
Biopriming of Maize Seeds with a Novel Bacterial Strain SH-6 to Enhance Drought Tolerance in South Korea

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Isolation and Characterization
2.2. Orange Media Test and Congo Red Assay
2.3. Polyethylene Glycol Tolerance Test
2.4. Phosphate Solubilizing-Index Assay
2.5. Siderophore Production Assay
2.6. Production of Indole Acetic Acid (IAA)
2.7. Molecular Characterization
2.8. Oxidative Stress Media Test
2.9. Quantification of Abscisic Acid (ABA) and Sugar Content in Bacterial Isolate SH-6
2.10. Biopriming Maize Seeds
= G4/10 × 100, G7/10 × 100,
2.11. Early Seedling Metrics
3. Statistical Analysis
4. Results
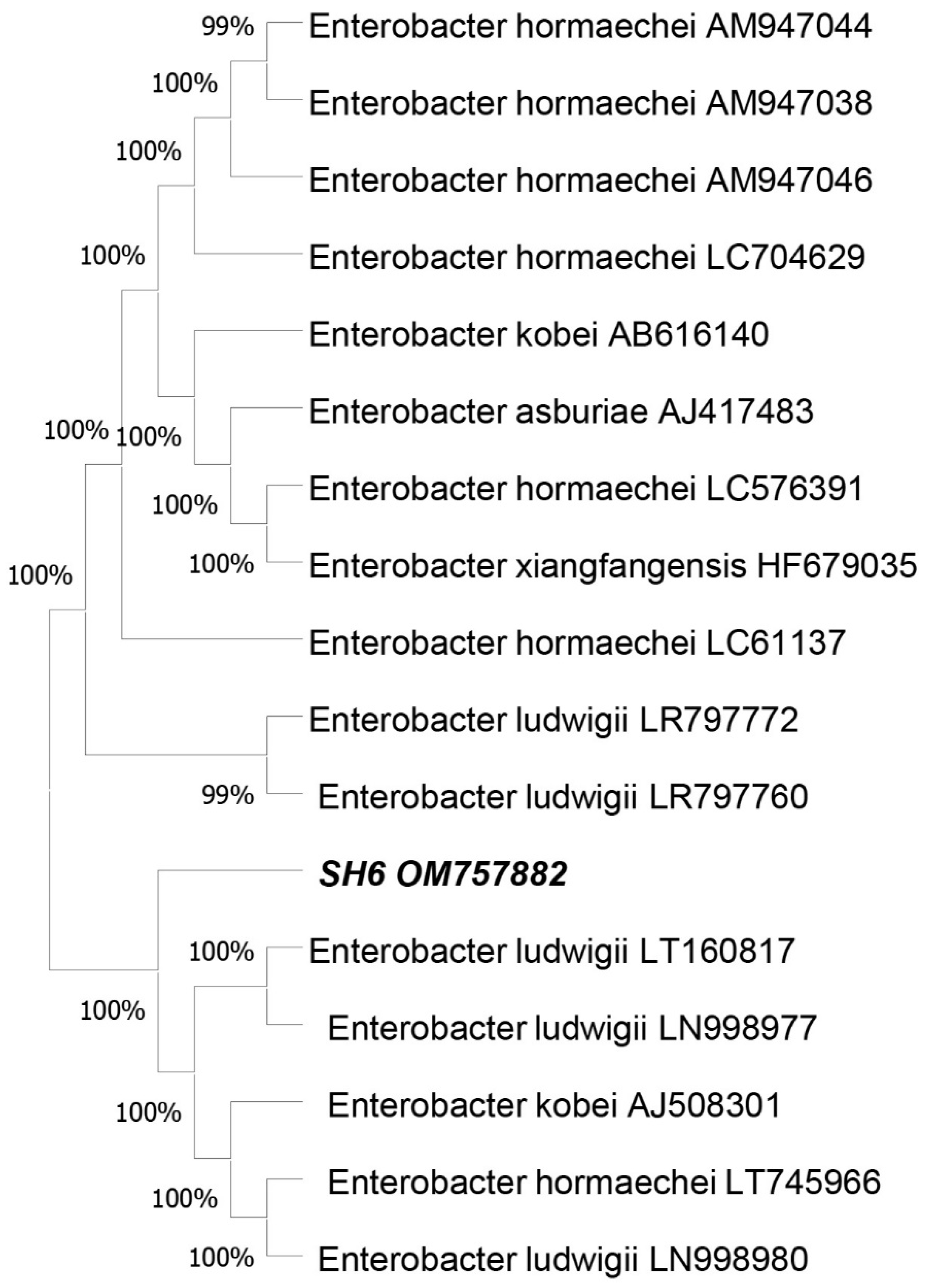
4.1. Molecular Characterization Assay
4.2. Analysis of Siderophore, Indole Acetic Acid, and Exopolysacchrides
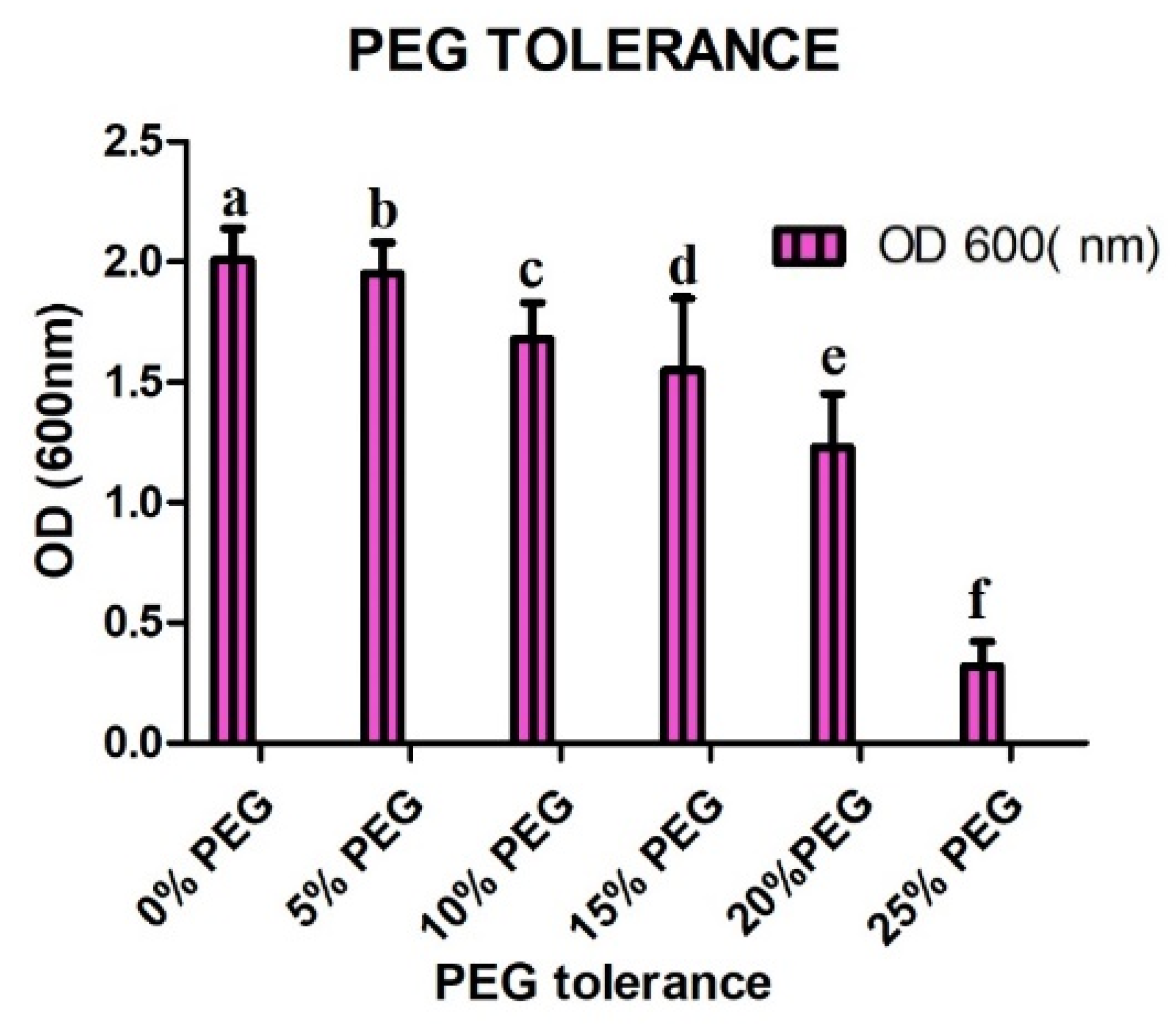
4.3. Drought Tolerance Assay Results
4.4. Sucrose and Abscisic Acid Analysis Results
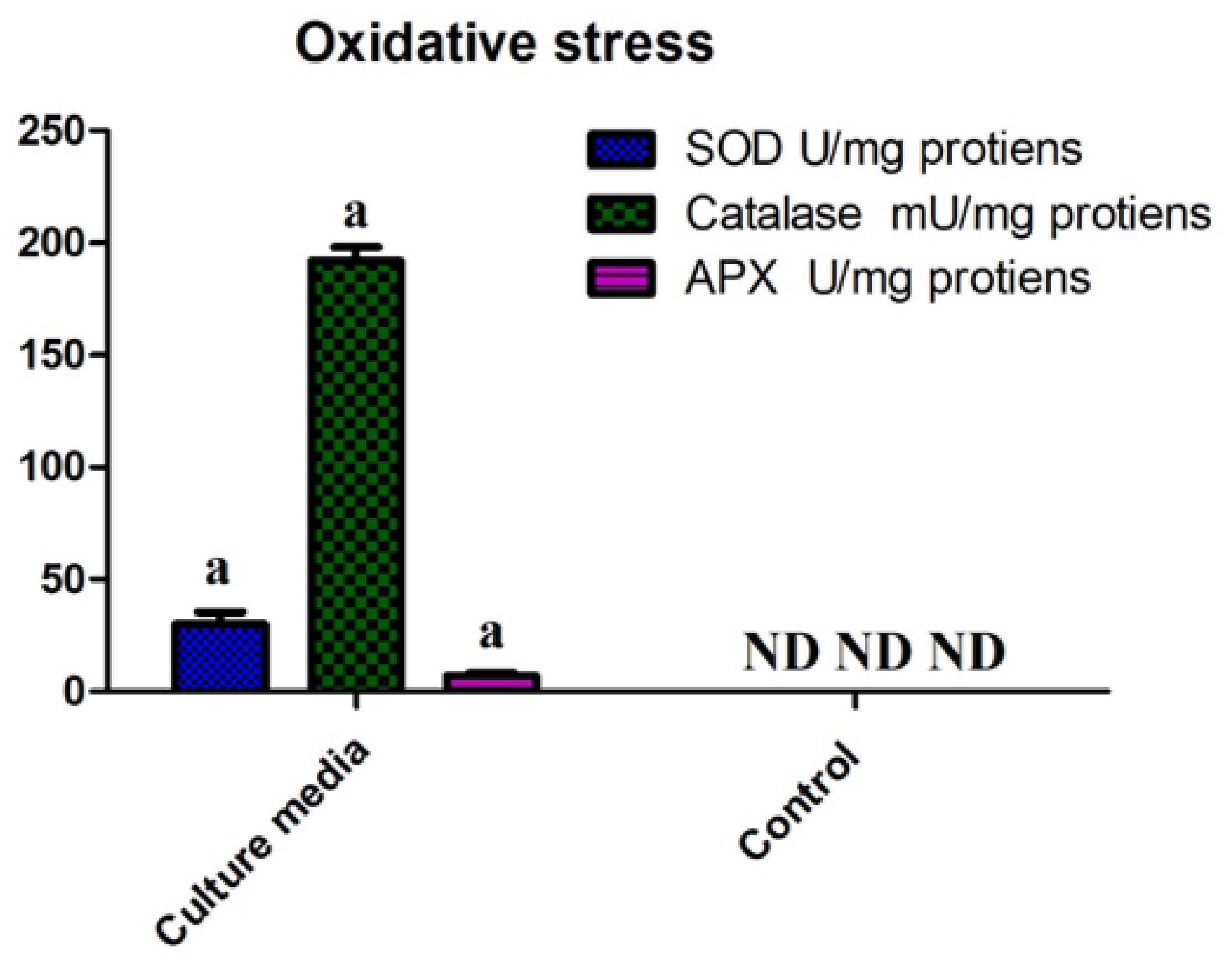
4.5. Oxidative Stress Tolerance Response
4.6. Effect of Seed Biopriming on Germination under Drought Tolerance
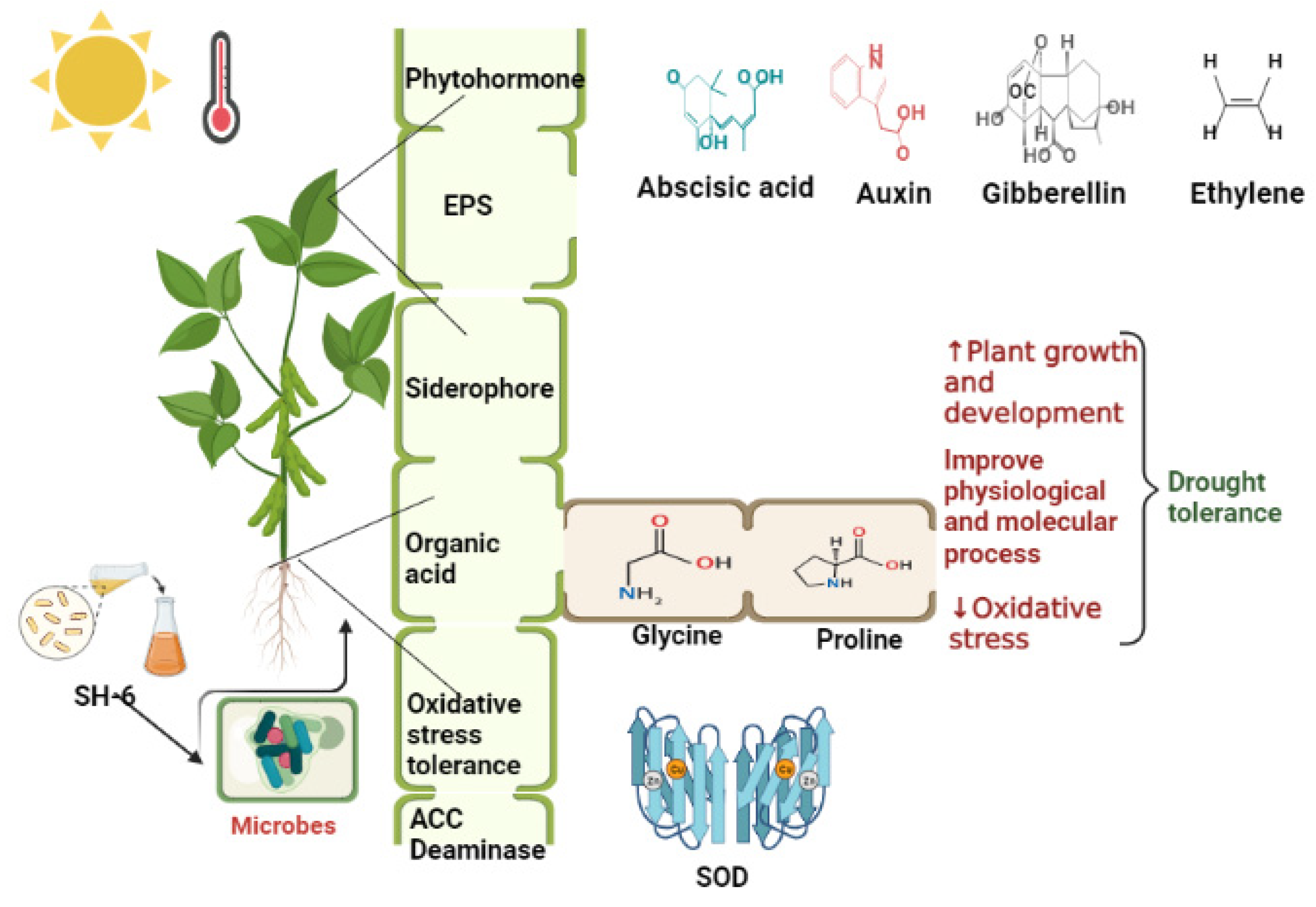
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siddique, K.H.; Johansen, C.; Turner, N.C.; Jeuffroy, M.-H.; Hashem, A.; Sakar, D.; Gan, Y.; Alghamdi, S.S. Innovations in agronomy for food legumes. A review. Agron. Sustain. Dev. 2012, 32, 45–64. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Toward a sustainable agriculture through plant biostimulants: From experimental data to practical applications. Agronomy 2020, 10, 1461. [Google Scholar] [CrossRef]
- Lichtfouse, E.; Navarrete, M.; Debaeke, P.; Souchère, V.; Alberola, C.; Ménassieu, J. Agronomy for sustainable agriculture: A review. Sustain. Agric. 2009, 29, 1–6. [Google Scholar] [CrossRef]
- Villalobos, F.J.; Fereres, E. Principles of Agronomy for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Brummer, E.C. Diversity, stability, and sustainable American agriculture. Agron. J. 1998, 90, J-17720. [Google Scholar] [CrossRef]
- Smil, V. Enriching the Earth: Fritz Haber, Carl Bosch, and the Transformation of World Food Production; MIT Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Dyson, T. World food trends and prospects to 2025. Proc. Natl. Acad. Sci. USA 1999, 96, 5929–5936. [Google Scholar] [CrossRef] [Green Version]
- Bisen, K.; Keswani, C.; Mishra, S.; Saxena, A.; Rakshit, A.; Singh, H. Unrealized potential of seed biopriming for versatile agriculture. In Nutrient Use Efficiency: From Basics to Advances; Springer: Berlin/Heidelberg, Germany, 2015; pp. 193–206. [Google Scholar]
- Singh, H. Seed biopriming: A comprehensive approach towards agricultural sustainability. Indian Phytopathol. 2016, 69, 203–209. [Google Scholar]
- Hossain, M.; Begum, M. Soil weed seed bank: Importance and management for sustainable crop production—A Review. J. Bangladesh Agric. Univ. 2015, 13, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Cardwell, V.B. Seed germination and crop production. Physiol. Basis Crop Growth Dev. 1984, 1, 53–92. [Google Scholar]
- Atkinson, R.J.; Thomas, E.; Roscioli, F.; Cornelius, J.P.; Zamora-Cristales, R.; Franco Chuaire, M.; Alcázar, C.; Mesén, F.; Lopez, H.; Ipinza, R. Seeding resilient restoration: An indicator system for the analysis of tree seed systems. Diversity 2021, 13, 367. [Google Scholar] [CrossRef]
- Vernooy, R.; Bessette, G.; Otieno, G. Resilient Seed Systems: Handbook; Bioversity International: Rome, Italy, 2019. [Google Scholar]
- Perotti, V.E.; Larran, A.S.; Palmieri, V.E.; Martinatto, A.K.; Permingeat, H.R. Herbicide resistant weeds: A call to integrate conventional agricultural practices, molecular biology knowledge and new technologies. Plant Sci. 2020, 290, 110255. [Google Scholar] [CrossRef]
- Kazimierczak, R.; Średnicka-Tober, D.; Hallmann, E.; Kopczyńska, K.; Zarzyńska, K. The impact of organic vs. conventional agricultural practices on selected quality features of eight potato cultivars. Agronomy 2019, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.P. Bio-priming of seeds. In Recent Advances in Crop Protection; Springer: Berlin/Heidelberg, Germany, 2012; pp. 8–90. [Google Scholar]
- Meena, S.K.; Rakshit, A.; Meena, V.S. Effect of seed bio-priming and N doses under varied soil type on nitrogen use efficiency (NUE) of wheat (Triticum aestivum L.) under greenhouse conditions. Biocatal. Agric. Biotechnol. 2016, 6, 68–75. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Mehralian, M. Seed bio-priming to improve germination, seedling growth and essential oil yield of Dracocephalum kotschyi Boiss, an endangered medicinal plant in Iran. Gesunde Pflanz. 2020, 72, 17–27. [Google Scholar] [CrossRef]
- Chin, J.M.; Lim, Y.Y.; Ting, A.S.Y. Biopolymers for biopriming of Brassica rapa seeds: A study on coating efficacy, bioagent viability and seed germination. J. Saudi Soc. Agric. Sci. 2021, 20, 198–207. [Google Scholar] [CrossRef]
- Moeinzadeh, A.; Sharif-Zadeh, F.; Ahmadzadeh, M.; Tajabadi, F.H. Biopriming of Sunflower (‘Helianthus annuus’ L.) Seed with’Pseudomonas fluorescens’ for Improvement of Seed Invigoration and Seedling Growth. Aust. J. Crop Sci. 2010, 4, 564–570. [Google Scholar]
- Rajendra Prasad, S.; Kamble, U.R.; Sripathy, K.; Udaya Bhaskar, K.; Singh, D. Seed bio-priming for biotic and abiotic stress management. In Microbial Inoculants in Sustainable Agricultural Productivity; Springer: Berlin/Heidelberg, Germany, 2016; pp. 211–228. [Google Scholar]
- Karthika, C.; Vanangamudi, K. Biopriming of maize hybrid COH (M) 5 seed with liquid biofertilizers for enhanced germination and vigour. Afr. J. Agric. Res. 2013, 8, 3310–3317. [Google Scholar]
- Piri, R.; Moradi, A.; Balouchi, H.; Salehi, A. Improvement of cumin (Cuminum cyminum) seed performance under drought stress by seed coating and biopriming. Sci. Hortic. 2019, 257, 108667. [Google Scholar] [CrossRef]
- Mahmood, A.; Turgay, O.C.; Farooq, M.; Hayat, R. Seed biopriming with plant growth promoting rhizobacteria: A review. FEMS Microbiol. Ecol. 2016, 92, fiw112. [Google Scholar] [CrossRef]
- Ali, L.G.; Nulit, R.; Ibrahim, M.H.; Yien, C.Y.S. Enhancement of germination and early seedling growth of rice (Oryza sativa) var. FARO44 by seed priming under normal and drought stressed conditions. J. Plant Nutr. 2020, 43, 1579–1593. [Google Scholar] [CrossRef]
- Manasa, P.; Maitra, S.; Reddy, M. Effect of summer maize-legume intercropping system on growth, productivity and competitive ability of crops. Int. J. Manag. Technol. Eng. 2018, 8, 2871–2875. [Google Scholar]
- Malhotra, S.K. Diversification in Utilization of Maize and Production. 2017, pp. 49–57. Available online: https://www.researchgate.net/publication/315045649_Diversification_in_Utilization_of_Maize_and_Production (accessed on 1 May 2022).
- Hanway, J.J.; Ritchie, S.W. Zea mays. In CRC Handbook of Flowering; CRC Press: Boca Raton, FL, USA, 2019; pp. 525–541. [Google Scholar]
- Strable, J.; Scanlon, M.J. Maize (Zea mays): A model organism for basic and applied research in plant biology. Cold Spring Harb. Protoc. 2009, 2009, pdb.emo132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnabás, B. Effect of water loss on germination ability of maize (Zea mays L.) pollen. Ann. Bot. 1985, 55, 201–204. [Google Scholar] [CrossRef]
- Thimme Gowda, P.; Manjunaththa, S.; Yogesh, T.; Satyareddi, S.A. Study on water requirement of Maize (Zea mays L.) using CROPWAT model in northern transitional zone of Karnataka. J. Environ. Sci. Comput. Sci. Eng. Technol. 2013, 2, 105–113. [Google Scholar]
- Steduto, P.; Hsiao, T.C.; Fereres, E.; Raes, D. Crop Yield Response to Water; Food and Agriculture Organization of the United Nations Rome: Rome, Italy, 2012; Volume 1028. [Google Scholar]
- Wang, H.; Liu, C. Experimental study on crop photosynthesis, transpiration and high efficient water use. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2003, 14, 1632–1636. [Google Scholar]
- Rahdari, P.; Hoseini, S. Drought stress: A review. Int. J. Agron. Plant Prod. 2012, 3, 443–446. [Google Scholar]
- Anjum, S.A.; Xie, X.-y.; Wang, L.-c.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Siddique, M.; Hamid, A.; Islam, M. Drought stress effects on water relations of wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Zand-Parsa, S.; Sepaskhah, A.; Ronaghi, A. Development and evaluation of integrated water and nitrogen model for maize. Agric. Water Manag. 2006, 81, 227–256. [Google Scholar] [CrossRef]
- Guo, S.; Wang, J.; Zhang, F.; Wang, Y.; Guo, P. An integrated water-saving and quality-guarantee uncertain programming approach for the optimal irrigation scheduling of seed maize in arid regions. Water 2018, 10, 908. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, A.C.; Savi, D.C.; Hungria, M.; Kava, V.; Glienke, C.; Galli-Terasawa, L.V. Bioprospecting of elite plant growth-promoting bacteria for the maize crop. Acta Scientiarum. Agron. 2020, 42, 1–11. [Google Scholar] [CrossRef]
- Mubeen, M.; Bano, A.; Ali, B.; Islam, Z.; Ahmad, A.; Hussain, S.; Fahad, S.; Nasim, W. Effect of plant growth promoting bacteria and drought on spring maize (Zea mays L.). Pak. J. Bot. 2021, 53, 731–739. [Google Scholar] [CrossRef]
- Trujillo-Reyes, J.; Majumdar, S.; Botez, C.; Peralta-Videa, J.; Gardea-Torresdey, J. Exposure studies of core–shell Fe/Fe3O4 and Cu/CuO NPs to lettuce (Lactuca sativa) plants: Are they a potential physiological and nutritional hazard? J. Hazard. Mater. 2014, 267, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.E.; Fischer, S.I.; Magris, S.; Mori, G.B. Isolation and characterization of bacteria from the rhizosphere of wheat. World J. Microbiol. Biotechnol. 2007, 23, 895–903. [Google Scholar] [CrossRef]
- Kim, Y.-N.; Khan, M.A.; Kang, S.-M.; Hamayun, M.; Lee, I.-J. Enhancement of drought-stress tolerance of Brassica oleracea var. italica L. by newly isolated Variovorax sp. YNA59. J. Microbiol. Biotechnol. 2020, 30, 1500–1509. [Google Scholar] [CrossRef]
- Kim, Y.-N.; Kang, S.-M.; Lee, K.-E.; Park, C.-W.; Park, H.-S.; Lee, I.-J. Improving Drought Stress Tolerance in Broccoli (Brassica oleracea var. italica L.) through the Soil Bacteria. Variovorax paradoxus YNA59 Treatment. Korean Hortic. Acad. Soc. 2019, 37, 41. [Google Scholar]
- Alam, S.; Khalil, S.; Ayub, N.; Rashid, M. In vitro solubilization of inorganic phosphate by phosphate solubilizing microorganisms (PSM) from maize rhizosphere. Int. J. Agric. Biol. 2002, 4, 454–458. [Google Scholar]
- Rashid, M.; Khalil, S.; Ayub, N.; Alam, S.; Latif, F. Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms (PSM) under in vitro conditions. Pak. J. Biol. Sci. 2004, 7, 187–196. [Google Scholar] [CrossRef]
- Alexander, D.; Zuberer, D. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Analysis of Indole-3-acetic Acid (IAA) Production in Klebsiellaby LC-MS/MS and the Salkowski Method. Bio-Protocol 2019, 9, e3230. [Google Scholar] [CrossRef]
- Platt Jr, R.S.; Thimann, K.V. Interference in Salkowski assay of indoleacetic acid. Science 1956, 123, 105–106. [Google Scholar] [CrossRef]
- Kang, S.; Sultana, T.; Loktev, V.B.; Wongratanacheewin, S.; Sohn, W.-M.; Eom, K.S.; Park, J.-K. Molecular identification and phylogenetic analysis of nuclear rDNA sequences among three opisthorchid liver fluke species (Opisthorchiidae: Trematoda). Parasitol. Int. 2008, 57, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.-G.; Li, S.-J.; Kang, Y.-X.; Liu, J.-J. Molecular characterization and expression analyses of an anthocyanin synthase gene from Magnolia sprengeri pamp. Appl. Biochem. Biotechnol. 2015, 175, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Halo, B.A.; Khan, A.L.; Waqas, M.; Al-Harrasi, A.; Hussain, J.; Ali, L.; Adnan, M.; Lee, I.-J. Endophytic bacteria (Sphingomonas sp. LK11) and gibberellin can improve Solanum lycopersicum growth and oxidative stress under Salinity. J. Plant Interact. 2015, 10, 117–125. [Google Scholar]
- Park, Y.-G.; Mun, B.-G.; Kang, S.-M.; Hussain, A.; Shahzad, R.; Seo, C.-W.; Kim, A.-Y.; Lee, S.-U.; Oh, K.Y.; Lee, D.Y. Bacillus aryabhattai SRB02 tolerates oxidative and nitrosative stress and promotes the growth of soybean by modulating the production of phytohormones. PLoS ONE 2017, 12, e0173203. [Google Scholar] [CrossRef] [Green Version]
- Netting, A.G.; Theobald, J.C.; Dodd, I.C. Xylem sap collection and extraction methodologies to determine in vivo concentrations of ABA and its bound forms by gas chromatography-mass spectrometry (GC-MS). Plant Methods 2012, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Verslues, P.E. Rapid quantification of abscisic acid by GC-MS/MS for studies of abiotic stress response. In Plant Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2017; pp. 325–335. [Google Scholar]
- Oyebanji, O.; Nweke, O.; Odebunmi, O.; Galadima, N.; Idris, M.; Nnodi, U.; Afolabi, A.; Ogbadu, G. Simple, effective and economical explant-surface sterilization protocol for cowpea, rice and sorghum seeds. Afr. J. Biotechnol. 2009, 8, 20. [Google Scholar]
- Ozkan, U. Determining germination responses of annual honeybee plant (Phacel a tanacet fol a Benth.) under salt and drought stress under in v tro conditions. Seeds 2021, 10, 100. [Google Scholar]
- Begum, N.; Wang, L.; Zhao, T. Effects of temperature regimes on seed germination and early growth of different soybean cultivars. Int. J. Appl. Exp. Biol. 2022, 1, 75–85. [Google Scholar]
- Siddique, K.; Belford, R.; Tennant, D. Root: Shoot ratios of old and modern, tall and semi-dwarf wheats in a Mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J. Maize. In Cereals; Springer: Berlin/Heidelberg, Germany, 2009; pp. 3–98. [Google Scholar]
- Dowswell, C.R.; Paliwal, R.L.; Cantrell, R.P. Maize in the Third World; CRC press: Boca Raton, FI, USA, 2019. [Google Scholar]
- Patidar, S.; Das, S.; Vishwakarma, R.; Kumari, P.; Mohanta, S.; Paradkar, V. Assessment of yield losses due to fall armyworm in maize. Pharma Innov. J. 2022, 11, 960–962. [Google Scholar]
- Chi, Y.X.; Gao, F.; Muhammad, I.; Huang, J.H.; Zhou, X.B. Effect of water conditions and nitrogen application on maize growth, carbon accumulation and metabolism of maize plant in subtropical regions. Arch. Agron. Soil Sci. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kour, D.; Abdel-Azeem, A.M.; Dikilitas, M.; Hesham, A.E.-L.; Ahluwalia, A.S. Microbes for Agricultural and Environmental Sustainability. J. Appl. Biol. Biotechnol. 2022, 10, 1–5. [Google Scholar] [CrossRef]
- Savari, M.; Damaneh, H.E.; Damaneh, H.E. Drought vulnerability assessment: Solution for risk alleviation and drought management among Iranian farmers. Int. J. Disaster Risk Reduct. 2022, 67, 102654. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Imran, M.; Kang, S.-M.; Park, Y.-S.; Wani, S.H.; Lee, I.-J. Research Progress in the Field of Microbial Mitigation of Drought Stress in Plants. Front. Plant Sci. 2022, 13, 870626. [Google Scholar] [CrossRef] [PubMed]
- Sharara, A.; Shekede, M.D.; Gwitira, I.; Masocha, M.; Dube, T. Fine-scale multi-temporal and spatial analysis of agricultural drought in agro-ecological regions of Zimbabwe. Geomat. Nat. Hazards Risk 2022, 13, 1342–1365. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.; Singh, N. Drought Tolerance in Rapeseed-Mustard: Conventional and Molecular Approaches. In Genomic Designing for Abiotic Stress Resistant Oilseed Crops; Springer: Berlin/Heidelberg, Germany, 2022; pp. 199–218. [Google Scholar]
- Tayebwa, O. Investigating the Use of Agrochemicals, Fumigants and Drug Concoctions in the Control of Acaricide-resistant Ticks in Kiruhura District. Ph.D. Thesis, Makerere University, Kampala, Uganda, 2022. [Google Scholar]
- Herrero, M.P.; Johnson, R. Drought stress and its effects on maize reproductive systems 1. Crop Sci. 1981, 21, 105–110. [Google Scholar] [CrossRef]
- Efeoğlu, B.; Ekmekçi, Y.; Çiçek, N. Physiological responses of three maize cultivars to drought stress and recovery. S. Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Mohammadkhani, N.; Heidari, R. Effects of drought stress on soluble proteins in two maize varieties. Turk. J. Biol. 2008, 32, 23–30. [Google Scholar]
- Negi, S.; Bharat, N.K. Seed Defense Biopriming. In Plant-Microbial Interactions and Smart Agricultural Biotechnology; CRC Press: Boca Raton, FI, USA, 2021; pp. 129–146. [Google Scholar]
- Sandini, I.E.; Pacentchuk, F.; Hungria, M.; Nogueira, M.A.; da Cruz, S.P.; Nakatani, A.S.; Araujo, R.S. Seed Inoculation with Pseudomonas fluorescens Promotes Growth, Yield and Reduces Nitrogen Application in Maize. Int. J. Agric. Biol. 2019, 22, 1369–1375. [Google Scholar]
- Sarkar, D.; Singh, S.; Parihar, M.; Rakshit, A. Seed bio-priming with microbial inoculants: A tailored approach towards improved crop performance, nutritional security, and agricultural sustainability for smallholder farmers. Curr. Res. Environ. Sustain. 2021, 3, 100093. [Google Scholar] [CrossRef]
- Ranathunge, N.P. Biopriming of Seeds for Plant Disease Tolerance and Phytopathogen Management. In Microbial Biocontrol: Sustainable Agriculture and Phytopathogen Management; Springer: Berlin/Heidelberg, Germany, 2022; pp. 355–369. [Google Scholar]
- Sood, M.; Kumar, V.; Rawal, R. Seed biopriming a novel method to control seed borne diseases of crops. In Biocontrol Agents and Secondary Metabolites; Elsevier: Amsterdam, The Netherlands, 2021; pp. 181–223. [Google Scholar]
- Yadav, R.S.; Singh, V.; Pal, S.; Meena, S.K.; Meena, V.S.; Sarma, B.K.; Singh, H.B.; Rakshit, A. Seed bio-priming of baby corn emerged as a viable strategy for reducing mineral fertilizer use and increasing productivity. Sci. Hortic. 2018, 241, 93–99. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Neal, A.L.; Ahmad, S.; Gordon-Weeks, R.; Ton, J. Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS ONE 2012, 7, e35498. [Google Scholar] [CrossRef] [Green Version]
- Becerra-Castro, C.; Monterroso, C.; Prieto-Fernández, A.; Rodríguez-Lamas, L.; Loureiro-Viñas, M.; Acea, M.; Kidd, P. Pseudometallophytes colonising Pb/Zn mine tailings: A description of the plant–microorganism–rhizosphere soil system and isolation of metal-tolerant bacteria. J. Hazard. Mater. 2012, 217, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, K.H.; Olsson, P.A.; Bååth, E. Structure and activity of the bacterial community in the rhizosphere of different plant species and the effect of arbuscular mycorrhizal colonisation. FEMS Microbiol. Ecol. 2002, 40, 223–231. [Google Scholar] [CrossRef]
- Watt, M.; Kirkegaard, J.; Passioura, J. Rhizosphere biology and crop productivity—A review. Soil Res. 2006, 44, 299–317. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Kopriva, S. Metabolic niches in the rhizosphere microbiome: New tools and approaches to analyse metabolic mechanisms of plant–microbe nutrient exchange. J. Exp. Bot. 2019, 70, 1087–1094. [Google Scholar] [CrossRef]
- Zenser, N.; Ellsmore, A.; Leasure, C.; Callis, J. Auxin modulates the degradation rate of Aux/IAA proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 11795–11800. [Google Scholar] [CrossRef] [Green Version]
- Raven, J. Transport of indoleacetic acid in plant cells in relation to pH and electrical potential gradients, and its significance for polar IAA transport. New Phytol. 1975, 74, 163–172. [Google Scholar] [CrossRef]
- Winkelmann, G. Microbial siderophore-mediated transport. Biochem. Soc. Trans. 2002, 30, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C. Siderophore mediated absorption of iron. Siderophores Microorg. Plants 1984, 58, 25–87. [Google Scholar]
- Finkelstein, R. Abscisic acid synthesis and response. Arab. Book/Am. Soc. Plant Biol. 2013, 11, e0166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.N.; Yadav, N. Stress-adaptive microbes for plant growth promotion and alleviation of drought stress in plants. Acta Sci. Agric. 2018, 2, 85–88. [Google Scholar]
- Singh, D.P.; Singh, V.; Gupta, V.K.; Shukla, R.; Prabha, R.; Sarma, B.K.; Patel, J.S. Microbial inoculation in rice regulates antioxidative reactions and defense related genes to mitigate drought stress. Sci. Rep. 2020, 10, 4818. [Google Scholar] [CrossRef]
- Dai, L.; Zhang, G.; Yu, Z.; Ding, H.; Xu, Y.; Zhang, Z. Effect of drought stress and developmental stages on microbial community structure and diversity in peanut rhizosphere soil. Int. J. Mol. Sci. 2019, 20, 2265. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Chakraborti, S.; Bera, K.; Sadhukhan, S.; Dutta, P. Bio-priming of seeds: Plant stress management and its underlying cellular, biochemical and molecular mechanisms. Plant Stress 2021, 3, 100052. [Google Scholar] [CrossRef]
- Hasan, M.M.; Gong, L.; Nie, Z.-F.; Li, F.-P.; Ahammed, G.J.; Fang, X.-W. ABA-induced stomatal movements in vascular plants during dehydration and rehydration. Environ. Exp. Bot. 2021, 186, 104436. [Google Scholar] [CrossRef]
- Bharath, P.; Gahir, S.; Raghavendra, A.S. Abscisic acid-induced stomatal closure: An important component of plant defense against abiotic and biotic stress. Front. Plant Sci. 2021, 12, 615114. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Groups | Fresh Biomass (mg) | Dry Biomass (mg) | Root Length cm | Hypocotyl Length cm |
|---|---|---|---|---|
| SH-6-M | 27.75 ± 0.06 a | 1.93 ± 0.08 a | 14 ± 0.05 a | 5.5 ± 0.01 a |
| SH-6-S | 20.96 ± 0.04 d | 1.91 ± 0.05 b | 0.5 ± 0.02 g | 4.2 ± 0.01 d |
| SH-6—(5%) PEG 6000 | 24.82 ± 0.02 b | 1.88 ± 0.07 c | 11 ± 0.02 b | 5.3 ± 0.03 b |
| SH-6—(10%) PEG 6000 | 21.27 ± 0.07 d | 1.78 ± 0.07 d | 10.7 ± 0.08 c | 5.2 ± 0.02 c |
| SH-6—(15%) PEG 6000 | 18.46 ± 0.08 e | 1.25 ± 0.03 f | 5.4 ± 0.01 e | 3.2 ± 0.02 e |
| SH-6—(20%) PEG 6000 | 16.96 ± 0.09 f | 1.33 ± 0.06 g | 7.9 ± 0.04 e | 1.9 ± 0.04 f |
| SH-6—(25%) PEG 6000 | 2.56 ± 0.09 g | 1.27 ± 0.08 h | 1.02 ± 0.04 f | 1.01 ± 0.06 g |
| Control | 21.34 ± 0.32 c | 1.62 ± 0.18 e | 9.69 ± 0.03 d | 4.55 ± 0.02 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaffique, S.; Khan, M.A.; Wani, S.H.; Imran, M.; Kang, S.-M.; Pande, A.; Adhikari, A.; Kwon, E.-H.; Lee, I.-J. Biopriming of Maize Seeds with a Novel Bacterial Strain SH-6 to Enhance Drought Tolerance in South Korea. Plants 2022, 11, 1674. https://doi.org/10.3390/plants11131674
Shaffique S, Khan MA, Wani SH, Imran M, Kang S-M, Pande A, Adhikari A, Kwon E-H, Lee I-J. Biopriming of Maize Seeds with a Novel Bacterial Strain SH-6 to Enhance Drought Tolerance in South Korea. Plants. 2022; 11(13):1674. https://doi.org/10.3390/plants11131674
Chicago/Turabian StyleShaffique, Shifa, Muhammad Aaqil Khan, Shabir Hussain Wani, Muhammad Imran, Sang-Mo Kang, Anjali Pande, Arjun Adhikari, Eun-Hae Kwon, and In-Jung Lee. 2022. "Biopriming of Maize Seeds with a Novel Bacterial Strain SH-6 to Enhance Drought Tolerance in South Korea" Plants 11, no. 13: 1674. https://doi.org/10.3390/plants11131674
APA StyleShaffique, S., Khan, M. A., Wani, S. H., Imran, M., Kang, S. -M., Pande, A., Adhikari, A., Kwon, E. -H., & Lee, I. -J. (2022). Biopriming of Maize Seeds with a Novel Bacterial Strain SH-6 to Enhance Drought Tolerance in South Korea. Plants, 11(13), 1674. https://doi.org/10.3390/plants11131674