CrossRef]
- Calderini, O.; Mariani, A. Identification of meiotic mutants producing 2n pollen in the Medicago sativa complex. J. Genet. Breed. 1994, 48, 317–320. [Google Scholar]
- Comai, L. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Mayrose, I.; Zhan, S.H.; Rothfels, C.J.; Magnuson-Ford, K.; Barker, M.S.; Rieseberg, L.H.; Otto, S.P. Recently formed polyploid plants diversify at lower rates. Science 2011, 333, 1257. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.S.; Pires, J.C. Unreduced gametes: Meiotic mishap or evolutionary mechanism? Trends Genet. 2015, 31, 5–10. [Google Scholar] [CrossRef]
- Soltis, D.E.; Segovia-Salcedo, M.C.; Jordon-Thaden, I.; Majure, L.; Miles, N.M.; Mavrodiev, E.V.; Mei, W.; Cortez, M.B.; Soltis, P.S.; Gitzendanner, M.A.; et al. Are polyploids really evolutionary dead-ends (again)? A critical reappraisal of Mayrose. New Phytol. 2014, 202, 1105–1117. [Google Scholar] [CrossRef]
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity 2013, 110, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Cuypers, T.D.; Hogeweg, P. A Synergism between Adaptive Effects and Evolvability Drives Whole Genome Duplication to Fixation. PLoS Comput. Biol. 2014, 10, e1003547. [Google Scholar] [CrossRef] [Green Version]
- Mable, B.K.; Otto, S.P. Masking and purging mutations following EMS treatment in haploid, diploid and tetraploid yeast (Saccharomyces cerevisiae). Genet. Res. 2001, 77, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Stadler, L.J. Chromosome number and the mutation rate in Avena and Triticum. Proc. Natl. Acad. Sci. USA 1929, 15, 876–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.J. Plant Cytogenetics; CRC Press LLC: Boca Raton, FL, USA, 2003. [Google Scholar]
- Stupar, R.M.; Bhaskar, P.B.; Yandell, B.S.; Rensink, W.A.; Hart, A.L.; Ouyang, S.; Veilleux, R.E.; Busse, J.S.; Erhardt, R.J.; Buell, C.R.; et al. Phenotypic and transcriptomic changes associated with potato autopolyploidization. Genetics 2007, 176, 2055–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allario, T.; Brumos, J.; Colmenero-Flores, J.M.; Tadeo, F.; Froelicher, Y.; Talon, M.; Navarro, L.; Ollitrault, P.; Morillon, R. Large changes in anatomy and physiology between diploid Rangpur lime (Citrus limonia) and its autotetraploid are not associated with large changes in leaf gene expression. J. Exp. Bot. 2011, 62, 2507–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aversano, R.; Scarano, M.T.; Aronne, G.; Caruso, I.; D’Amelia, V.; De Micco, V.; Fasano, C.; Termolino, P.; Carputo, D. Genotype-specific changes associated to early synthesis of autotetraploids in wild potato species. Euphytica 2015, 202, 307–316. [Google Scholar] [CrossRef]
- Innes, L.A.; Denton, M.D.; Dundas, I.S.; Peck, D.M.; Humphries, A.W. The effect of ploidy number on vigor, productivity, and potential adaptation to climate change in annual Medicago species. Crop Sci. 2020, 61, 89–103. [Google Scholar] [CrossRef]
- Anssour, S.; Krügel, T.; Sharbel, T.F.; Saluz, H.P.; Bonaventure, G.; Baldwin, I.T. Phenotypic, genetic and genomic consequences of natural and synthetic polyploidization of Nicotiana attenuata and Nicotiana obtusifolia. Ann. Bot. 2009, 103, 1207–1217. [Google Scholar] [CrossRef] [Green Version]
- Riddle, N.C.; Jiang, H.; An, L.; Doerge, R.W.; Birchler, J.A. Gene expression analysis at the intersection of ploidy and hybridity in maize. Theor. Appl. Genet. 2010, 120, 341–353. [Google Scholar] [CrossRef]
- Ramanna, M.S.; Jacobsen, E. Relevance of sexual polyploidization for crop improvement—A review. Euphytica 2003, 133, 3–8. [Google Scholar] [CrossRef]
- Obajimi, A.O.; Bingham, E.T. Inbreeding cultivated alfalfa in one tetraploid-haploid-tetraploid cycle: Effects on morphology, fertility, and cytology. Crop Sci. 1973, 13, 36. [Google Scholar] [CrossRef]
- Hahn, M.A.; Lanz, T.; Fasel, D.; Müller-Schärer, H. Increased seed survival and seedling emergence in a polyploid plant invader. Am. J. Bot. 2013, 100, 1555–1561. [Google Scholar] [CrossRef] [Green Version]
- Rosellini, D.; Ferradini, N.; Allegrucci, S.; Capomaccio, S.; Zago, E.D.; Leonetti, P.; Balech, B.; Aversano, R.; Carputo, D.; Reale, L.; et al. Sexual polyploidization in Medicago sativa L.: Impact on the phenotype, gene transcription, and genome methylation. G3 Genes Genomes Genet. 2016, 6, 925–938. [Google Scholar]
- Bond, W.J.; Honig, M.; Maze, K.E. Seed size and seedling emergence: An allometric relationship and some ecological implications. Oecologia 1999, 120, 132–136. [Google Scholar] [CrossRef]
- Wang, J.; Tian, L.; Lee, H.-S.; Wei, N.E.; Jiang, H.; Watson, B.; Madlung, A.; Osborn, T.C.; Doerge, R.W.; Comai, L.; et al. Genomewide nonadditive gene regulation in Arabidopsis allotetraploids. Genetics 2006, 172, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Bingham, E.T. Maximizing heterozygosity in autotetraptoids. In Polyploidy: Biological Relevance; Lewis, W.H., Ed.; Plenum Press: New York, NY, USA, 1980; pp. 471–489. [Google Scholar]
- Bingham, E.T. Isolation of haploids of tetraploid alfalfa. Crop Sci. 1971, 11, 433–435. [Google Scholar] [CrossRef]
- Barcaccia, G.; Tavoletti, S.; Falcinelli, M.; Veronesi, F. Verification of the parthenogenetic capability of unreduced eggs in an alfalfa mutant by a progeny test based on morphological and molecular markers. Plant Breed. 1997, 116, 475–479. [Google Scholar] [CrossRef]
- Asker, S.; Jerling, L. Apomixis in Plants; CRC Press, Inc: Boca Raton, FL, USA, 1992; ISBN 9780849345456. [Google Scholar]
- Lucretti, S.; Doležel, J. Cell cycle synchronization, chromosome isolation, and flow-sorting in plants. In Methods in Plant Cell Biology; Galbraith, D.W., Bohnert, H., Bourque, D.P., Eds.; Academic Press: New York, NY, USA, 1995; pp. 61–83. [Google Scholar]
- Albertini, E. Investigation and manipolation of reproductive systems in Medicago spp. and Poa pratensis L. Ph.D. Thesis, University of Torino, Torin, Italy, 2000. [Google Scholar]
- Barcaccia, G.; Albertini, E.; Falcinelli, M. AFLP fingerprinting in Pelargonium peltatum: Its development and potential in cultivar identification. J. Hortic. Sci. Biotechnol. 1999, 74, 243–250. [Google Scholar] [CrossRef]
- Barcaccia, G.; Palumbo, F.; Sgorbati, S.; Albertini, E.; Pupilli, F. A reappraisal of the evolutionary and developmental pathway of apomixis and its genetic control in angiosperms. Genes 2020, 11, 859. [Google Scholar] [CrossRef]
- Oleszczuk, S.; Grzechnik, N.; Mason, A.S.; Zimny, J. Heritability of meiotic restitution and fertility restoration in haploid triticale. Plant Cell Rep. 2019, 38, 1515–1525. [Google Scholar] [CrossRef] [Green Version]
- Veronesi, F.; Mariani, A.; Tavoletti, S. Screening for 2N gamete producers in diploid species of genus Medicago. Genet. Agrar. 1988, 42, 187–199. [Google Scholar]
- Brownfield, L.; Köhler, C. Unreduced gamete formation in plants: Mechanisms and prospects. J. Exp. Bot. 2011, 62, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Lesins, K.A.; Lesins, I. Genus Medicago (Leguminosae). In A Taxogenetic Study; Junk, W., Ed.; The Hague Publishers: Boston, MA, USA, 1979; pp. 10–58. [Google Scholar]
- Diwan, N.; Bouton, J.H.; Kochert, G.; Cregan, P.B. Mapping of simple sequence repeat (SSR) DNA markers in diploid and tetraploid alfalfa. Theor. Appl. Genet. 2000, 101, 165–172. [Google Scholar] [CrossRef]
- Brouwer, D.J.; Osborn, T.C. A molecular marker linkage map of tetraploid alfalfa (Medicago sativa L.). Theor. Appl. Genet. 1999, 99, 1194–1200. [Google Scholar] [CrossRef]
- Kiss, G.B.; Csanádi, G.; Kálmán, K.; Kaló, P.; Ökrész, L. Construction of a basic genetic map for alfalfa using RFLP, RAPD, isozyme and morphological markers. Mol. Gen. Genet. 1993, 238, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Echt, C.S.; Kidwell, K.K.; Knapp, S.J.; Osborn, T.C.; McCoy, T.J. Linkage mapping in diploid alfalfa (Medicago sativa). Genome 1994, 37, 61–71. [Google Scholar] [CrossRef]
- Tavoletti, S.; Pesaresi, P.; Barcaccia, G.; Albertini, E.; Veronesi, F. Mapping the jp (jumbo pollen) gene and QTLs involved in multinucleate microspore formation in diploid alfalfa. Theor. Appl. Genet. 2000, 101, 372–378. [Google Scholar] [CrossRef]
- Barcaccia, G.; Albertini, E.; Rosellini, D.; Tavoletti, S.; Veronesi, F. Inheritance and mapping of 2n-egg production in diploid alfalfa. Genome 2000, 43, 528–537. [Google Scholar] [CrossRef]
- Barcaccia, G.; Varotto, S.; Meneghetti, S.; Albertini, E.; Porceddu, A.; Parrini, P.; Lucchin, M. Analysis of gene expression during flowering in apomeiotic mutants of Medicago spp.: Cloning of ESTs and candidate genes for 2n eggs. Sex. Plant Reprod. 2001, 14, 233–238. [Google Scholar] [CrossRef]
- Zamariola, L.; Tiang, C.L.; De Storme, N.; Pawlowski, W.; Geelen, D. Chromosome segregation in plant meiosis. Front. Plant Sci. 2014, 5, 279. [Google Scholar] [CrossRef] [Green Version]
- Silkova, O.G.; Loginova, D.B. Sister chromatid separation and monopolar spindle organization in the first meiosis as two mechanisms of unreduced gametes formation in wheat–rye hybrids. Plant Reprod. 2016, 29, 199–213. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kang, X.; Zhu, Q. Variation in pollen formation and its cytological mechanism in an allotriploid white poplar. Tree Genet. Genomes 2010, 6, 281–290. [Google Scholar] [CrossRef]
- Mariani, A.; Campanoni, P.; Gianì, S.; Breviario, D. Meiotic mutants of Medicago sativa show altered levels of α- and β-tubulin. Genome 2000, 43, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Barcaccia, G.; Varotto, S.; Albertini, E.; Parrini, P.; Lucchin, M. MOB (Mps-one-binder), a multi gene family that may shed light on apomeiosis in alfalfa mutants. In Proceedings of the XVII Int Congress on Sexual Plant Reproduction, Lublin, Poland, 9–13 July 2002; p. 30. [Google Scholar]
- Luca, F.C.; Winey, M. Mob1, an essential yeast gene required for completion of mitosis and maintenance of ploidy. Mol. Biol. Cell 1998, 9, 29–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luca, F.C.; Mody, M.; Kurischko, C.; Roof, D.M.; Giddings, T.H.; Winey, M. Saccharomyces cerevisiae Mob1p Is Required for Cytokinesis and Mitotic Exit. Mol. Cell. Biol. 2001, 21, 6972–6983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.; Hao, Y.; Yang, X. Molecular characterization of human homologs of yeast MOB1. Int. J. Cancer 2010, 126, 2079–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citterio, S.; Albertini, E.; Varotto, S.; Feltrin, E.; Soattin, M.; Marconi, G.; Sgorbati, S.; Lucchin, M.; Barcaccia, G. Alfalfa Mob1-like genes are expressed in reproductive organs during meiosis and gametogenesis. Plant Mol. Biol. 2005, 58, 789–807. [Google Scholar] [CrossRef] [PubMed]
- Citterio, S.; Piatti, S.; Albertini, E.; Aina, R.; Varotto, S.; Barcaccia, G. Alfalfa Mob1-like proteins are involved in cell proliferation and are localized in the cell division plane during cytokinesis. Exp. Cell Res. 2006, 312, 1050–1064. [Google Scholar] [CrossRef]
- Yang, S.S.; Xu, W.W.; Tesfaye, M.; Lamb, J.F.S.; Jung, H.G.; Samac, D.A.; Vance, C.P.; Gronwald, J.W. Single-Feature Polymorphism Discovery in the Transcriptome of Tetraploid Alfalfa. Plant Genome 2009, 2, 224–232. [Google Scholar] [CrossRef]
- Li, X.; Acharya, A.; Farmer, A.D.; Crow, J.A.; Bharti, A.K.; Kramer, R.S.; Wei, Y.; Han, Y.; Gou, J.; May, G.D.; et al. Prevalence of single nucleotide polymorphism among 27 diverse alfalfa genotypes as assessed by transcriptome sequencing. BMC Genom. 2012, 13. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Han, Y.; Wei, Y.; Acharya, A.; Farmer, A.D.; Ho, J.; Monteros, M.J.; Brummer, E.C. Development of an alfalfa SNP array and its use to evaluate patterns of population structure and linkage disequilibrium. PLoS ONE 2014, 9, e84329. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Jiang, L.; Chen, Y.; Shu, Y.; Bai, Y.; Guo, C. Deep-sequencing transcriptome analysis of field-grown Medicago sativa L. crown buds acclimated to freezing stress. Funct. Integr. Genomics 2016, 16, 495–511. [Google Scholar] [CrossRef]
- Zeng, N.; Yang, Z.; Zhang, Z.; Hu, L.; Chen, L. Comparative transcriptome combined with proteome analyses revealed key factors involved in alfalfa (Medicago sativa) response to waterlogging stress. Int. J. Mol. Sci. 2019, 20, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Xu, Y.; Hettenhausen, C.; Lu, C.; Shen, G.; Zhang, C.; Li, J.; Song, J.; Lin, H.; Wu, J. Comparative analysis of alfalfa (Medicago sativa L.) leaf transcriptomes reveals genotype-specific salt tolerance mechanisms. BMC Plant Biol. 2018, 18, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Xiong, C.; Yan, L.; Zhang, Z.; Ma, L.; Wang, Y.; Liu, Y.; Liu, Z. Transcriptome analyses reveal candidate genes potentially involved in al stress response in alfalfa. Front. Plant Sci. 2017, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, P.; Mowery, J.; Eisenback, J.D.; Shao, J.; Nemchinov, L.G. Cellular and transcriptional responses of resistant and susceptible cultivars of alfalfa to the root lesion nematode, Pratylenchus penetrans. Front. Plant Sci. 2019, 10, 971. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Shi, Y.; Cheng, N.; Du, H.; Fan, W.; Wang, C. De novo characterization of fall dormant and nondormant alfalfa (Medicago sativa L) leaf transcriptome and identification of candidate genes related to fall dormancy. PLoS ONE 2015, 10, 1371. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.S.; Tu, Z.J.; Cheung, F.; Xu, W.W.; Lamb, J.A.F.S.; Jung, H.J.G.; Vance, C.P.; Gronwald, J.W. Using RNA-Seq for gene identification, polymorphism detection and transcript profiling in two alfalfa genotypes with divergent cell wall composition in stems. BMC Genom. 2011, 12, 199. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, J.A.; Fu, F.; Bucciarelli, B.; Yang, S.S.; Samac, D.A.; Lamb, J.A.F.S.; Monteros, M.J.; Graham, M.A.; Gronwald, J.W.; Krom, N.; et al. The Medicago sativa gene index 1.2: A web-accessible gene expression atlas for investigating expression differences between Medicago sativa subspecies. BMC Genom. 2015, 16, 502. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.; Liu, Y.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 2494. [Google Scholar] [CrossRef]
- Pillet, J.; Yu, H.W.; Chambers, A.H.; Whitaker, V.M.; Folta, K.M. Identification of candidate flavonoid pathway genes using transcriptome correlation network analysis in ripe strawberry (Fragaria × ananassa) fruits. J. Exp. Bot. 2015, 66, 4455–4467. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, Y.; He, X.; Du, J.; Zhang, R.; Ma, Y.; Hu, X.; Zhang, Z.; Chen, Q.; Wan, X. Identification of MYB Transcription Factors Regulating Theanine Biosynthesis in Tea Plant Using Omics-Based Gene Coexpression Analysis. J. Agric. Food Chem. 2020, 68, 918–926. [Google Scholar] [CrossRef]
- Barcaccia, G.; Palumbo, F.; Scariolo, F.; Vannozzi, A.; Borin, M.; Bona, S. Potentials and challenges of genomics for breeding Cannabis Cultivars. Front. Plant Sci. 2020, 11, 573299. [Google Scholar] [CrossRef]
- Yang, C.; Hamamura, Y.; Sofroni, K.; Böwer, F.; Stolze, S.C.; Nakagami, H.; Schnittger, A. SWITCH 1/DYAD is a WINGS APART-LIKE antagonist that maintains sister chromatid cohesion in meiosis. Nat. Commun. 2019, 10, 1755. [Google Scholar] [CrossRef] [Green Version]
- Agashe, B.; Prasad, C.K.; Siddiqi, I. Identification and analysis of DYAD: A gene required for meiotic chromosome organisation and female meiotic progression in Arabidopsis. Development 2002, 129, 3935–3943. [Google Scholar] [CrossRef]
- Ravi, M.; Marimuthu, M.P.A.; Siddiqi, I. Gamete formation without meiosis in Arabidopsis. Nature 2008, 451, 1121–1124. [Google Scholar] [CrossRef]
- Grelon, M.; Vezon, D.; Gendrot, G.; Pelletier, G. AtSPO11-1 is necessary for efficient meiotic recombination in plants. EMBO J. 2001, 20, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Chelysheva, L.; Diallo, S.; Vezon, D.; Gendrot, G.; Vrielynck, N.; Belcram, K.; Rocques, N.; Márquez-Lema, A.; Bhatt, A.M.; Horlow, C.; et al. AtREC8 and AtSCC3 are essential to the monopolar orientation of the kinetochores during meiosis. J. Cell Sci. 2005, 118, 4621–4632. [Google Scholar] [CrossRef] [Green Version]
- D’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Simon, M.; Jenczewski, E.; Mercier, R. Mutations in AtPS1 (Arabidopsis thaliana Parallel Spindle 1) lead to the production of diploid pollen grains. PLoS Genet. 2008, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. The Arabidopsis mutant jason produces unreduced first division restitution male gametes through a parallel/ fused spindle mechanism in meiosis II. Plant Physiol. 2011, 155, 1403–1415. [Google Scholar] [CrossRef] [Green Version]
- Erilova, A.; Brownfield, L.; Exner, V.; Rosa, M.; Twell, D.; Scheid, O.M.; Hennig, L.; Köhler, C. Imprinting of the Polycomb group gene MEDEA serves as a ploidy sensor in Arabidopsis. PLoS Genet. 2009, 5, e1000663. [Google Scholar] [CrossRef] [Green Version]
- D’Erfurth, I.; Cromer, L.; Jolivet, S.; Girard, C.; Horlow, C.; Sun, Y.; To, J.P.C.; Berchowitz, L.E.; Copenhaver, G.P.; Mercier, R. The CYCLIN-A CYCA1;2/TAM is required for the meiosis I to meiosis II transition and cooperates with OSD1 for the prophase to first meiotic division transition. PLoS Genet. 2010, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jha, A.K.; Chen, R.; Doonan, J.H.; Yang, M. Polyploidy-associated genomic instability in Arabidopsis thaliana. Genesis 2010, 48, 254–263. [Google Scholar] [CrossRef]
- Dissmeyer, N.; Nowack, M.K.; Pusch, S.; Stals, H.; Inzé, D.; Grini, P.E.; Schnittger, A. T-loop phosphorylation of Arabidopsis CDKA;1 is required for its function and can be partially substituted by an aspartate residue. Plant Cell 2007, 19, 972–985. [Google Scholar] [CrossRef] [Green Version]
- D’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Mercier, R. Turning meiosis into mitosis. PLoS Biol. 2009, 7, e1000124. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.Y.; Spielman, M.; Coles, J.P.; Li, Y.; Ghelani, S.; Bourdon, V.; Brown, R.C.; Lemmon, B.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE encodes a kinesin required for male meiotic cytokinesis in Arabidopsis. Plant J. 2003, 34, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Pupilli, F.; Barcaccia, G. Cloning plants by seeds: Inheritance models and candidate genes to increase fundamental knowledge for engineering apomixis in sexual crops. J. Biotechnol. 2012, 159, 291–311. [Google Scholar] [CrossRef]
- Albertini, E.; Barcaccia, G.; Carman, J.G.; Pupilli, F. Did apomixis evolve from sex or was it the other way around? J. Exp. Bot. 2019, 70, 2951–2964. [Google Scholar] [CrossRef]
- Podio, M.; Cáceres, M.E.; Samoluk, S.S.; Seijo, J.G.; Pessino, S.C.; Ortiz, J.P.A.; Pupilli, F. A methylation status analysis of the apomixis-specific region in Paspalum spp. suggests an epigenetic control of parthenogenesis. J. Exp. Bot. 2014, 65, 6411–6424. [Google Scholar] [CrossRef] [Green Version]
- Stelly, D.; Peloquin, S.J.; Palmer, R.G.; Crane, C.F. Mayer’s hemalum-methyl salicylate: A stain-clearing technique for observations within whole ovules. Stain Technol. 1984, 59, 155–161. [Google Scholar] [CrossRef]
- Barcaccia, G. Development, comparability and potential applications of RAPD markers in the genus Medicago. J. Genet. Breed. 1994, 48, 161–168. [Google Scholar]
- Barcaccia, G.; Mazzucato, A.; Albertini, E.; Zethof, J.; Gerats, A.; Pezzotti, M.; Falcinelli, M. Inheritance of parthenogenesis in Poa pratensis L.: Auxin test and AFLP linkage analyses support monogenic control. Theor. Appl. Genet. 1998, 97, 74–82. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BC1 Plant Code | Nuclear DNA (pg) | Ploidy Level | Genetic Similarity | Observed Ovules | Diplospory (%) | Nucleolus Diameter (μm) | Self-Fertility | Mean Cross-Fertility | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| With TNE | With Pollinator | Meiotic | Diplosporic | With 2x | With 4x | ||||||
| A2 | 1.471 | 2x | 0.992 | 0.137 | 98 | 15.31 | 1.53 ± 0.03 | 2.58 ± 0.13 | 0.009 | 0.190 | 0.036 |
| B1 | 2.972 | 4x | 0.608 | 0.451 | 132 | 19.70 | 1.56 ± 0.08 | 2.71 ± 0.11 | 0 | 0.008 | 0.935 |
| B2 | 2.982 | 4x | 0.549 | 0.510 | 203 | 21.67 | 1.45 ± 0.07 | 2.26 ± 0.12 | 0 | 0.006 | 0.848 |
| B3 | 2.855 | 4x | 0.431 | 0.627 | 164 | 0 | − | − | 0.022 | 0.019 | 1.466 |
| B4 | 2.689 | 4x | 0.686 | 0.373 | 110 | 0 | − | − | 0.018 | 0.014 | 1.523 |
| B5 | 2.724 | 4x | 0.725 | 0.333 | 108 | 13.89 | 1.31 ± 0.08 | 2.37 ± 0.15 | 0 | 0.041 | 0.530 |
| B6 | 3.110 | 4x | 0.588 | 0.471 | 94 | 2.13 | − | − | 0 | 0.019 | 0.796 |
| B8 | 3.195 | 4x | 0.667 | 0.392 | 83 | 40.96 | 1.38 ± 0.09 | 2.26 ± 0.10 | 0 | 0.018 | 0.645 |
| C3 | 2.780 | 4x | 0.529 | 0.529 | 151 | 15.23 | 1.81 ± 0.07 | 2.62 ± 0.19 | 0.016 | 0 | 0.964 |
| C6 | 2.791 | 4x | 0.471 | 0.588 | 119 | 5.04 | 1.58 ± 0.05 | 2.35 ± 0.10 | 0 | 0 | 0.511 |
| C7 | 3.132 | 4x | 0.511 | 0.549 | 112 | 6.25 | 1.44 ± 0.06 | 2.02 ± 0.05 | 0 | 0.010 | 0.677 |
| C9 | 3.165 | 4x | 0.509 | 0.549 | 96 | 3.13 | − | − | 0 | 0.024 | 1.233 |
| D1 | 3.088 | 4x | 0.618 | 0.400 | 208 | 19.23 | 1.55 ± 0.05 | 2.44 ± 0.11 | 0 | 0.019 | 1.446 |
| D2 | 2.869 | 4x | 0.618 | 0.364 | 98 | 0 | − | − | 0.089 | 0 | 2.071 |
| D3 | 1.534 | 2x | 0.909 | 0.145 | 95 | 57.89 | 1.39 ± 0.10 | 2.38 ± 0.09 | 0 | 0 | 0.538 |
| E1 | 3.077 | 4x | 0.709 | 0.309 | 137 | 17.52 | 1.53 ± 0.04 | 2.57 ± 0.13 | 0 | 0 | 1.479 |
| F1 | 2.986 | 4x | 0.618 | 0.364 | 123 | 2.44 | 1.60 ± 0.09 | 1.93 ± 0.07 | 0.021 | 0.011 | 0.401 |
| F2 | 1.443 | 2x | 0.891 | 0.164 | 54 | 7.41 | 1.52 ± 0.06 | 2.29 ± 0.14 | 0 | 0.720 | 0.022 |
| DTA plants | 2.961 | 4x | 0.589 | 0.454 | 129 | 17.72 | 1.51 ± 0.07 | 2.40 ± 0.12 | 0.007 | 0.013 | 1.035 |
| TAIR ID | Gene Name—TAIR | Type | Defect Type | Genomic Data | Transcriptomic Data | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene Locus ID | Location | Bit | E-Value | Transcript | Bit | E-Value | ||||
| AT5G51330 | DYAD, SWI1, SWITCH1 | FDR | ♀ | MS.gene066919 | chr8.4 | 389 | 5 × 10−124 | MSAD_308971, MSAD_242787 | 1481 | 0.0 |
| MS.gene59890 | chr8.3 | 387 | 3 × 10−123 | 1279 | 0.0 | |||||
| MS.gene016283 | chr8.2 | 387 | 3 × 10−123 | 1279 | 0.0 | |||||
| MS.gene067251 | chr8.1 | 370 | 1 × 10−116 | 1438 | 0.0 | |||||
| AT1G34355 | ATPS1, PARALLEL SPINDLE 1, PS1 | FDR | ♂ | MS.gene006027 | chr1.3 | 220 | 1 × 10−57 | MSAD_236607, MSAD_225312 | 2228 | 0.0 |
| MS.gene041728 | chr1.1 | 214 | 1 × 10−55 | 2414 | 0.0 | |||||
| MS.gene33974 | chr1.4 | 214 | 2 × 10−55 | 2410 | 0.0 | |||||
| MS.gene60815 | chr1.2 | 209 | 9 × 10−54 | 2357 | 0.0 | |||||
| AT1G06660 | JASON | FDR | ♂ | MS.gene019180 | chr3.3 | 254 | 1 × 10−77 | MSAD_324822, MSAD_301063 | 915 | 0.0 |
| MS.gene000509 | chr3.2 | 249 | 9 × 10−76 | 917 | 0.0 | |||||
| MS.gene69498 | chr3.1 | 246 | 1 × 10−74 | 915 | 0.0 | |||||
| MS.gene22197 | chr3.4 | 241 | 8 × 10−73 | 837 | 0.0 | |||||
| MS.gene038276 | chr5.4 | 244 | 2 × 10−73 | MSAD_213698, MSAD_330161 | 974 | 0.0 | ||||
| MS.gene47715 | chr5.3 | 243 | 5 × 10−73 | 973 | 0.0 | |||||
| MS.gene48029 | chr5.2 | 242 | 1 × 10−72 | 975 | 0.0 | |||||
| MS.gene017059 | chr5.1 | 241 | 4 × 10−72 | 964 | 0.0 | |||||
| AT3G13170 | ATSPO11-1, SPO11-1 | FDR | ♂/♀ | MS.gene63906 | chr5.1 | 520 | 0.0 | MSAD_256423, MSAD_243443, MSAD_256426, MSAD_243438 | 746 | 0.0 |
| AT5G05490 | ATREC8, DETERMINATE INFERTILE 1, DIF1, REC8, SYN1, SYNAPTIC 1 | FDR | ♂/♀ | MS.gene29556 | chr5.2 | 364 | 2 × 10−116 | MSAD_301520 | 1079 | 0.0 |
| MS.gene32484 | chr5.1 | 363 | 7 × 10−116 | 1079 | 0.0 | |||||
| MS.gene80251 | chr5.3 | 338 | 4 × 10−106 | 1196 | 0.0 | |||||
| MS.gene80593 | chr5.4 | 330 | 3 × 10−103 | 1165 | 0.0 | |||||
| MS.gene017435 | chr5.1 | 326 | 1 × 10−101 | 1042 | 0.0 | |||||
| MS.gene94220 | chr5.4 | 335 | 4 × 10−105 | MSAD_276303, MSAD_221382 | 1156 | 0.0 | ||||
| MS.gene071227 | chr5.1 | 334 | 9 × 10−105 | 1158 | 0.0 | |||||
| MS.gene29681 | chr5.3 | 333 | 2 × 10−104 | 1161 | 0.0 | |||||
| AT3G48750 | CDKA;1, CELL DIVISION CONTROL 2 | SDR | ♂ | MS.gene35086 | chr4.4 | 520 | 0.0 | MSAD_325109, MSAD_253844 | 610 | 0.0 |
| MS.gene28739 | chr4.3 | 520 | 0.0 | 610 | 0.0 | |||||
| MS.gene36424 | chr4.1 | 520 | 0.0 | 610 | 0.0 | |||||
| MS.gene028366 | chr4.2 | 518 | 0.0 | 604 | 0.0 | |||||
| MS.gene67884 | chr2.4 | 520 | 0.0 | MSAD_299311, MSAD_218596 | 610 | 0.0 | ||||
| MS.gene55900 | chr2.3 | 520 | 0.0 | 610 | 0.0 | |||||
| MS.gene00063 | chr2.1 | 520 | 0.0 | 610 | 0.0 | |||||
| AT1G77390 | CYCA1;2, TAM, TARDY ASYNCHRONOUS MEIOSIS | SDR | ♂/♀ | MS.gene46260 | chr3.1 | 421 | 5 × 10−142 | MSAD_316334 | 987 | 0.0 |
| MS.gene013782 | chr3.4 | 421 | 5 × 10−142 | 994 | 0.0 | |||||
| MS.gene02568 | chr3.2 | 420 | 2 × 10−141 | 1026 | 0.0 | |||||
| MS.gene03197 | chr3.3 | 400 | 4 × 10−134 | 986 | 0.0 | |||||
| MS.gene02569 | chr3.2 | 386 | 3 × 10−128 | MSAD_316333, MSAD_214796 | 975 | 0.0 | ||||
| AT3G57860 | GIG1, OMISSION OF SECOND DIVISION, OSD1 | SDR | ♂/♀ | MS.gene017947 | chr7.1 | 119 | 3 × 10−31 | MSAD_259667, MSAD_259662 | 476 | 2 × 10−171 |
| MS.gene072366 | chr7.4 | 115 | 5 × 10−30 | 478 | 1 × 10−171 | |||||
| MS.gene50450 | chr7.2 | 115 | 5 × 10−30 | 478 | 1 × 10−171 | |||||
| AT3G43210 | ARABIDOPSIS NPK1-ACTIVATING KINESIN 2, ATNACK2, NACK2, TES, TETRASPORE, STUD | SDR | ♂ | MS.gene26619 | chr8.4 | 1160 | 0.0 | MSAD_280910, MSAD_270495 | 1908 | 0.0 |
| MS.gene26706 | chr8.4 | 1158 | 0.0 | 1909 | 0.0 | |||||
| MS.gene030239 | chr8.2 | 1157 | 0.0 | 1966 | 0.0 | |||||
| MS.gene012173 | chr8.4 | 825 | 0.0 | 1483 | 0.0 | |||||
| MS.gene042382 | chr6.2 | 1054 | 0.0 | MSAD_264334, MSAD_234901 | 1995 | 0.0 | ||||
| MS.gene81904 | chr6.4 | 1052 | 0.0 | 2004 | 0.0 | |||||
| MS.gene05369 | chr6.3 | 1050 | 0.0 | 1990 | 0.0 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palumbo, F.; Pasquali, E.; Albertini, E.; Barcaccia, G. A Review of Unreduced Gametes and Neopolyploids in Alfalfa: How to Fill the Gap between Well-Established Meiotic Mutants and Next-Generation Genomic Resources. Plants 2021, 10, 999. https://doi.org/10.3390/plants10050999
Palumbo F, Pasquali E, Albertini E, Barcaccia G. A Review of Unreduced Gametes and Neopolyploids in Alfalfa: How to Fill the Gap between Well-Established Meiotic Mutants and Next-Generation Genomic Resources. Plants. 2021; 10(5):999. https://doi.org/10.3390/plants10050999
Chicago/Turabian StylePalumbo, Fabio, Elisa Pasquali, Emidio Albertini, and Gianni Barcaccia. 2021. "A Review of Unreduced Gametes and Neopolyploids in Alfalfa: How to Fill the Gap between Well-Established Meiotic Mutants and Next-Generation Genomic Resources" Plants 10, no. 5: 999. https://doi.org/10.3390/plants10050999
APA StylePalumbo, F., Pasquali, E., Albertini, E., & Barcaccia, G. (2021). A Review of Unreduced Gametes and Neopolyploids in Alfalfa: How to Fill the Gap between Well-Established Meiotic Mutants and Next-Generation Genomic Resources. Plants, 10(5), 999. https://doi.org/10.3390/plants10050999