
Context-Dependent Effects of Trichoderma Seed Inoculation on Anthracnose Disease and Seed Yield of Bean (Phaseolus vulgaris): Ambient Conditions Override Cultivar-Specific Differences

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
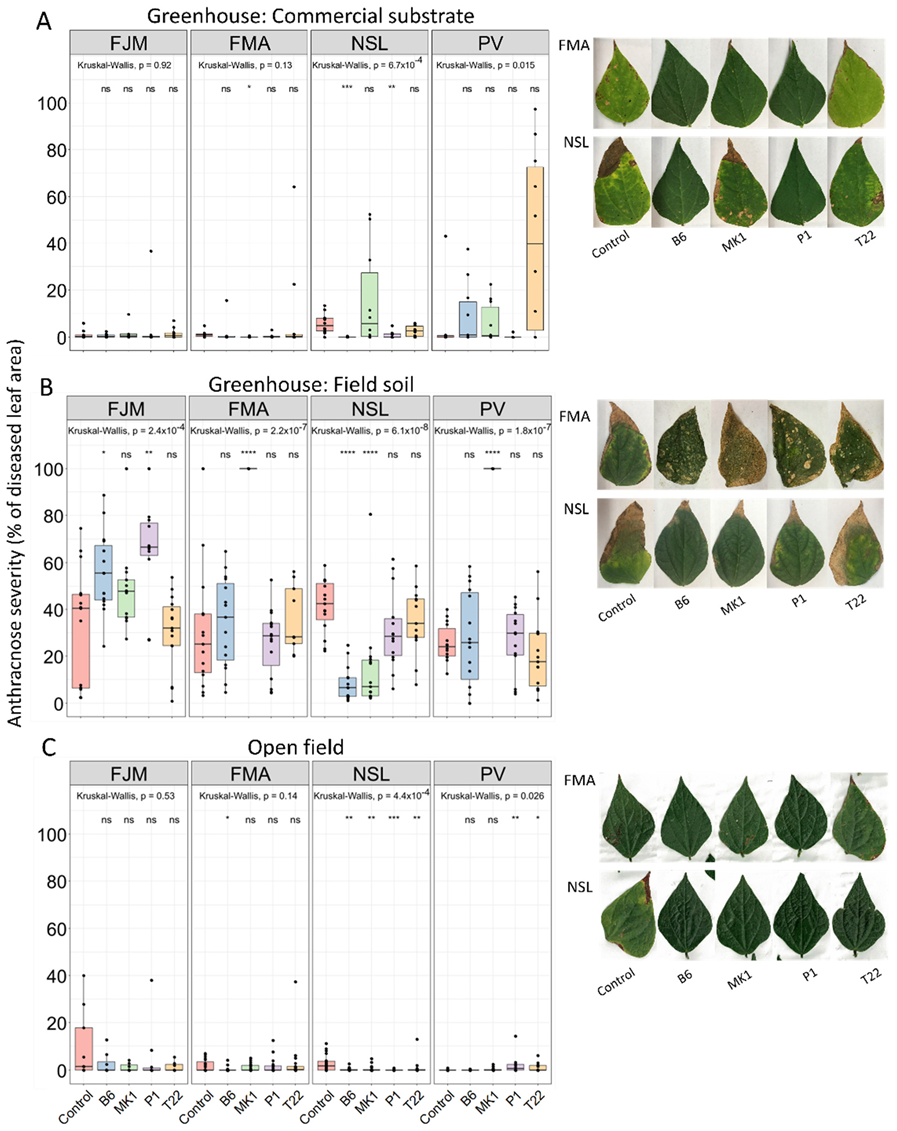
2.1. Ambient Conditions Override Cultivar-Specific Effects of Trichoderma-Inoculation on Anthracnose Severity of Bean Plants
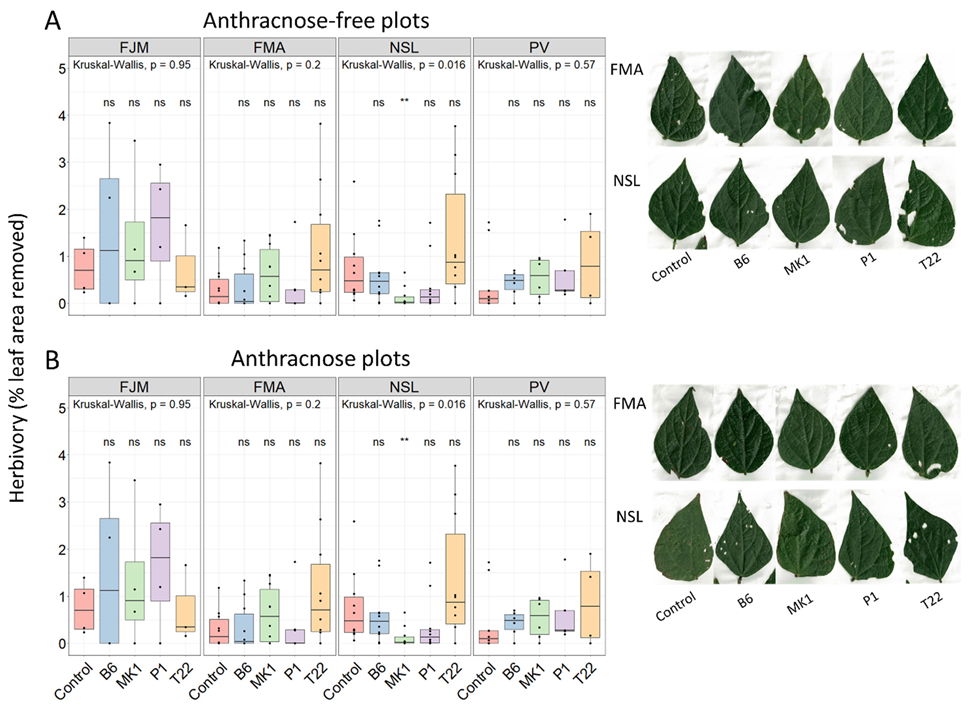
2.2. Mortality and Damage by Non-Controlled Environmental Factors in the Field
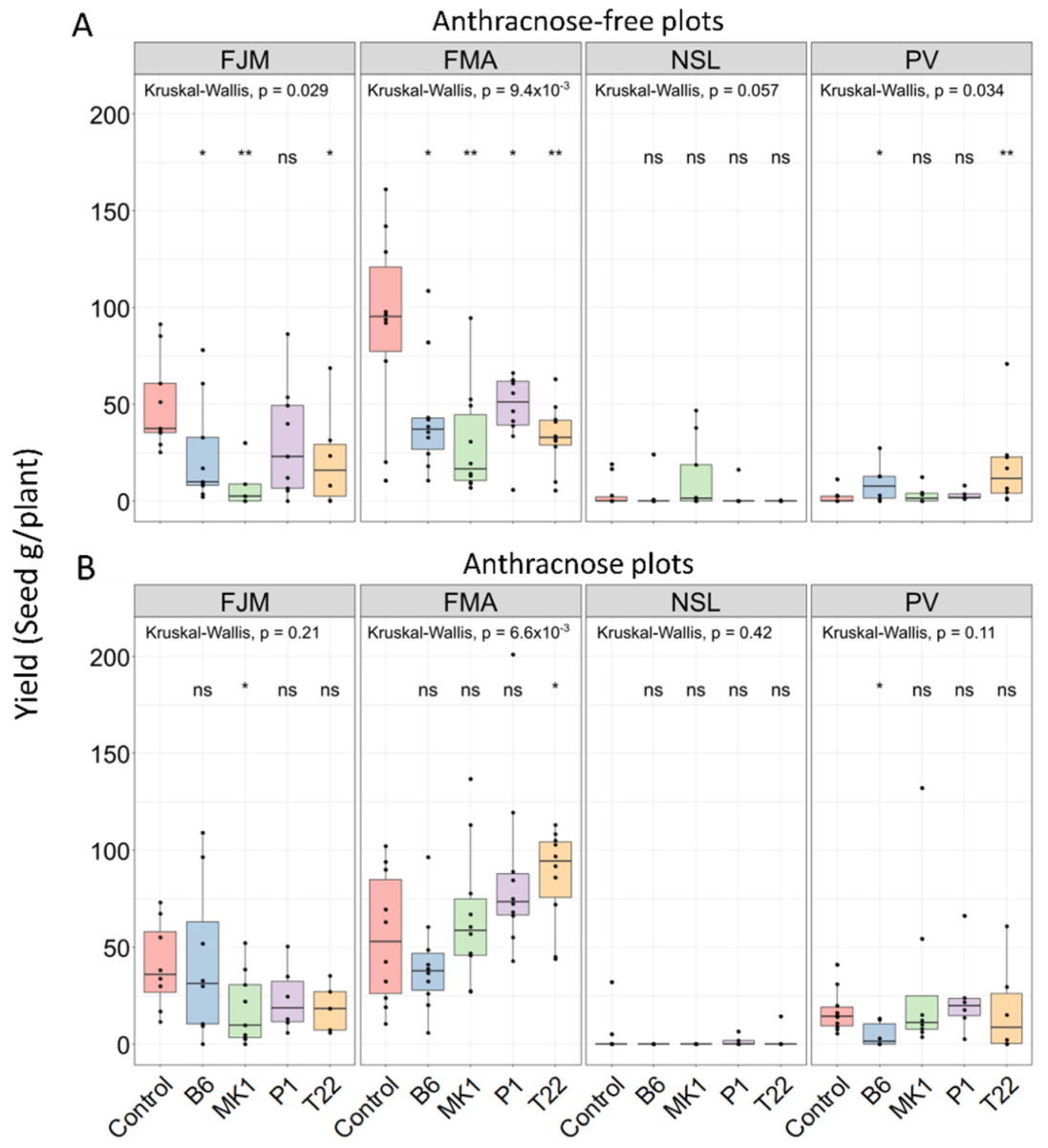
2.3. Seed Yield
3. Conclusions and Outlook
4. Materials and Methods
4.1. Biological Material
4.2. Culturing Conditions and Inoculation
4.3. Quantification of Disease Severity
4.4. Field Experiment: Damage by Non-Target Enemies and Yield
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Partida-Martinez, L.P.P.; Heil, M. The microbe-free plant: Fact or artefact? Front. Plant Sci. 2011, 2, 100. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E. Trichoderma—Not just for biocontrol anymore. Phytoparasitica 2011, 39, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Adetunji, C.O.; Anani, O.A. Bio-fertilizer from Trichoderma: Boom for agriculture production and management of soil- and root-borne plant pathogens. In Innovations in Food Technology: Current Perspectives and Future Goals; Mishra, P., Mishra, R.R., Adetunji, C.O., Eds.; Springer Nature: Singapore, 2020; pp. 245–256. [Google Scholar]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Gomes, E.N.; Elsherbiny, E.A.; Aleem, B.; Bennett, J.W. Beyond classical biocontrol: New perspectives on Trichoderma. In Fungal Biotechnology and Bioengineering. Fungal Biology; Hesham, A.L., Upadhyay, R., Sharma, G., Manoharachary, C., Gupta, V.K., Eds.; Springer Nature Switzerland AG 2020: Cham, Switzerland, 2020; pp. 437–455. [Google Scholar]
- Martinez-Medina, A.; Fernandez, I.; Sánchez-Guzmán, M.; Jung, S.; Pascual, J.; Pozo, M. Deciphering the hormonal signalling network behind the systemic resistance induced by Trichoderma harzianum in tomato. Front. Plant Sci. 2013, 4, 206. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Hermosa, R.; Rubio, M.B.; Cardoza, R.E.; Nicolás, C.; Monte, E.; Gutiérrez, S. The contribution of Trichoderma to balancing the costs of plant growth and defense. Int. Microbiol. 2013, 16, 69–80. [Google Scholar] [PubMed]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esparza-Reynoso, S.; Pelagio-Flores, R.; López-Bucio, J. Chapter 3—Mechanism of plant immunity triggered by Trichoderma. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Zeilinger, S., Singh, H.B., Druzhinina, I., Eds.; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, MA, USA, 2020; pp. 57–73. [Google Scholar]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agricult. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Lopez-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Del-Val, E.; Larsen, J. The root endophytic fungus Trichoderma atroviride induces foliar herbivory resistance in maize plants. Appl. Soil Ecol. 2018, 124, 45–53. [Google Scholar] [CrossRef]
- Bazghaleh, N.; Prashar, P.; Woo, S.; Vandenberg, A. Effects of lentil genotype on the colonization of beneficial Trichoderma species and biocontrol of Aphanomyces root rot. Microorganisms 2020, 8, 1290. [Google Scholar] [CrossRef]
- Galletti, S.; Paris, R.; Cianchetta, S. Selected isolates of Trichoderma gamsii induce different pathways of systemic resistance in maize upon Fusarium verticillioides challenge. Microbiol. Res. 2020, 233, 126406. [Google Scholar] [CrossRef] [PubMed]
- Ousley, M.A.; Lynch, J.M.; Whipps, J.M. Effect of Trichoderma on plant growth: A balance between inhibition and growth promotion. Microb. Ecol. 1993, 26, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Mendez, W. Trichoderma interactions in vegetable rhizosphere under tropical weather conditions. In Trichoderma, Rhizosphere Biology; Sharma, A.K., Sharma, P., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2020; pp. 293–314. [Google Scholar]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–plant–pathogen interactions for better development of biocontrol applications. J. Fungi 2021, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Salzano, M.; Villano, C.; Aversano, R.; Lorito, M.; Ruocco, M.; Docimo, T.; Piccinelli, A.L.; D’Agostino, N.; Tucci, M. Transcriptome reprogramming, epigenetic modifications and alternative splicing orchestrate the tomato root response to the beneficial fungus Trichoderma harzianum. Horticult. Res. 2019, 6, 5. [Google Scholar] [CrossRef]
- Malinich, E.A.; Wang, K.; Mukherjee, P.K.; Kolomiets, M.; Kenerley, C.M. Differential expression analysis of Trichoderma virens RNA reveals a dynamic transcriptome during colonization of Zea mays roots. BMC Genom. 2019, 20, 280. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Van Wees, S.C.M.; Pieterse, C.M.J. Airborne signals from Trichoderma fungi stimulate iron uptake responses in roots resulting in priming of jasmonic acid-dependent defences in shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ. 2017, 40, 2691–2705. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, D.; Bossi, S.; Cascone, P.; Digilio, M.C.; Prieto, J.D.; Fanti, P.; Guerrieri, E.; Iodice, L.; Lingua, G.; Lorito, M.; et al. Tomato below ground-above ground interactions: Trichoderma longibrachiatum affects the performance of Macrosiphum euphorbiae and its natural antagonists. Mol. Plant Microbe Interact. 2013, 26, 1249–1256. [Google Scholar] [CrossRef] [Green Version]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 2011, 12, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Guzman, P.; Porras-Troncoso, M.D.; Olmedo-Monfil, V.; Herrera-Estrella, A. Trichoderma species: Versatile plant symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Parwi; Isnatin, U.; Hamawi, M.; Etica, U. Growth and yield of shallot (Allium cepa L.) in response of organic fertilizers and Trichoderma asperellum. J. Phys. Conf. Ser. 2019, 1381, 012004. [Google Scholar] [CrossRef]
- Naznin, A.; Hossain, M.; Ara, K.A.-M.; Hoque, A.; Islam, M.; Hasan, T. Influence of organic amendments and bio-control agent on yield and quality of Tuberose. J. Horticult. 2015, 2, 4. [Google Scholar]
- Caporale, A.G.; Vitaglione, P.; Troise, A.D.; Pigna, M.; Ruocco, M. Influence of three different soil types on the interaction of two strains of Trichoderma harzianum with Brassica rapa subsp. sylvestris cv. esculenta, under soil mineral fertilization. Geoderma 2019, 350, 11–18. [Google Scholar] [CrossRef]
- Nepali, B.; Subedi, S.; Bhattarai, S.; Marahatta, S.; Bhandari, D.; Shrestha, J. Bio-fertilizer activity of Trichoderma viride and Pseudomonas fluorescens as growth and yield promoter for maize. J. Agric. Sci. 2020, 2, 191–195. [Google Scholar]
- Mahato, S.; Bhuju, S.; Shrestha, J. Effect of Trichoderma viride as biofertilizer on growth and yield of wheat. Malays. J. Sustain. Agric. 2018, 2, 1–5. [Google Scholar] [CrossRef]
- Haque, M.; Ilias, G.N.M.; Molla, A.H. Impact of Trichoderma-enriched biofertilizer on the growth and yield of mustard (Brassica rapa L.) and tomato (Solanum lycopersicon Mill.). Agric. J. 2012, 10, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart, A.; et al. Environmental growth conditions of Trichoderma spp. affects indole acetic acid derivatives, volatile organic compounds, and plant growth promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzies, J.G. A strain of Trichoderma viride pathogenic to germinating seedlings of cucumber, pepper and tomato. Plant Pathol. 1993, 42, 784–791. [Google Scholar] [CrossRef]
- Moni Ram, R.; Singh, H.B. Trichoderma spp.: An opportunistic pathogen. Biotech Today 2018, 8, 16–24. [Google Scholar]
- McFadden, A.G.; Sutton, J.C. Relationship of populations of Trichoderma spp. in soil to disease in maize. Can. J. Plant Sci. 1975, 55, 579–586. [Google Scholar] [CrossRef]
- Baker, R.; Elad, Y.; Chet, I. The controlled experiment in the scientific method with special emphasis on biological control. Phytopathology 1984, 74, 1019–1021. [Google Scholar] [CrossRef] [Green Version]
- Sunpapao, A.; Chairin, T.; Ito, S. The biocontrol by Streptomyces and Trichoderma of leaf spot disease caused by Curvularia oryzae in oil palm seedlings. Biol. Control 2018, 123, 36–42. [Google Scholar] [CrossRef]
- Yobo, K.S.; Laing, M.D.; Hunter, C.H. Effects of single and dual applications of selected Trichoderma and Bacillus isolates on performance of dry bean seedlings grown in composted pine bark growth medium under shadehouse conditions. J. Plant Nutr. 2009, 32, 1271–1289. [Google Scholar] [CrossRef]
- Marín-Guirao, J.I.; Rodríguez-Romera, P.; Lupión-Rodríguez, B.; Camacho-Ferre, F.; Tello-Marquina, J.C. Effect of Trichoderma on horticultural seedlings’ growth promotion depending on inoculum and substrate type. J. Appl. Microbiol. 2016, 121, 1095–1102. [Google Scholar] [CrossRef]
- Illescas, M.; Rubio, M.B.; Hernández-Ruiz, V.; Morán-Diez, M.E.; Martínez de Alba, A.E.; Nicolás, C.; Monte, E.; Hermosa, R. Effect of inorganic N top dressing and Trichoderma harzianum seed-inoculation on crop yield and the shaping of root microbial communities of wheat plants cultivated under high basal N fertilization. Front. Plant Sci. 2020, 11, 575861. [Google Scholar] [CrossRef]
- Crop Science Dept, Oregon State University Extended Campus (O.E.C.) Classification of Crops and Their Role in Human Nutrition. Available online: https://oregonstate.edu/instruct/css/330/two/index2.htm (accessed on 6 February 2021).
- Castro-Guerrero, N.A.; Isidra-Arellano, M.C.; Mendoza-Cozatl, D.G.; Valdés-López, O. Common bean: A legume model on the rise for unraveling responses and adaptations to iron, zinc, and phosphate deficiencies. Front. Plant Sci. 2016, 7, 600. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.R.; Kmiecik, K. Common bean: Economic importance and relevance to biological science research. In The Common Bean Genome; Pérez de la Vega, M., Santalla, M., Marsolais, F., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–20. [Google Scholar]
- Rendón-Anaya, M.; Montero-Vargas, J.M.; Saburido-Álvarez, S.; Vlasova, A.; Capella-Gutierrez, S.; Ordaz-Ortiz, J.J.; Aguilar, O.M.; Vianello-Brondani, R.P.; Santalla, M.; Delaye, L.; et al. Genomic history of the origin and domestication of common bean unveils its closest sister species. Genome Biol. 2017, 18, 60. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, S.; Singh Rathore, U.; Kumar, K. Multi-trait Trichoderma for improving plant health of grain legumes. In Microbial Mitigation of Stress Response of Food Legumes, 1st ed.; Amaresan, N., Murugesan, S., Kumar, K., Sankaranarayanan, A., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 85–102. [Google Scholar]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Márquez-Benavidez, L.; Rizo-León, M.Á.; Montaño-Arias, N.M.; Ruiz-Nájera, R.; Sánchez-Yáñez, J.M. Respuesta de Phaseolus vulgaris a la inoculación de diferentes dosis de Trichoderma harzianum con el fertilizante nitrogenado reducido al 50%. J. Selva Andin. Res. Soc. 2017, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Mayo-Prieto, S.; Marra, R.; Vinale, F.; Rodríguez-González, Á.; Woo, S.L.; Lorito, M.; Gutiérrez, S.; Casquero, P.A. Effect of Trichoderma velutinum and Rhizoctonia solani on the metabolome of bean plants (Phaseolus vulgaris L.). Internat. J. Mol. Sci. 2019, 20, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo, S.; Gutierrez, S.; Malmierca, M.; Lorenzana, A.; Campelo, M.; Hermosa, R.; Casquero, P. Influence of Rhizoctonia solani and Trichoderma spp. in growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Front. Plant Sci. 2015, 6, 00685. [Google Scholar] [CrossRef] [Green Version]
- González-Marquetti, I.; Infante-Martínez, D.; Arias-Vargas, Y.; Gorrita-Ramírez, S.; Hernández-García, T.; De la Noval-Pons, B.M.; Martínez-Coca, B.; Peteira, B. Efecto de Trichoderma asperellum Samuels, Lieckfeldt & Nirenberg sobre indicadores de crecimiento y desarrollo de Phaseolus vulgaris L. cultivar BAT-304. Rev. Prot. Veget. 2019, 34, e05. [Google Scholar]
- Pereira, J.L.; Queiroz, R.M.L.; Charneau, S.O.; Felix, C.R.; Ricart, C.A.O.; Da Silva, F.L.; Steindorff, A.S.; Ulhoa, C.J.; Noronha, E.F. Analysis of Phaseolus vulgaris response to its association with Trichoderma harzianum (ALL-42) in the presence or absence of the phytopathogenic fungi Rhizoctonia solani and Fusarium solani. PLoS ONE 2014, 9, e98234. [Google Scholar] [CrossRef]
- Ghoniem, A.A.; Abd El-Hai, K.M.; El-khateeb, A.Y.; Eldadamony, N.M.; Mahmoud, S.F.; Elsayed, A. Enhancing the potentiality of Trichoderma harzianum against Pythium pathogen of beans using chamomile (Matricaria chamomilla, L.) flower extract. Molecules 2021, 26, 1178. [Google Scholar] [CrossRef]
- Bedine Boat, M.A.; Sameza, M.L.; Iacomi, B.; Tchameni, S.N.; Boyom, F.F. Screening, identification and evaluation of Trichoderma spp. for biocontrol potential of common bean damping-off pathogens. Biocontrol Sci. Technol. 2019, 30, 228–242. [Google Scholar] [CrossRef]
- Carvalho, D.D.C.; Lobo Junior, M.; Martins, I.; Inglis, P.W.; Mello, S.C.M. Biological control of Fusarium oxysporum f. sp. phaseoli by Trichoderma harzianum and its use for common bean seed treatment. Trop. Plant Pathol. 2014, 39, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Toghueo, R.M.K.; Eke, P.; Zabalgogeazcoa, Í.; De Aldana, B.R.V.; Nana, L.W.; Boyom, F.F. Biocontrol and growth enhancement potential of two endophytic Trichoderma spp. from Terminalia catappa against the causative agent of common bean root rot (Fusarium solani). Biol. Control 2016, 96, 8–20. [Google Scholar] [CrossRef]
- Abeysinghe, S. Systemic resistance induced by Trichoderma harzianum RU01 against Uromyces appendiculatus on Phaseolus vulgaris. J. Natl. Sci. Found. Sri Lanka 2009, 37, 203–207. [Google Scholar] [CrossRef]
- Padder, B.A.; Sharma, P.N.; Awale, H.E.; Kelly, J.D. Colletotrichum lindemuthianum, the causal agent of bean anthracnose. J. Plant Pathol. 2017, 99, 318–330. [Google Scholar]
- Singh, S.P.; Schwartz, H.F. Breeding common bean for resistance to diseases: A review. Crop Sci. 2010, 50, 2199–2223. [Google Scholar] [CrossRef]
- Mendgen, K.; Hahn, M. Plant infection and the establishment of fungal biotrophy. Trends Plant Sci. 2002, 7, 352–356. [Google Scholar] [CrossRef] [Green Version]
- Perfect, S.E.; Hughes, H.B.; O’Connell, R.J.; Green, J.R. Colletotrichum: A model genus for studies on pathology and fungal–plant interactions. Fungal Genet. Biol. 1999, 27, 186–198. [Google Scholar] [CrossRef]
- Barros, S.T.; Oliveira, N.T.; Bastos, S.T.G. Trichoderma spp. en el control biológico de Colletotricum lindemuthianum (Sacc.etmagn) Scribb, agente de la antracnosis del frijol (Phaseolus vulgaris L.). Bol. Micol. 1995, 10, 5–11. [Google Scholar] [CrossRef]
- Harman, G.E.; Petzoldt, R.; Comis, A.; Chen, J. Interactions between Trichoderma harzianum strain T22 and maize inbred line Mo17 and effects of these interactions on diseases caused by Pythium ultimum and Colletotrichum graminicola. Phytopathology 2004, 94, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Christmann, P.E.T.P.; Dalla Pria, M.; Siqueira Hennipman, H.; Godoy, A.R. In vitro control of Colletotrichum lindemuthianum by Trichoderma spp. and in vivo with alternative products. Int. J. Adv. Eng. Res. Sci. 2019, 6, 74–80. [Google Scholar] [CrossRef]
- Acosta-Gallegos, J.A.; Ochoa-Marquez, R.; Arrieta-Montiel, M.P.; Ibarra-Perez, F.; Pajarito-Ravelero, A.; Sánchez-Valdez, I. Registration of “Pinto Villa” common bean. Crop Sci. 1995, 35, 1211. [Google Scholar] [CrossRef]
- Ibarra-Perez, F.J.; Acosta-Gallegos, J.A.; Cazares-Enriquez, B.; Rosales-Serna, R.; Kelly, J.D. Registration of ‘Flor de Mayo 2000’ common bean. Crop Sci. 2005, 45, 2657–2658. [Google Scholar] [CrossRef]
- Castellanos-Ramos, J.Z.; Guzmán-Maldonado, H.; Kelly, J.D.; Acosta-Gallegos, J.A. Registration of ‘Flor de Junio Marcela’ common bean. Crop Sci. 2003, 43, 1121–1122. [Google Scholar] [CrossRef]
- Quintana-Rodriguez, E.; Morales-Vargas, A.T.; Molina-Torres, J.; Ádame-Alvarez, R.M.; Acosta-Gallegos, J.A.; Heil, M. Plant volatiles cause direct, induced and associational resistance in common bean to the fungal pathogen Colletotrichum lindemuthianum. J. Ecol. 2015, 103, 250–260. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Mengiste, T.D.; Myers, J.R.; Egel, D.S.; Hoagland, L.A. Tomato domestication attenuated responsiveness to a beneficial soil microbe for plant growth promotion and induction of systemic resistance to foliar pathogens. Front. Microbiol. 2020, 11, e604566. [Google Scholar] [CrossRef]
- Córdova-Campos, O.; Adame-Álvarez, R.M.; Acosta-Gallegos, J.; Heil, M. Domestication affected the basal and induced disease resistance in common bean (Phaseolus vulgaris). Eur. J. Plant Pathol. 2012, 134, 367–379. [Google Scholar] [CrossRef]
- Vos, I.A.; Pieterse, C.M.J.; Van Wees, S.C.M. Costs and benefits of hormone-regulated plant defences. Plant Pathol. 2013, 62, 43–55. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.; Kang, J.-H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Macías-Rodríguez, L.; Contreras-Cornejo, H.A.; Adame-Garnica, S.G.; Del-Val, E.; Larsen, J. The interactions of Trichoderma at multiple trophic levels: Inter-kingdom communication. Microbiol. Res. 2020, 240, 126552. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Appels, F.; Van Wees, S. Impact of salicylic acid- and jasmonic acid-regulated defences on root colonization by Trichoderma harzianum T-78. Plant Signal. Behav. 2017, 12, e1345404. [Google Scholar] [CrossRef] [Green Version]
- Morán-Diez, M.E.; Tranque, E.; Bettiol, W.; Monte, E.; Hermosa, R. Differential response of tomato plants to the application of three Trichoderma species when evaluating the control of Pseudomonas syringae populations. Plants 2020, 9, 626. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Viveros-Bremauntz, F.; Del-Val, E.; Macías-Rodríguez, L.; López-Carmona, D.A.; Alarcón, A.; González-Esquivel, C.E.; Larsen, J. Alterations of foliar arthropod communities in a maize agroecosystem induced by the root-associated fungus Trichoderma harzianum. J. Pest Sci. 2021, 94, 363–374. [Google Scholar] [CrossRef]
- Carvalho, D.D.C.; Marques de Mello, S.C.; Lobo Júnior, M.; Alaerson, M.G. Biocontrol of seed pathogens and growth promotion of common bean seedlings by Trichoderma harzianum. Pesqui. Agropecu. Bras. 2011, 46, 822–828. [Google Scholar] [CrossRef] [Green Version]
- Aly, A.; Hussein, E.; El-Awamri, A.; Habeb, M.; Abd-Elsalam, K. Beneficial and deleterious effects of Trichoderma harzianum and T. longibrachiatum on growth of cotton seedlings and their biocontrol capacity against seedling damping-off. Rom. Agric. Res. 2014, 4, 1–9. [Google Scholar]
- Naseby, D.C.; Pascual, J.A.; Lynch, J.M. Effect of biocontrol strains of Trichoderma on plant growth, Pythium ultimum populations, soil microbial communities and soil enzyme activities. J. Appl. Microbiol. 2000, 88, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Ban, G.; Akanda, S.; Maino, M. The effect of Trichoderma on the growth and development of tomato and bean under greenhouse and field conditions. Ann. Trop. Res. 2018, 40, 35–45. [Google Scholar] [CrossRef]
- Twizeyimana, M.; Ojiambo, P.S.; Ikotun, T.; Paul, C.; Hartman, G.L.; Bandyopadhyay, R. Comparison of field, greenhouse, and detached-leaf evaluations of soybean germplasm for resistance to Phakopsora pachyrhizi. Plant Dis. 2007, 91, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.; Heil, M. Costs and trade-offs associated with induced resistance. Physiol. Mol. Plant Pathol. 2007, 71, 3–17. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 July 2021).
- RStudio Team. RStudio: Integrated Development for R 2020; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 1 July 2021).
- Wickman, H. ggPlots2 Elegant Graphics for Data Analysis 2016; Springer-International Publishing: Cham, Switzerland, 2016; p. 260. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2′ Based Publication Ready Plots. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 1 July 2021).




{kind=link}
{kind=link}
{kind=link}
| Source | df | Sum of Squares | Mean Square | S | p |
|---|---|---|---|---|---|
| Setup | 2 | 599 | 299.56 | 3.7590 | 0.0238868 |
| Cultivar | 3 | 376 | 125.33 | 1.5727 | 0.1949059 |
| Trichoderma | 4 | 1582 | 395.51 | 4.9631 | 0.0006132 |
| Setup:Cultivar | 6 | 5340 | 889.93 | 11.1674 | 8.470 × 10−12 |
| Setup:Trichoderma | 8 | 2430 | 303.76 | 3.8118 | 0.0002237 |
| Cultivar:Trichoderma | 12 | 3447 | 287.21 | 3.6041 | 3.270 × 10−5 |
| Setup:Cultivar:Trichoderma | 24 | 7732 | 322.18 | 4.0428 | 8.799 × 10−10 |
| Residuals | 569 | 45,344 | 49.69 |
| Soilborne Mortality | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Anthracnose-Free | Anthracnose Plot | ||||||||||
| Ctrl. | B6 | MK1 | P1 | T22 | Ctrl. | B6 | MK1 | P1 | T22 | ||
| FJM | 7 | 8 | 7 | 10 | 10 | FJM | 8 | 8 | 7 | 10 | 11 |
| FMA | 4 | 5 | 3 | 6 | 3 | FMA | 6 | 6 | 4 | 4 | 4 |
| NSL | 6 | 5 | 7 | 5 | 8 | NSL | 5 | 10 | 10 | 11 | 10 |
| PV | 6 | 10 | 10 | 11 | 8 | PV | 6 | 10 | 8 | 10 | 10 |
| Mortality before Reproduction | |||||||||||
| Anthracnose-Free | Anthracnose Plot | ||||||||||
| Ctrl. | B6 | MK1 | P1 | T22 | Ctrl. | B6 | MK1 | P1 | T22 | ||
| FJM | 1 | 0 | 4 | 0 | 1 | FJM | 0 | 1 | 1 | 0 | 0 |
| FMA | 0 | 0 | 0 | 0 | 0 | FMA | 0 | 0 | 0 | 0 | 0 |
| NSL | 7 | 9 | 3 | 10 | 6 | NSL | 9 | 6 | 6 | 3 | 5 |
| PV | 6 | 0 | 2 | 0 | 0 | PV | 0 | 3 | 0 | 0 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Moreno, K.; Ruocco, M.; Monti, M.M.; Vega, O.M.d.l.; Heil, M. Context-Dependent Effects of Trichoderma Seed Inoculation on Anthracnose Disease and Seed Yield of Bean (Phaseolus vulgaris): Ambient Conditions Override Cultivar-Specific Differences. Plants 2021, 10, 1739. https://doi.org/10.3390/plants10081739
Gutiérrez-Moreno K, Ruocco M, Monti MM, Vega OMdl, Heil M. Context-Dependent Effects of Trichoderma Seed Inoculation on Anthracnose Disease and Seed Yield of Bean (Phaseolus vulgaris): Ambient Conditions Override Cultivar-Specific Differences. Plants. 2021; 10(8):1739. https://doi.org/10.3390/plants10081739
Chicago/Turabian StyleGutiérrez-Moreno, Karina, Michelina Ruocco, Maurilia Maria Monti, Octavio Martínez de la Vega, and Martin Heil. 2021. "Context-Dependent Effects of Trichoderma Seed Inoculation on Anthracnose Disease and Seed Yield of Bean (Phaseolus vulgaris): Ambient Conditions Override Cultivar-Specific Differences" Plants 10, no. 8: 1739. https://doi.org/10.3390/plants10081739
APA StyleGutiérrez-Moreno, K., Ruocco, M., Monti, M. M., Vega, O. M. d. l., & Heil, M. (2021). Context-Dependent Effects of Trichoderma Seed Inoculation on Anthracnose Disease and Seed Yield of Bean (Phaseolus vulgaris): Ambient Conditions Override Cultivar-Specific Differences. Plants, 10(8), 1739. https://doi.org/10.3390/plants10081739