1. Introduction
Invasive species are non-indigenous species that are introduced outside their native ranges either deliberately or inadvertently and can threaten native biodiversity, alter the structure and function of ecosystems, disrupt natural and agricultural landscapes, and result in large-scale economic losses [
1,
2,
3,
4]. Invasive species alter the dynamics of plant communities by reducing the amounts of nutrients, water, and space available to native species and by changing the soil chemistry, hydrological pattern, and moisture-holding capacity in the region of invasion [
5,
6,
7,
8,
9].
Climate change exacerbates threats to, and losses of, biodiversity through multiple mechanisms, including reductions in climatic barriers and the facilitation of the spread of invasive species [
5,
6]. Therefore, there is great concern regarding the introduction, establishment, and naturalization of invasive species as a result of climate change and the consequent problems caused by invasive species in natural ecosystems worldwide [
10,
11]. Over the last century, the global temperature has increased by 0.78 °C, and it is predicted to increase by 2.6 °C to 4.8 °C by 2100 [
12]. The rate of climate change in South Korea is projected to be higher than that of the rest of the world [
13]. In South Korea, the average temperature has increased 1.8 °C over the last 100 years, and it is predicted to increase 1.75 °C temperature by 2050, 2.35 °C by 2070, and 5.7 °C by 2100 compared to the 19881~2005 average under the representative concentration pathway (RCP) 8.5 scenario [
13]. Thus, various environmental problems, including the spread of invasive species, are expected to be particularly serious in South Korea.
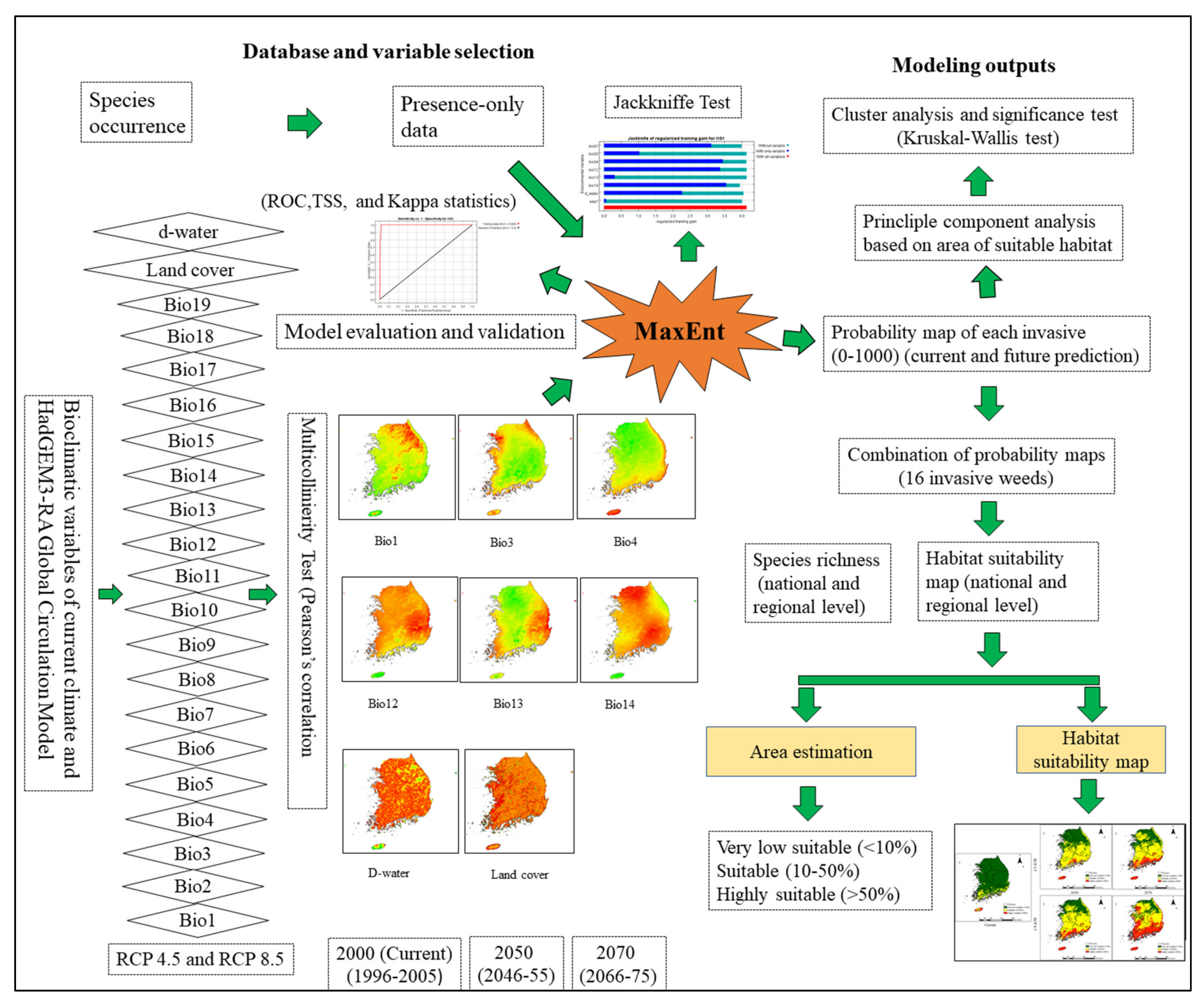
Predicting the potential distributions of invasive species under current and future climate change conditions will help prioritize the species and locations to target for early detection and will help leaders adopt the best preventive measures [
14]. Hence, there is a pressing need to develop reliable methods for predicting invasions ahead of time so that surveillance or management policies can be adjusted to reduce the risk of the establishment, spread, and overall expansion of invasive species [
15]. Species distribution models (SDMs) are considered the most reliable technique for simulating the range of suitable habitats of native and invasive species in response to climatic and environmental variables [
9,
16]. More recently, SDMs have been widely used in the fields of ecology, conservation biology, biogeography, and natural resource management [
11,
16,
17,
18,
19,
20]. Among the various available SDMs, the maximum entropy (MaxEnt) model is a frequently used machine learning technique that can obtain high predictive performance with a limited species presence and environmental variable data [
21,
22,
23,
24]. The MaxEnt model is well known for modeling invasive species because it only uses species presence data; absence data for invasive species may not always be reliable because the ranges of the species may be expanding and not yet have reached equilibrium, which may lead to incorrect interpretation [
25].
Among the various types of invasive species, the types of plants that are considered invasive weeds include small seasonal herbs to intermediate perennial shrubs growing on open and degraded lands, transportation corridors, riversides, and seashores [
11,
26,
27,
28]. Anthropogenic activities, such as the development, maintenance, and expansion of roads and railway connections, trade and tourism, and natural processes that occur via air, water, and wild animals, are known to be major vectors for the dispersal of invasive weeds across the world, including in and to South Korea [
6,
11,
29]. In South Korea, crop fields, orchards, pastures, and forests are at a high risk of invasion by invasive weeds [
27,
30,
31,
32,
33,
34]. Altogether, 320 taxa of invasive and alien plants were listed in South Korea in 2016, of which more than 95% are invasive weeds. This includes the 16 ecologically most disruptive weeds, such as
Rumex acetosella,
Paspalum distichum, and
Conyza canadensis [
28,
30,
31]. Invasive alien species have been reported to cause economic damage of 22.6 billion Korean Won (KRW; approximately 19.6 million USD as of July 2021) and the government of South Korea invests approximately 5 billion KRW (approximately 4.3 million USD as of July 2021) per year in the control and management of invasive species [
35].
To maintain the control and management of invasive weeds, regular studies are required to understand the ecology and distribution of these invasions, as well as their impacts on agriculture and forestry. Although many ecological studies have been carried out to investigate invasive species [
27,
28,
31,
32,
33,
36], very few studies have addressed plant invasions under climate change in South Korea [
24,
37,
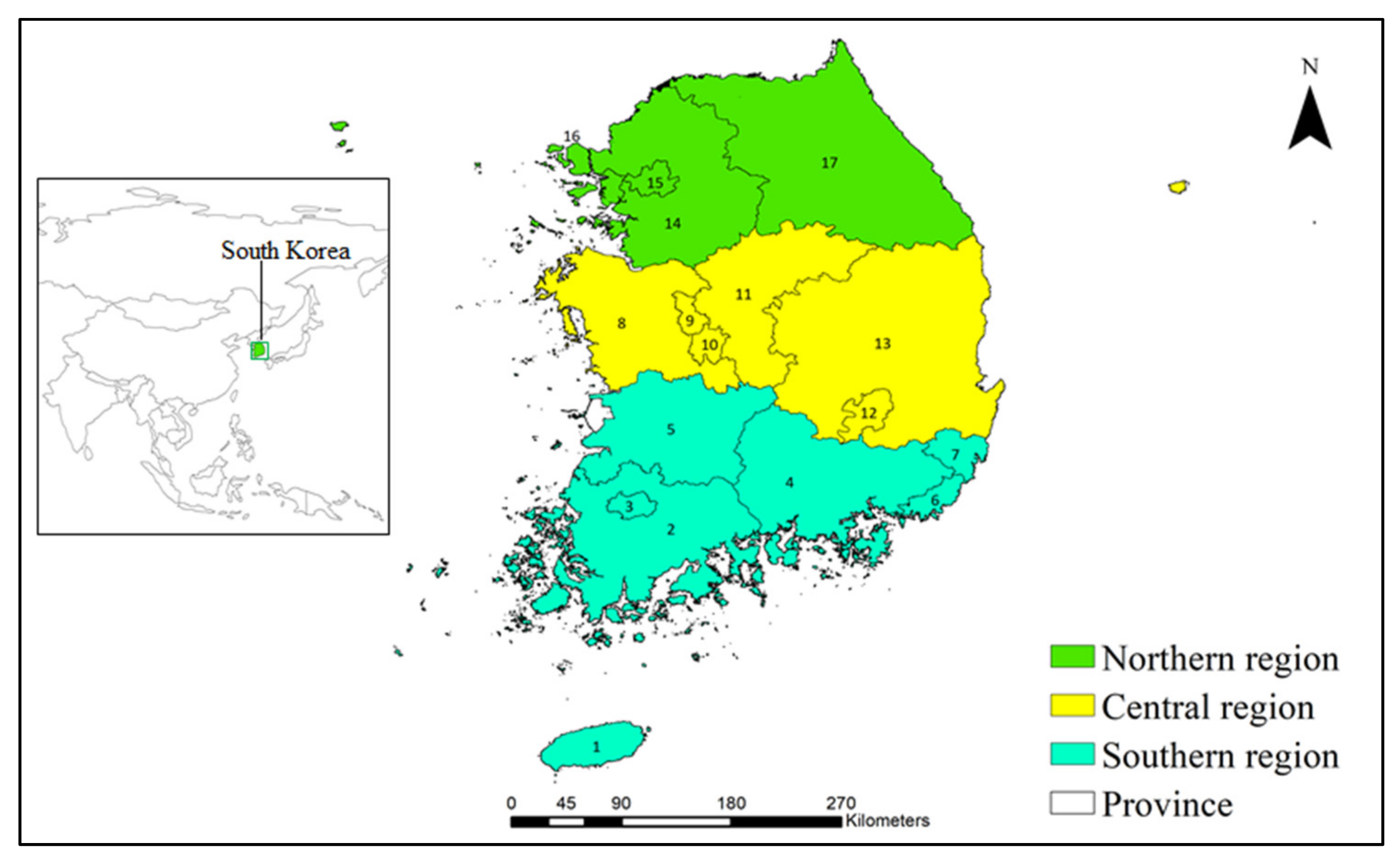
38]. The southern region of South Korea, including Jeju Province, Jeollanam Province, Gyeongsangnam Province, Busan City, and Ulsan City, is recognized as a high-risk region for the introduction and establishment of tropical and subtropical invasive weeds [
30,
31]. It is essential to identify areas that are potentially at risk of invasion under future climate change in the central and northern regions of the country. Therefore, this study was designed to assess the habitat suitability of the 16 most disruptive weeds [
31] in agricultural and natural ecosystems that are mainly distributed in the southern region of South Korea, including Jeju Island, under projected bioclimatic scenarios (RCP 4.5 and RCP 8.5) and land cover changes. Although 320 invasive weeds have been recorded in South Korea, we were able to collect the minimum number of species occurrence records required for MaxEnt modeling [
39] for the 16 most problematic weeds, and consequently, only these species are used in this study. Species richness was estimated in different regions and across the country. This study will provide fundamental information about current and future potential habitats, which could be useful for developing control and management strategies for invasive weeds.
3. Discussion
Our study investigated the potential habitats of 16 problematic invasive weeds that are currently found in the southern region of South Korea using the MaxEnt modeling approach. The current suitable habitats for the studied invasive weeds that were predicted by the model were highly matched with the current existing records. The average AUC (0.982 ± 0.004), TSS (0.881 ± 0.881), and Kappa (0.64 ± 0.065) values indicated that the model performance was excellent, and that perfect agreement existed between the observations and predictions [
41,
42]. In this study, temperature seasonality, annual mean temperature, and annual precipitation were the dominant driving factors for the determination of the habitats of the studied invasive weeds. Temperature seasonality balances photosynthesis and regulates the growth, reproduction, and other physiological functions of plants [
43].
Similarly, increasing the annual mean temperature and annual precipitation may create suitable habitats for invasive weeds while altering the distribution and abundance of existing native species, which reduces competition with local species [
44]. Therefore, these variables could be critical in determining the spread of invasive weeds in the future. Similar cases were reported by Adhikari et al. [
24] and Wang et al. [
45] for predicting the habitat expansion of invasive and alien plants. However, the other variables examined in this study only had minor contributions to the model.
Climate change may directly or indirectly influence the introduction, dispersion, and establishment of invasive and alien species, and may decrease the resilience of natural ecosystems to invasive species [
6]. Climate change may also modify the geographical ranges and environmental impacts of invasive species, as well as the economic costs necessary for their management [
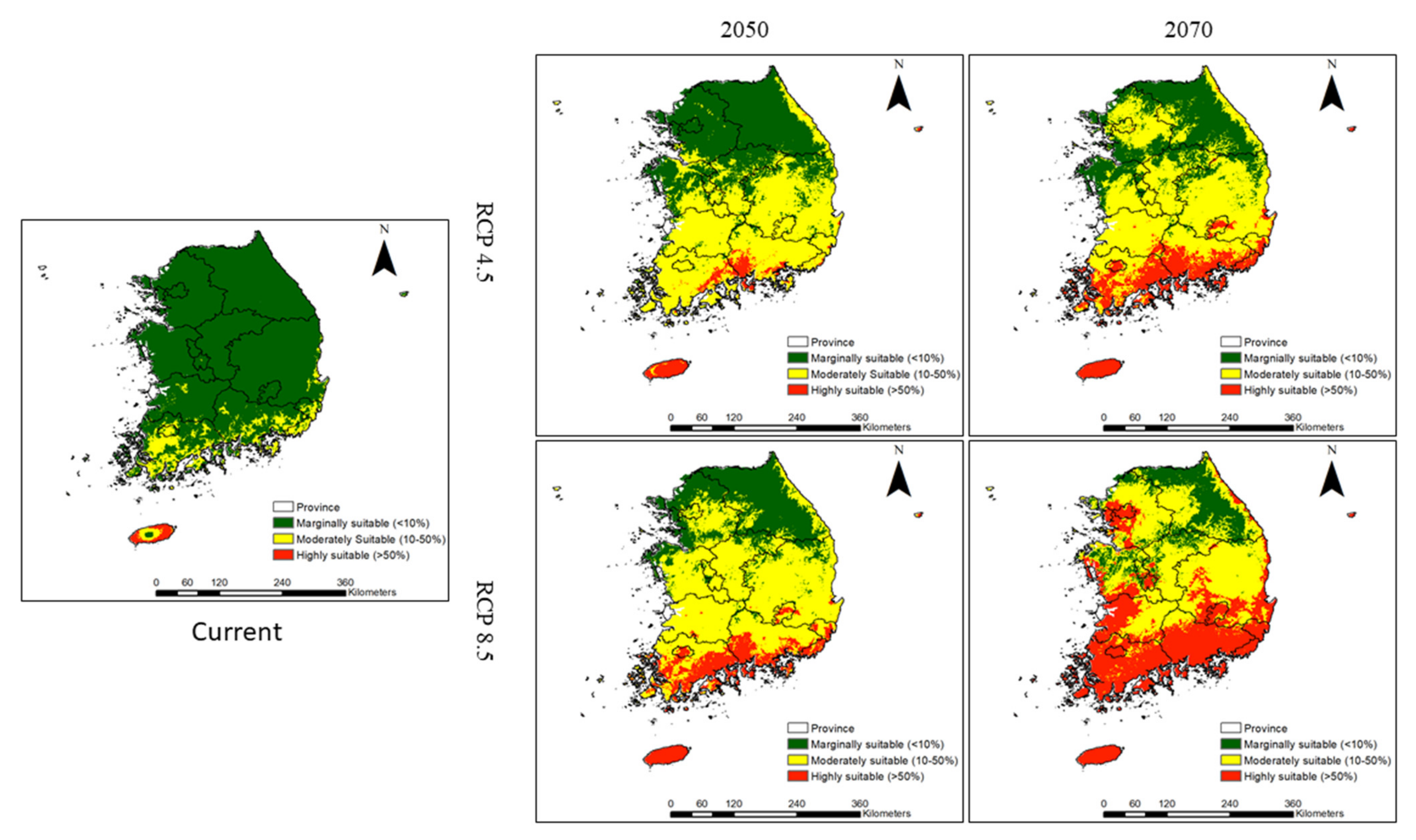
46]. Our model shows that climate change is likely to substantially increase the habitat suitability of invasive weeds in the southern region of South Korea. These results reinforce the spatially explicit evidence that supports the earlier hypothesis that warming temperatures will expand the suitable habitats of invasive plants northwards [
47]. This is also evidenced by previous observations and projections on the impacts of warmer climatic conditions [
5,
24,
48,
49]. For most of the studied invasive weeds, habitat suitability will expand toward the central and northern regions of South Korea under climate change, and this impact will be particularly visible in Chungcheongnam Province, Chungcheongbuk Province, Gyeongsangbuk Province, Gyeonggi Province, and Seoul City, which are up to 454 km from the southern region (for example, Jeju Province and Jeollanam Province), by 2070. Consistent with our findings, several studies have attempted to model the habitat suitability of non-native and invasive species in South Korea and across the globe and have projected the expansion of their ranges northwards in response to climate change [
5,
9,
23,
24,
50]. Some studies on invasive plants showed that the majority of range shifts that were expected to occur by 2070 will occur as early as 2050 in Europe [
51]. To our knowledge, this is the first study to describe the potential impacts of climate change and land cover change on these invasive weeds in South Korea. This study forms part of our ongoing project. We will compare the results obtained in this study with those obtained from other ecological niche models.
In this study, we predicted the habitat suitability of 16 problematic invasive weed species. The rate and extent of habitat suitability were not projected to be consistent among all invasive weed species.
A.
sinicus,
B.
unioloides,
C.
ambrosioides and
S.
officinale var.
leiocarpum are estimated to have relatively high habitat suitability areas, covering 78.44% and 86.91% of the land area of the country by 2050, and 2070 (RCP 4.5), respectively; these values are comparable to those reported in previous studies performed in South Korea [
24,
33,
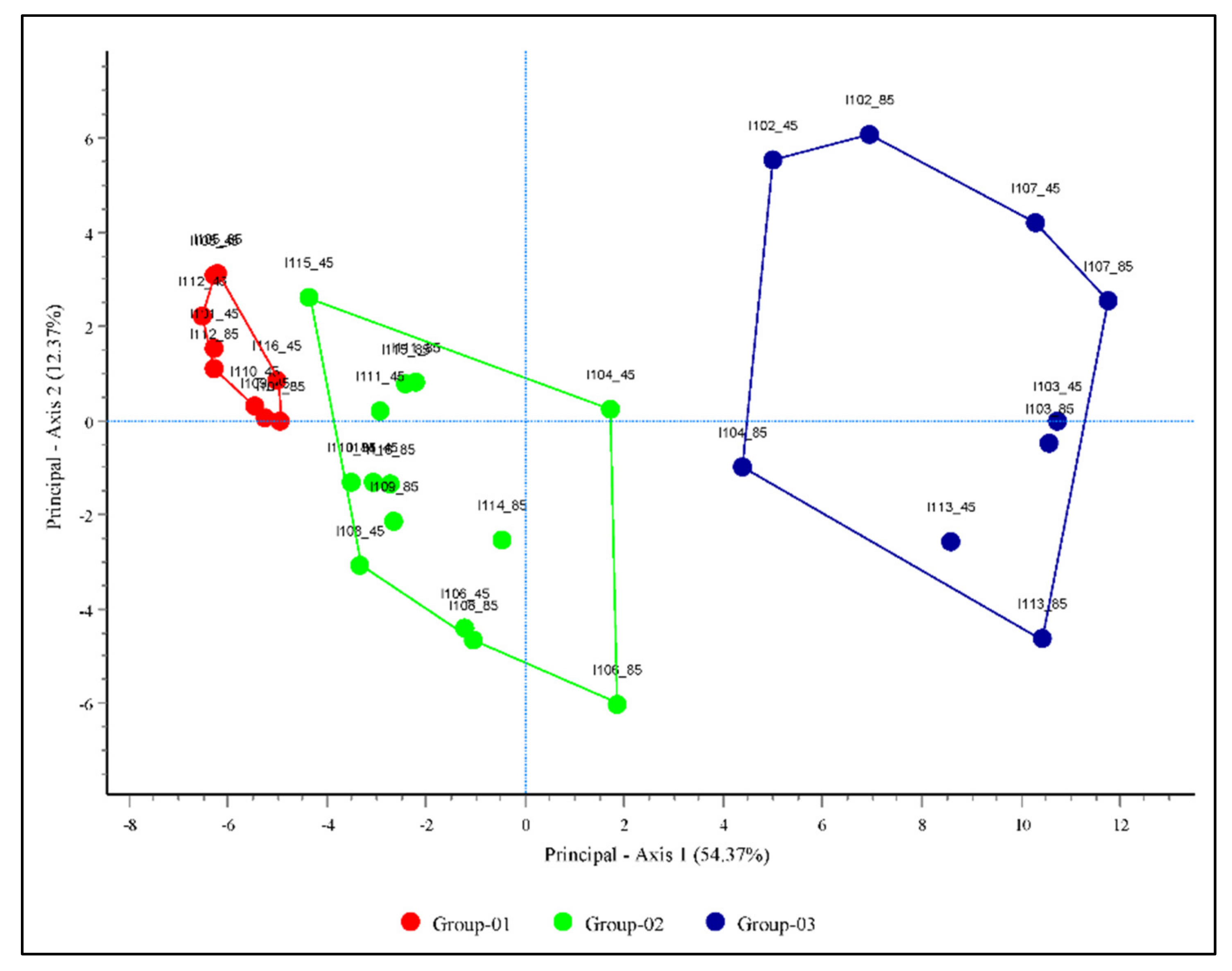
37]. The PCA revealed that the studied invasive weeds could be divided into three groups based on similarities in their invasion potentials and distribution patterns. The invasive weeds included in groups 01, 02, and 03 had low, intermediate, and high invasion potentials and distribution patterns, respectively. Group 01 is currently found in Jeju Province (Jeju Island) and is projected to have very limited habitat expansion along the coastal side of the southern region, for example in Gyeongsang Province, indicating that continental climatic features such as large diurnal and seasonal temperature ranges, low annual precipitation, and low relative humidity may not favor the expansion of these species [
52]. However, the invasive weeds present in groups 02 and 03 are projected to expand continuously toward the central and northern regions with various rates of invasion, indicating that these species could have greater tolerance to continental climates.
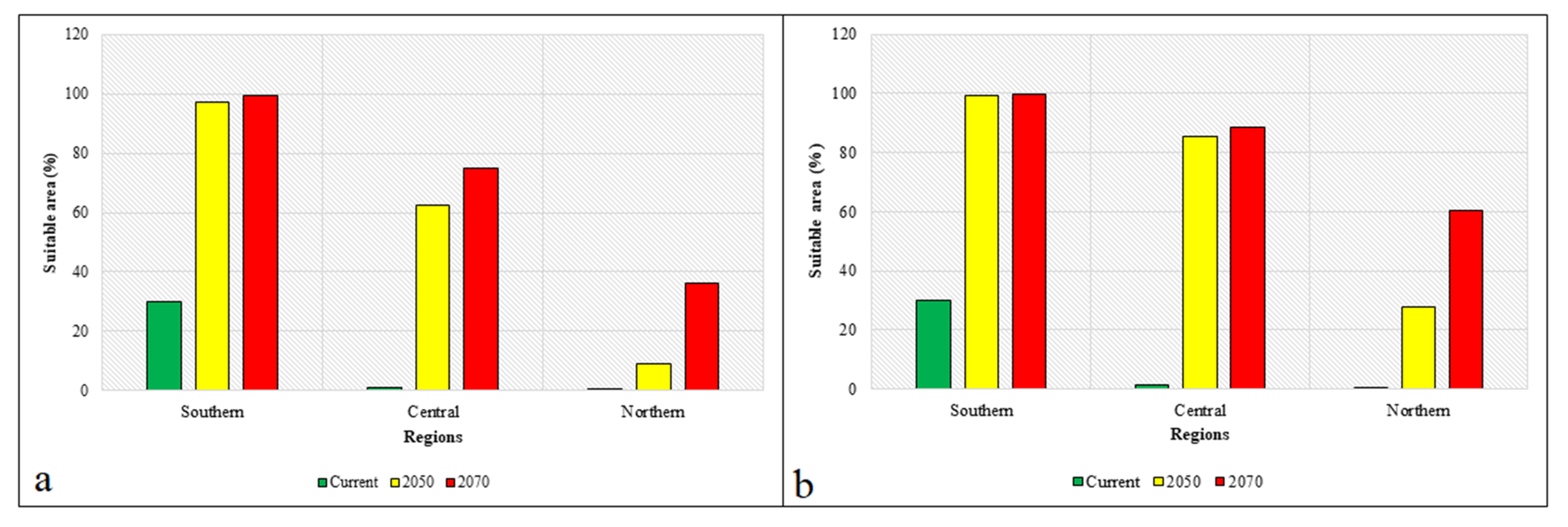
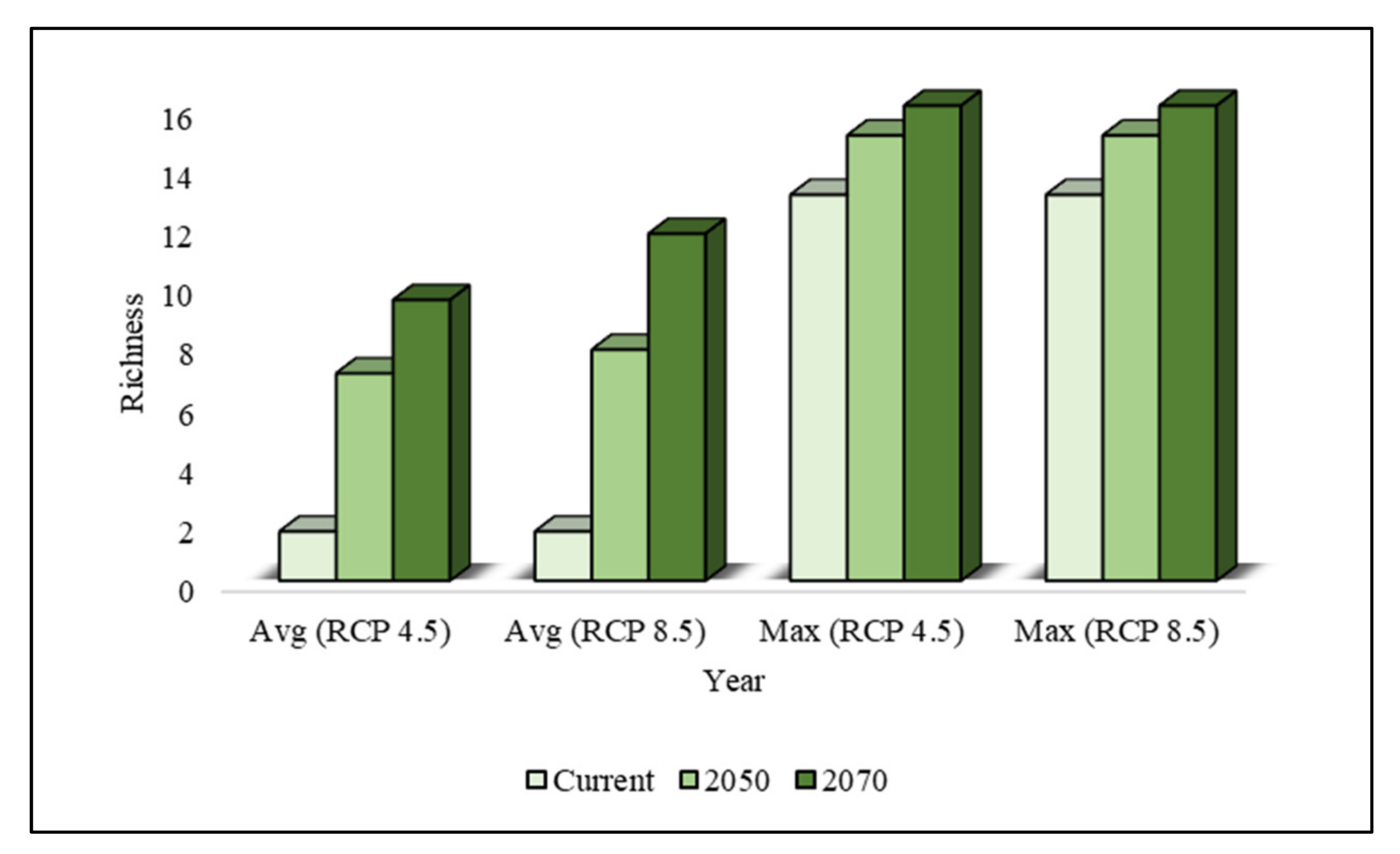
In comparison to other regions, the average and maximum species richness values calculated in the southern region, especially in Jeju Province, were estimated to be the highest under current climatic conditions. Currently, the average species richness in the southern region is estimated to be 11.30, and this region has been invaded by a maximum of 15 invasive weeds, including
Sida rhombifolia,
S.
gallica and
Spergularia rubra; however, the northern region, for example, Gangwan Province, has estimated average and maximum richness values of 0.014 and 4, respectively. These results showed that the introduction, establishment, and dispersion of the studied invasive weeds occur more readily in the southern region. The climatic conditions of this region are characterized by a warm-temperate and humid climate, which favors invasive weeds originating from the tropical and subtropical climates of South America, southern Europe, China, and South Asia [
31]. Usually, invasive weeds that are indigenous to tropical and subtropical countries have much higher critical thermal maxima than native species, suggesting that these invasive species can thrive at higher temperatures and may successfully outcompete native species under climate change [
6]. Under current climatic conditions, the average winter temperature in the southern region is approximately 3 °C, which could favor the survival of warm-adapted invasive weeds in the winter season. However, in the central and northern regions, the average winter temperatures reach approximately −10 °C, which could limit the distribution of such invasive weeds, as described by Hou et al. [
53] and Petitpierre et al. [
54]. With increasing temperatures under climate change, suitable habitats for invasive weeds will expand toward the central and northern regions of South Korea due to the removal of current climatic barriers and the shifting of plant hardiness zones northward [
6,
55], and consequently, species richness is predicted to increase in the future. In addition, climate change will negatively affect native species and ecosystems by changing their phenology, composition, distribution, and adaptability through changes in environmental conditions and by creating difficulties that prevent native species from surviving and competing with invaders, which could be conducive to invasive species taking over newly empty niches [
6,
56].
These future changes in habitat suitability depend not only on the climatic temperature and precipitation variables used in the model, but also on many non-climatic factors, including land topography, soil and habitat characteristics, and on the morphological and physiological advantages of individual plant species, such as short life cycles, high fecundity, strong dispersal abilities, and phenotypic plasticity, which allow them to survive adverse climate conditions [
5,
23,
57,
58]. Thus, under the same climatic conditions in the future, the climatically suitable habitat for each invasive weed species could be different. Many invasive weeds are characteristically recognized as broadly ecologically and environmentally tolerant, such as
S.
gallica and
S.
officinale are salt-resistant and can grow in coastal areas and in very disturbed areas along roadsides [
59,
60]. Similarly, some invasive weeds, such as
Apium leptophyllum,
M. caroliniana,
C.
ambrosioides and
Sida rhombifolia, grow in diverse habitats, including croplands, farmlands, riversides, and dry and drained lands [
27,
28,
31,
32]. Therefore, the suitable habitats for all invasive weeds will expand in the future.
The studied invasive weed species were all introduced to South Korea either intentionally or unintentionally (
Table 8).
A.
sinicus is native to China and was intentionally introduced to South Korea before 1937 to improve pastures; the species subsequently invaded grasslands and interior forests [
31]. Similarly,
B.
unioloides and
Lolium multiflorum were introduced from South America and southern Europe for use as cattle-feed, and have since become invaders of grasslands, forests, and crop lands [
28,
31]. Other invasive weeds were unintentionally introduced in South Korea from different countries in America, Europe, and Asia, probably via foreign trade, tourism, and tidal activity in the seas [
24,
28,
30,
31], and many anthropogenic activities, such as road construction, land cover changes, and the importation of agricultural seeds from foreign countries have accelerated their invasion rates. Invasive weeds have adverse impacts on agricultural and wild ecosystems through increased labor input for weeding, reduced crop production, the replacement of the native forage of cattle and wild herbivores such as roe deer (
Capreolus pygargus) [
61], and their negative effects on forest ecosystems [
62]. Therefore, the economic losses and negative impacts of invasive weeds on food security, biodiversity, and ecosystem services in the near future could increase considerably if control and preventive measures are not adopted in time.
Although this study provided valuable information about the potential habitat suitability of invasive weeds in different provinces of South Korea under current and future climatic conditions, our models were dependent on bioclimatic and some environmental variables, such as land cover change, distance from the road, and distance from water and is disregarding, other important predictors such as land topography, soil characteristics, dispersal capacities, biotic interaction (e.g., facilitation and competition), and vectors driving species invasions, as described by Pysek and Richardson [
63] and Buri et al. [
64]. This study is a part of ongoing research; we would consider using other variables, including topographic and soil characteristics, dispersion capacities, and biotic interactions, to obtain more accurate predictions in the near future.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}