
Comparative LC–LTQ–MS–MS Analysis of the Leaf Extracts of Lantana camara and Lantana montevidensis Growing in Egypt with Insights into Their Antioxidant, Anti-Inflammatory, and Cytotoxic Activities

 , ,
, , 
 , , ,
, , , 
Abstract
:1. Introduction
2. Results and Discussion
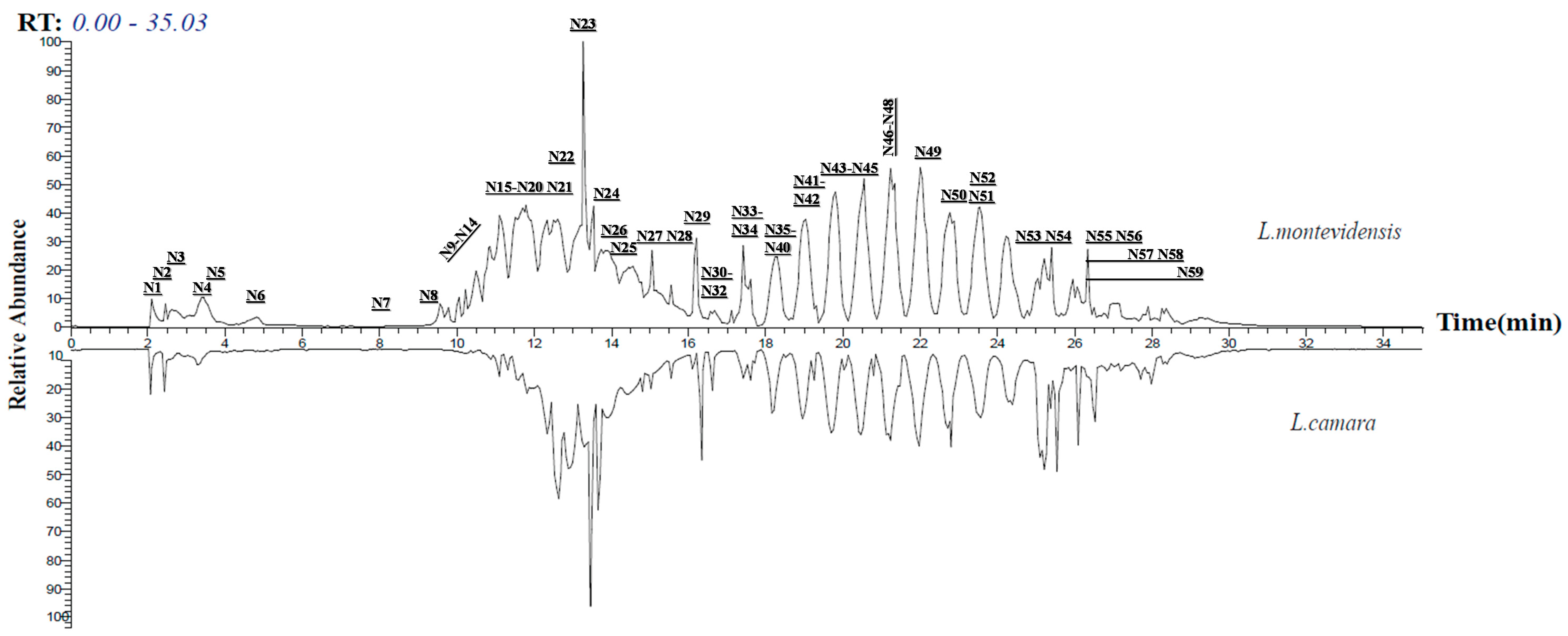
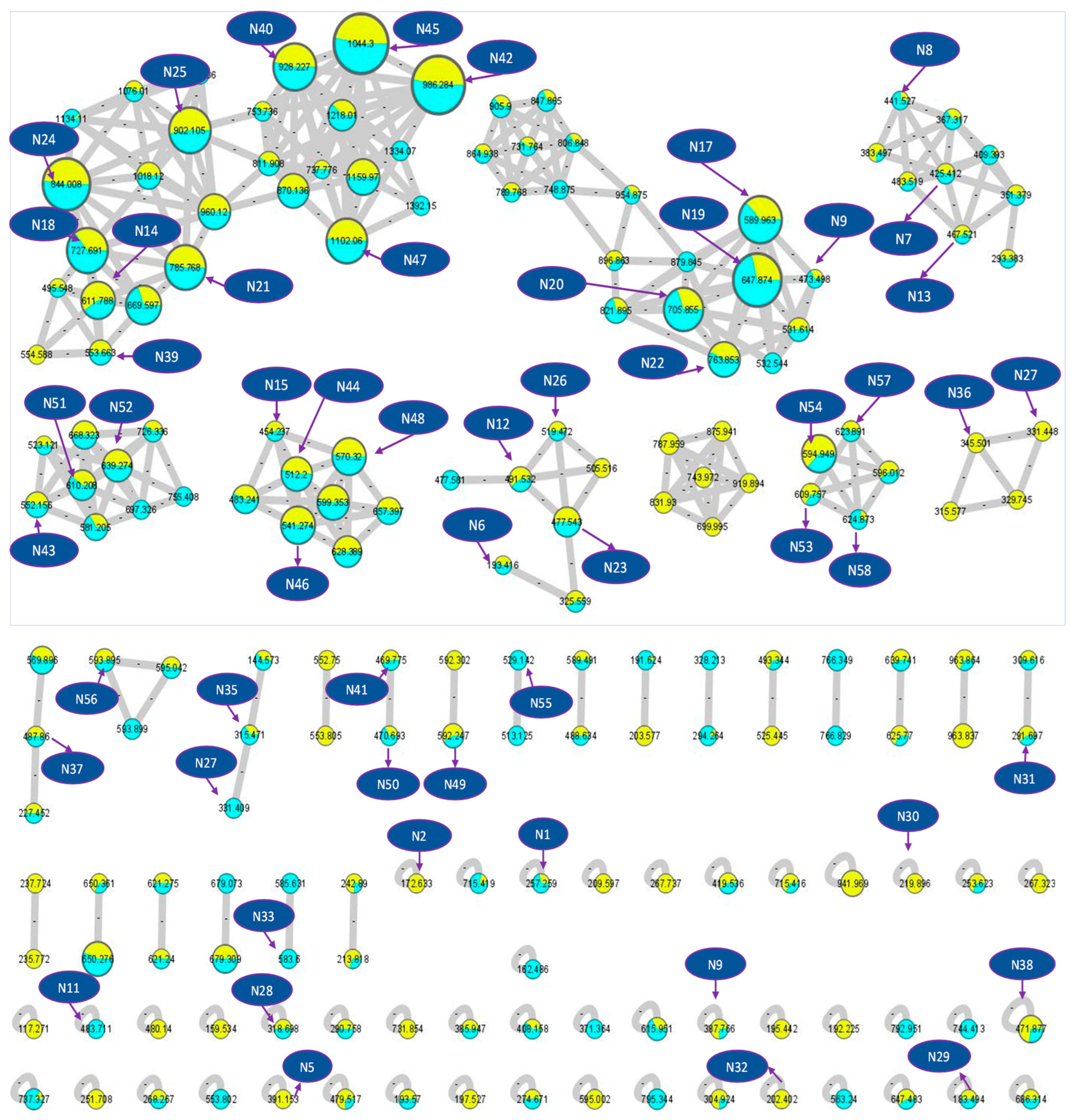
2.1. LC–LTQ–MS–MS Analysis and GNPS-Aided Annotation of L. camara and L. montevidensis Constituents
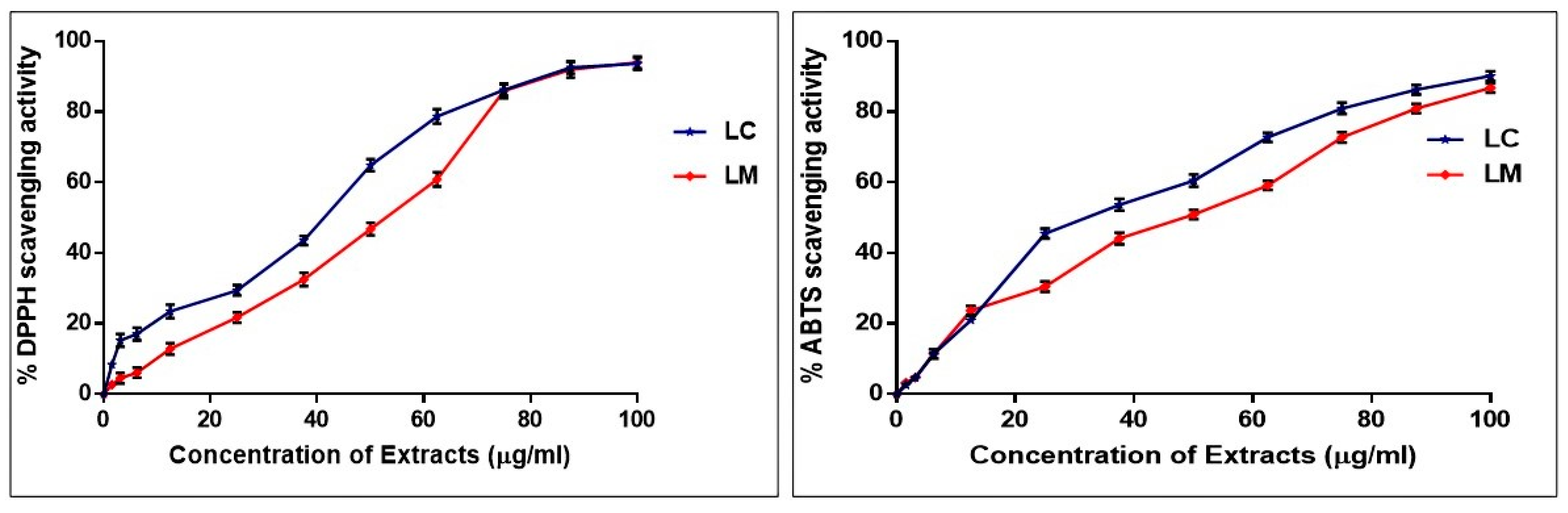
2.2. Assessment of the Antioxidant Effects of L. camara and L. montevidensis Extracts
2.2.1. DPPH• Assay
2.2.2. ABTS+ Assay
2.3. In Vitro Assessment of the Anti-Inflammatory Effects of L. camara and L. montevidensis
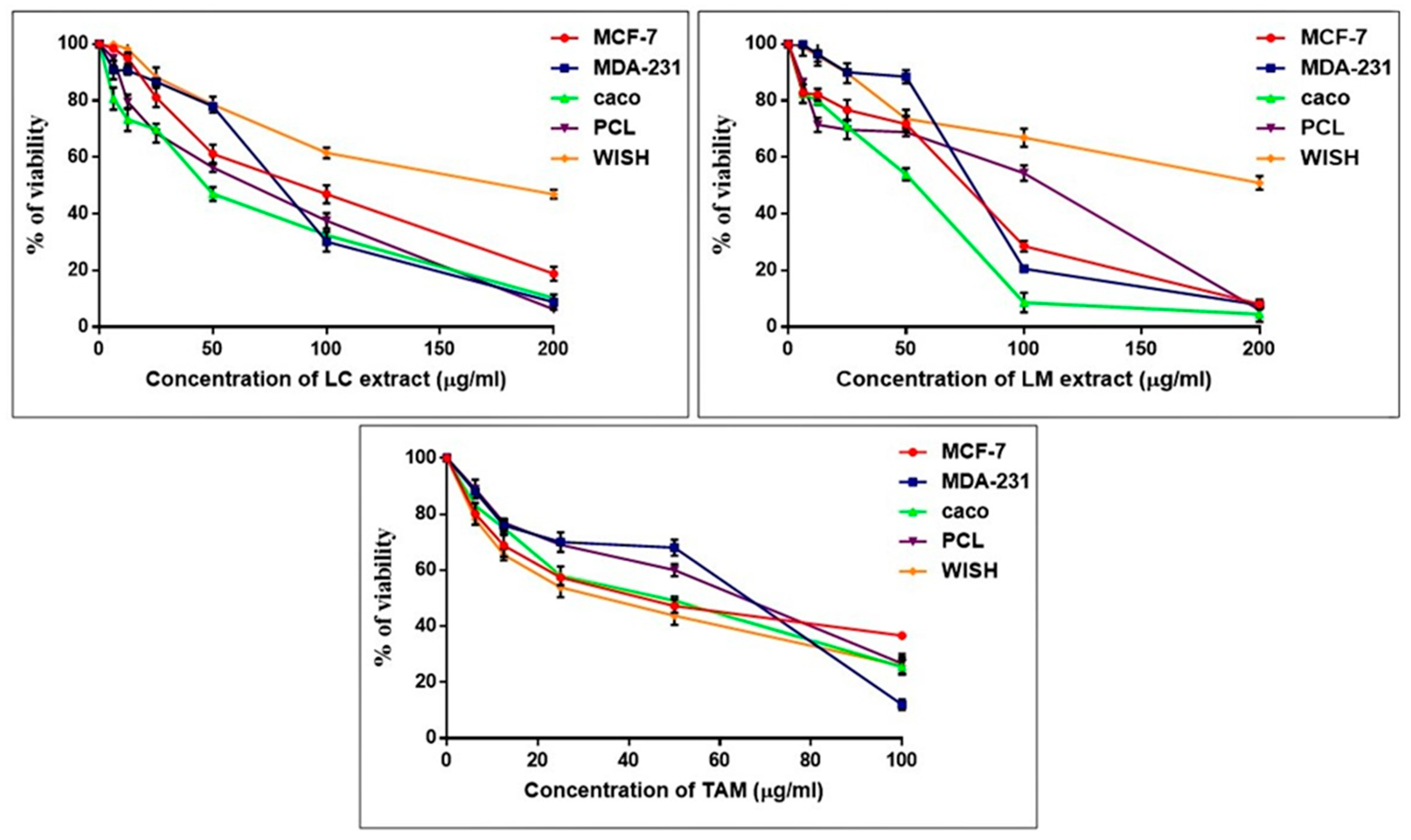
2.4. In Vitro Cytotoxicity Studies on L. camara and L. montevidensis Extracts
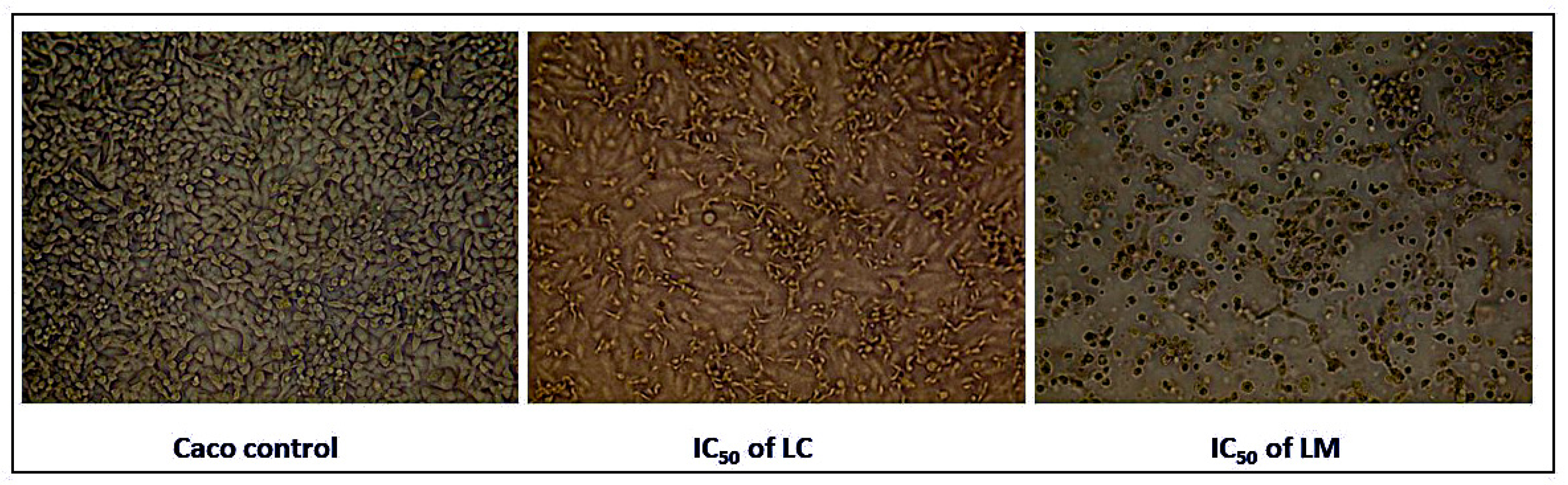
2.4.1. Alterations in Morphological Features of Treated Cancer Cells
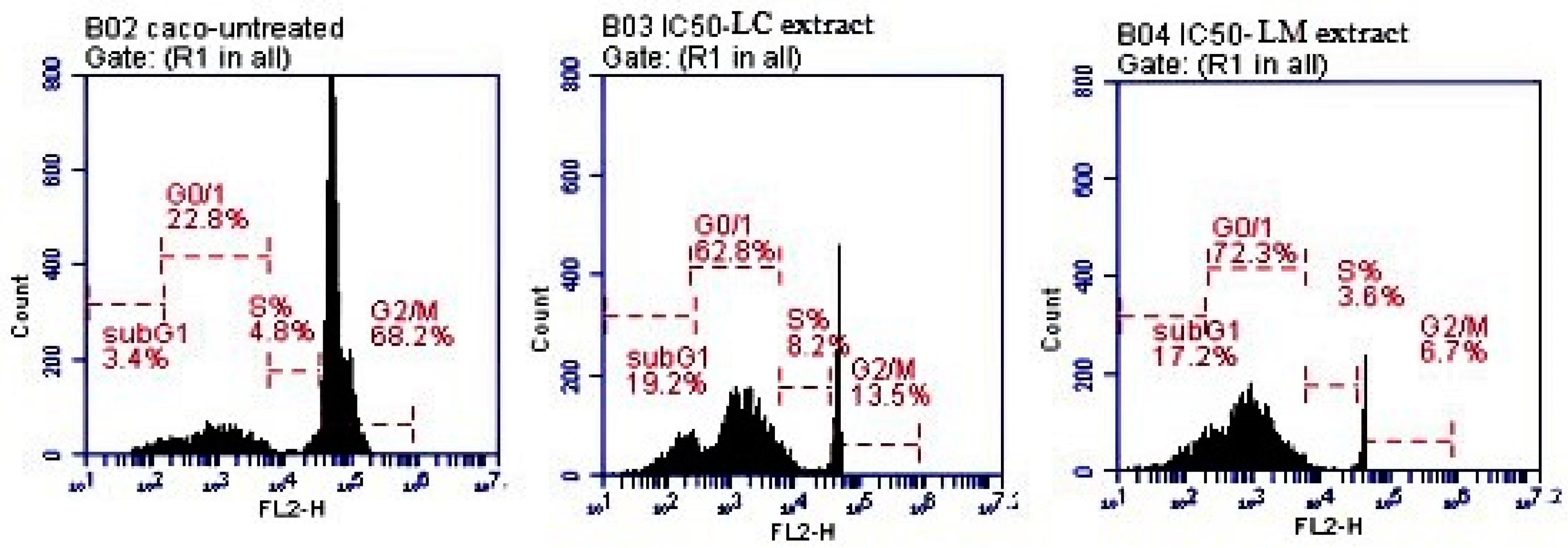
2.4.2. Analysis of the Cell Cycle
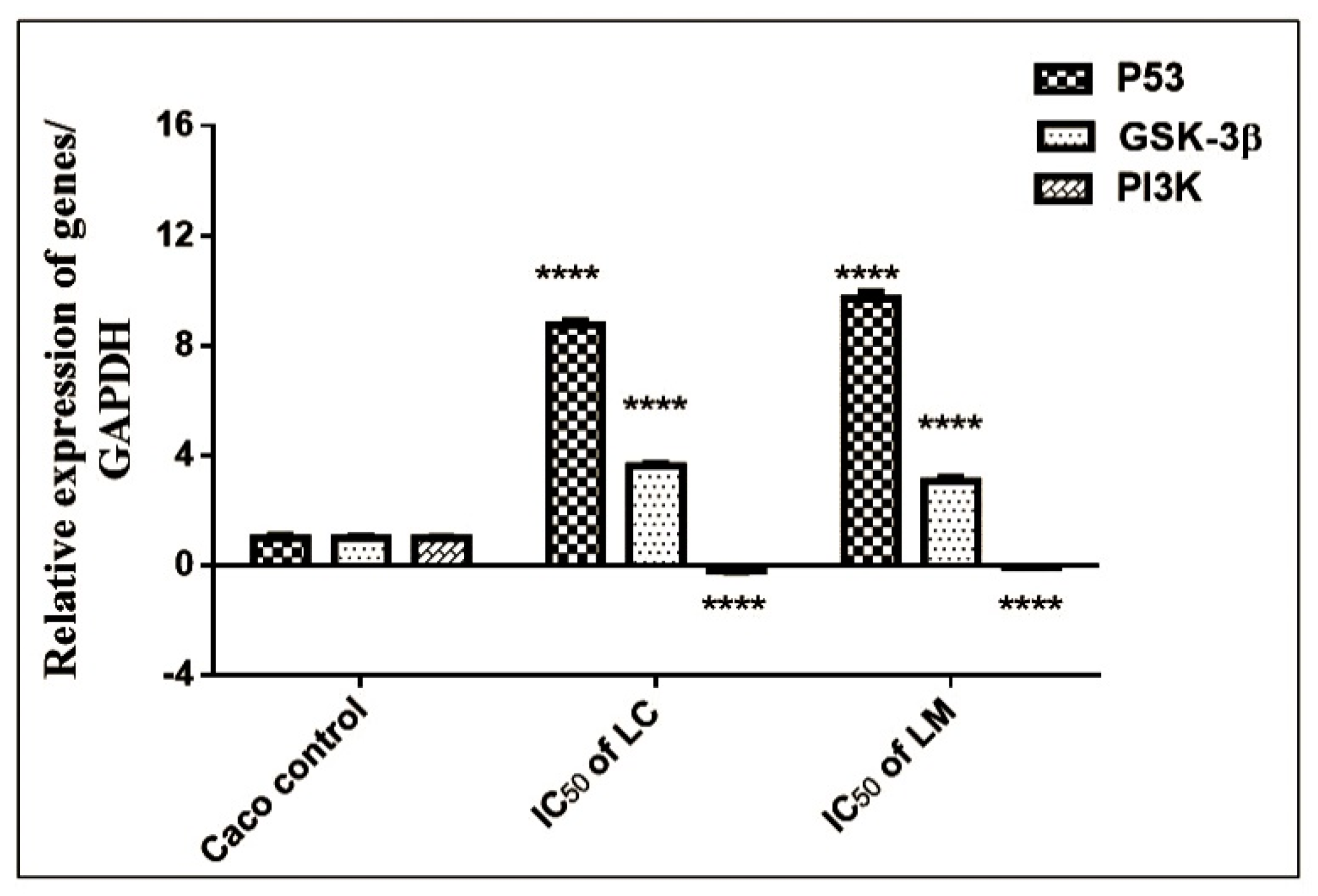
2.4.3. qRT-PCR Assessment
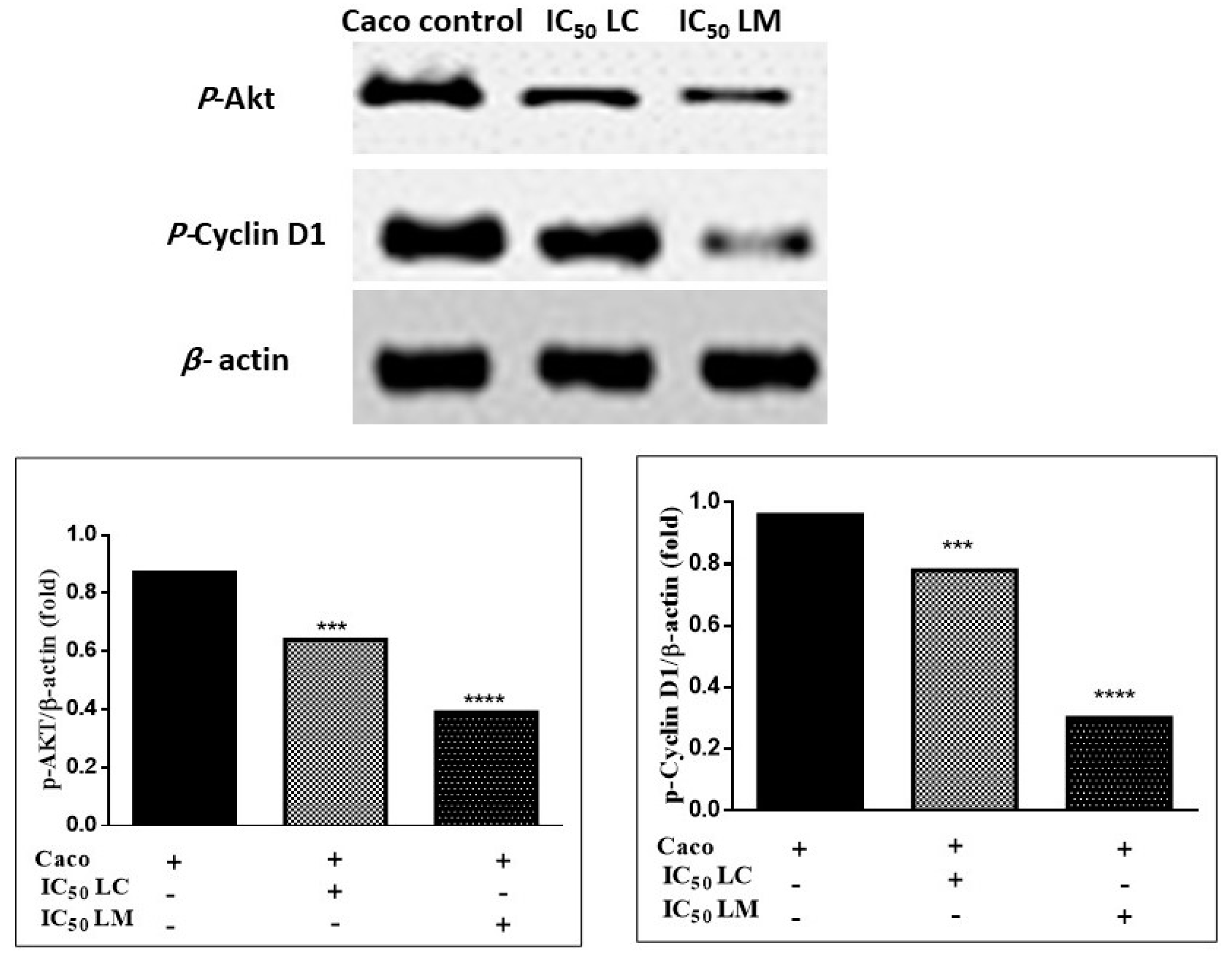
2.4.4. Immunoblotting Assay
3. Materials and Methods
3.1. Plant Collection and Extraction
3.2. LC–LTQ–MS–MS Analysis of L. camara and L. montevidensis Extracts
3.3. In Vitro Assessment of the Antioxidant Activities of L. camara and L. montevidensis Extracts
3.3.1. DPPH• Free Radical Scavenging
3.3.2. ABTS+ Radical Scavenging Activity
3.4. In Vitro Assessment of the Anti-Inflammatory Activity of L. camara and L. montevidensis
3.4.1. Preparation of Human Neutrophils
3.4.2. Lactate Dehydrogenase (LDH) Assay
3.4.3. Measurement of Superoxide Generation
3.4.4. Measurement of Elastase Release
3.5. In Vitro Cytotoxicity Investigation of L. camara and L. montevidensis Extracts
3.5.1. Cell Lines Maintenance and Treatment
3.5.2. Cell Morphology Study
3.5.3. Cell Cycle Examination
3.5.4. Quantitative Real-Time PCR (qRT-PCR)
3.5.5. Western Blot Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, G.P.; Raghubanshi, A.S.; Singh, J.S. Lantana invasion: An overview. Weed Biol. Manag. 2005, 5, 157–165. [Google Scholar] [CrossRef]
- Negi, G.C.S.; Sharma, S.; Vishvakarma, S.C.R.; Samant, S.S.; Maikhuri, R.K.; Prasad, R.C.; Palni, L.M.S. Ecology and Use of Lantana camara in India. Bot. Rev. 2019, 85, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Saxena, M.; Saxena, J.; Khare, S. A brief review on: Therapeutical values of Lantana camara plant. Int. J. Pharm. Life Sci. 2012, 3, 1551–1554. [Google Scholar]
- dos Santos Lencina, J.; Bonfa Moslaves, I.S.; de Araujo Isaias Muller, J.; Carvalho, R.; Amianti, C.; Bonfim, I.; Alves, F.M.; Carollo, C.A.; Candeloro, L.; dos Santos Júnior, A.A.; et al. Lantana canescens (Kunth) inhibits inflammatory and hyperalgesic responses in murine models. J. Ethnopharmacol. 2021, 280, 114461. [Google Scholar] [CrossRef]
- Dawood, A.S.; Chua, L.S.; Tan, T.S.; Alshemary, A.F. Apoptotic mechanism of lantadene A from Lantana camara leaves against prostatic cancer cells. Egypt. J. Chem. 2021, 64, 7503–7510. [Google Scholar] [CrossRef]
- Amany, R.; Yara, K.; Tharwat, R.; Gamal, H.; Khaulood, H. Multidrug-resistant Staphylococcus bacteria isolated from pregnant women and the antimicrobial effect of Lantana camara L. different extracts. Egypt. J. Exp. Biol. 2021, 17, 33. [Google Scholar]
- Qureshi, H.; Anwar, T.; Ali, Q.; Haider, M.Z.; Habib, N.; Fatima, S.; Waseem, M.; Bibi, Y.; Arshad, M.; Adkins, S.W. Isolation of natural herbicidal compound from Lantana camara. Int. J. Environ. Anal. Chem. 2021, 101, 631–638. [Google Scholar] [CrossRef]
- Edem, G.D.; Okon, K.A.; Essien, S.I.; Bassey, E.-O.I. Lantana camara: A potent influential factor in improving the gastric mucosa of wistar rats ravaged by ulcer. Biol. Clin. Sci. Res. J. 2021, 2021. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Sabti, M.Z.; Mousa, S.H. In vitro accumulation potentials of heavy metals in big-sage (Lantana camara L.) plant. DYSONA-Life Sci. 2021, 2, 12–17. [Google Scholar] [CrossRef]
- Mohamed, N.M.; Makboul, M.A.; Farag, S.F.; Tarawneh, A.H.; Khan, S.I.; Brooks, T.A.; Wang, Y.-H.; Ross, S.A. Iridoid and phenylpropanoid glycosides from the roots of Lantana montevidensis. Med. Chem. Res. 2017, 26, 1117–1126. [Google Scholar] [CrossRef]
- Sousa, E.O.; Rocha, J.B.T.; Barros, L.M.; Barros, A.R.C.; Costa, J.G.M. Phytochemical characterization and in vitro antioxidant properties of Lantana camara L. and Lantana montevidensis Briq. Ind. Crops Prod. 2013, 43, 517–522. [Google Scholar] [CrossRef]
- Barreto, F.S.; Sousa, E.O.; Rodrigues, F.F.G.; Costa, J.G.M.; Campos, A.R. Antibacterial Activity of Lantana camara Linn Lantana montevidensis Brig Extracts from Cariri-Ceara, Brazil. J. Young Pharm. 2010, 2, 42–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, N.M.; Makboul, M.A.; Farag, S.F.; Jain, S.; Jacob, M.R.; Tekwani, B.L.; Ross, S.A. Triterpenes from the roots of Lantana montevidensis with antiprotozoal activity. Phytochem. Lett. 2016, 15, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Hald, A.; Lotharius, J. Oxidative stress and inflammation in Parkinson’s disease: Is there a causal link? Exp. Neurol. 2005, 193, 279–290. [Google Scholar] [CrossRef] [PubMed]
- García, N.; Zazueta, C.; Aguilera-Aguirre, L. Oxidative Stress and Inflammation in Cardiovascular Disease. Oxidative Med. Cell. Longev. 2017, 2017, 5853238. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hong, M.; Tan, H.-Y.; Wang, N.; Feng, Y. Insights into the Role and Interdependence of Oxidative Stress and Inflammation in Liver Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 4234061. [Google Scholar] [CrossRef]
- Burgos-Morón, E.; Abad-Jiménez, Z.; Martínez de Marañón, A.; Iannantuoni, F.; Escribano-López, I.; López-Domènech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J. Clin. Med. 2019, 8, 1385. [Google Scholar] [CrossRef] [Green Version]
- Rivera, J.C.; Dabouz, R.; Noueihed, B.; Omri, S.; Tahiri, H.; Chemtob, S. Ischemic Retinopathies: Oxidative Stress and Inflammation. Oxidative Med. Cell. Longev. 2017, 2017, 3940241. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, K.; Hagio, T.; Matsuoka, S. The role of neutrophil elastase in acute lung injury. Eur. J. Pharmacol. 2002, 451, 1–10. [Google Scholar] [CrossRef]
- Fayez, S.; Ayoub, I.M.; Mostafa, N.M.; Moussa, A.Y.; Gamal El-Din, M.I.; El-Shazly, M. Nutraceuticals in Cancer Therapy. In Handbook of Oxidative Stress in Cancer: Therapeutic Aspects; Chakraborti, S., Ed.; Springer: Singapore, 2021; pp. 1–20. [Google Scholar]
- Thun, M.J.; DeLancey, J.O.; Center, M.M.; Jemal, A.; Ward, E.M. The global burden of cancer: Priorities for prevention. Carcinogenesis 2010, 31, 100–110. [Google Scholar] [CrossRef] [Green Version]
- von Meyenfeldt, M. Cancer-associated malnutrition: An introduction. Eur. J. Oncol. Nurs. 2005, 9, S35–S38. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-D.; Yuan, C.-F.; Bu, Y.-Q.; Wu, X.-M.; Wan, J.-Y.; Zhang, L.; Hu, N.; Liu, X.-J.; Zu, Y.; Liu, G.-L.; et al. Fangchinoline inhibits cell proliferation via Akt/GSK-3beta/cyclin D1 signaling and induces apoptosis in MDA-MB-231 breast cancer cells. Asian Pac. J. Cancer Prev. 2014, 15, 769–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Manning, B.D.; Cantley, L.C. Targeting the PI3K-Akt pathway in human cancer: Rationale and promise. Cancer Cell 2003, 4, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Yanaga, F.; Sasaguri, T. GSK-3β regulates cyclin D1 expression: A new target for chemotherapy. Cell. Signal. 2008, 20, 581–589. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; Burman, R.; Mansour, A.; Turki, Z.; Boulos, L.; Gullbo, J.; Goransson, U. The traditional medical uses and cytotoxic activities of sixty-one Egyptian plants: Discovery of an active cardiac glycoside from Urginea maritima. J. Ethnopharmacol. 2013, 145, 746–757. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; Yosri, N.; Khalifa, S.A.M.; Guo, Z.; Musharraf, S.G.; Xiao, J.; Saeed, A.; Du, M.; Khatib, A.; Abdel-Daim, M.M. Exploring natural products-based cancer therapeutics derived from egyptian flora. J. Ethnopharmacol. 2021, 269, 113626. [Google Scholar] [CrossRef]
- El-Garawani, I.M.; El-Sabbagh, S.M.; Abbas, N.H.; Ahmed, H.S.; Eissa, O.A.; Abo-Atya, D.M.; Khalifa, S.A.M.; El-Seedi, H.R. A newly isolated strain of Halomonas sp.(HA1) exerts anticancer potential via induction of apoptosis and G2/M arrest in hepatocellular carcinoma (HepG2) cell line. Sci. Rep. 2020, 10, 14076. [Google Scholar] [CrossRef]
- El-Garawani, I.; Hassab El-Nabi, S.; El Kattan, A.; Sallam, A.; Elballat, S.; Abou-Ghanima, S.; El Azab, I.H.; El-Seedi, H.R.; Am Khalifa, S.; El-Shamy, S. The ameliorative role of Acacia senegal gum against the oxidative stress and genotoxicity induced by the radiographic contrast medium (ioxitalamate) in albino rats. Antioxidants 2021, 10, 221. [Google Scholar] [CrossRef]
- Elrasoul, A.S.A.; Mousa, A.A.; Orabi, S.H.; Mohamed, M.A.E.-G.; Gad-Allah, S.M.; Almeer, R.; Abdel-Daim, M.M.; Khalifa, S.A.M.; El-Seedi, H.R.; Eldaim, M.A.A. Antioxidant, anti-inflammatory, and anti-apoptotic effects of Azolla pinnata ethanolic extract against lead-induced hepatotoxicity in rats. Antioxidants 2020, 9, 1014. [Google Scholar] [CrossRef]
- Sousa, E.O.; Miranda, C.M.; Nobre, C.B.; Boligon, A.A.; Athayde, M.L.; Costa, J.G. Phytochemical analysis and antioxidant activities of Lantana camara and Lantana montevidensis extracts. Ind. Crops Prod. 2015, 70, 7–15. [Google Scholar] [CrossRef]
- Sharma, O.P.; Sharma, S.; Pattabhi, V.; Mahato, S.B.; Sharma, P.D. A review of the hepatotoxic plant Lantana camara. Crit. Rev. Toxicol. 2007, 37, 313–352. [Google Scholar] [CrossRef] [PubMed]
- Cittadini, M.C.; García-Estévez, I.; Escribano-Bailón, M.T.; Rivas-Gonzalo, J.C.; Valentich, M.A.; Repossi, G.; Soria, E.A. Modulation of fatty acids and interleukin-6 in glioma cells by South American tea extracts and their phenolic compounds. Nutr. Cancer 2018, 70, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Tamma, R.V.; Nigg, H.N. HPLC identification of allelopathic compounds from Lantana camara. J. Chem. Ecol. 1989, 15, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, B.-E. A review of African medicinal and aromatic plants. Med. Aromat. Plants World-Afr. 2017, 3, 19–60. [Google Scholar]
- Begum, S.; Ahmed, M.; Siddiqui, B.S.; Khan, A.; Saify, Z.S.; Arif, M. Triterpenes, a sterol and a monocyclic alcohol from Momordica charantia. Phytochemistry 1997, 44, 1313–1320. [Google Scholar] [CrossRef]
- Lata, R.R. Extraction and Bioactivity of Organic Extracts of Lantana camara Leaves. Master’s Thesis, The University of the South Pacific, Suva, Fiji, 2020. [Google Scholar]
- Singh, S.K.; Tripathi, V.J.; Singh, R.H. 3β, 24-Dihydroxyolean-12-en-28-oic acid, a pentacyclic triterpene acid from Lantana indica. Phytochemistry 1990, 29, 3360–3362. [Google Scholar] [CrossRef]
- Shamsee, Z.R.; Al-Saffar, A.Z.; Al-Shanon, A.F.; Al-Obaidi, J.R. Cytotoxic and cell cycle arrest induction of pentacyclic triterpenoides separated from Lantana camara leaves against MCF-7 cell line in vitro. Mol. Biol. Rep. 2019, 46, 381–390. [Google Scholar] [CrossRef]
- Sousa, E.O.; Costa, J.G. Genus Lantana: Chemical aspects and biological activities. Rev. Bras. Farmacogn. 2012, 22, 1115–1180. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Li, Y.; Murakami, T.; Yamahara, J.; Yoshikawa, M. Protective effects of oleanolic acid oligoglycosides on ethanol- or indomethacin-induced gastric mucosal lesions in rats. Life Sci. 1998, 63, Pl245–Pl250. [Google Scholar] [CrossRef]
- de Sousa, E.O.; de Almeida, S.C.; Damasceno, S.S.; Nobre, C.B.; da Costa, J.G.M. Lantana camara L. and Lantana montevidensis (Spreng.) Briq. In Medicinal and Aromatic Plants of South America; Springer: Berlin/Heidelberg, Germany, 2018; pp. 275–288. [Google Scholar]
- Nagao, T.; Abe, F.; Kinjo, J.; Okabe, H. Antiproliferative constituents in plants 10. Flavones from the leaves of Lantana montevidensis B RIQ. and consideration of structure–activity relationship. Biol. Pharm. Bull. 2002, 25, 875–879. [Google Scholar] [CrossRef] [Green Version]
- Darwish, R.S.; El-Banna, A.A.; Ghareeb, D.A.; El-Hosseny, M.F.; Seadawy, M.G.; Dawood, H.M. Chemical profiling and unraveling of anti-COVID-19 biomarkers of red sage (Lantana camara L.) cultivars using UPLC-MS/MS coupled to chemometric analysis, in vitro study and molecular docking. J. Ethnopharmacol. 2022, 291, 115038. [Google Scholar] [CrossRef] [PubMed]
- Weyerstahl, P.; Wahlburg, H.C.; Marschall, H.; Rustaiyan, A. Terpenes and terpene derivatives, XXXII. New cadinene and bisabolene derivatives from the essential oil of Pulicaria gnaphalodes. Liebigs Ann. Chem. 1993, 1993, 1117–1123. [Google Scholar] [CrossRef]
- Rahma, N.A.; Rohman, A. UPLC MS/MS Profile and Antioxidant Activities from Nonpolar Fraction of Patiwala (Lantana camara) Leaves Extract. Separations 2022, 9, 75. [Google Scholar]
- Hart, N.; Lamberton, J.; Sioumis, A.; Suares, H. New triterpenes of Lantana camara. A comparative study of the constituents of several taxa. Aust. J. Chem. 1976, 29, 655–671. [Google Scholar] [CrossRef]
- Hussain, H.; Hussain, J.; Al-Harrasi, A.; Shinwari, Z.K. Chemistry of some species genus Lantana. Pak. J. Bot. 2011, 43, 51–62. [Google Scholar]
- Makboul, M.A.; Attia, A.A.; Farag, S.F.; Mohamed, N.M.; Ross, S.A. Chemical constituents with free-radical scavenging activity from the leaves of Lantana montevidensis (Spreng.) Briq. Phcog J. 2014, 6, 27–31. [Google Scholar] [CrossRef]
- Gaillard, P.; HautevIlle, M.; Picq, M.; Duclos, M.-C.; Dubois, M.; Prigent, A.-F. Selective inhibition of rat heart cAMP phosphodiesterases by lipophilic C-methyl-2-phenyl-4H-1-benzopyran-4-ones (C-methylflavones). Chem. Pharm. Bull. 1996, 44, 1571–1576. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Ayub, A.; Shaheen Siddiqui, B.; Fayyaz, S.; Kazi, F. Nematicidal triterpenoids from Lantana camara. Chem. Biodivers. 2015, 12, 1435–1442. [Google Scholar] [CrossRef]
- Begum, S.; Zehra, S.Q.; Siddiqui, B.S.; Fayyaz, S.; Ramzan, M. Pentacyclic triterpenoids from the aerial parts of Lantana camara and their nematicidal activity. Chem. Biodivers. 2008, 5, 1856–1866. [Google Scholar] [CrossRef]
- Begum, S.; Zehra, S.Q.; Wahab, A.; Siddiqui, B.S. Triterpenoidal secondary metabolites from Lantana camara Linn. Helv. Chim. Acta 2006, 89, 1932–1941. [Google Scholar] [CrossRef]
- Begum, S.; Ayub, A.; Qamar Zehra, S.; Shaheen Siddiqui, B.; Iqbal Choudhary, M. Leishmanicidal triterpenes from Lantana camara. Chem. Biodivers. 2014, 11, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hady, H.; El-Sayed, M.M.; Abdel-Hady, A.A.; Hashash, M.M.; Abdel-Hady, A.M.; Aboushousha, T.; Abdel-Hameed, E.-S.S.; Abdel-Lateef, E.E.-S.; Morsi, E.A. Nephroprotective Activity of methanolic extract of Lantana camara and squash (Cucurbita pepo) on cisplatin-induced nephrotoxicity in rats and identification of certain chemical constituents of Lantana camara by HPLC-ESI-MS. Pharmacogn. J. 2018, 10, 136–147. [Google Scholar] [CrossRef]
- Sharma, O.P.; Singh, A.; Sharma, S. Levels of lantadenes, bioactive pentacyclic triterpenoids, in young and mature leaves of Lantana camara var. aculeata. Fitoterapia 2000, 71, 487–491. [Google Scholar] [CrossRef]
- Siddiqui, B.S.; Raza, S.M.; Begum, S.; Siddiqui, S.; Firdous, S. Pentacyclic triterpenoids from Lantana camara. Phytochemistry 1995, 38, 681–685. [Google Scholar] [CrossRef]
- Wollenweber, E.; Dorr, M.; Muniappan, R.; Siems, K. Flavonoid aglycones and triterpenoids from the leaf exudate of Lantana camara and Lantana montevidensis. Biochem. Syst. Ecol. 1997, 25, 269–270. [Google Scholar] [CrossRef]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Casals, I.; Andrés-Lacueva, C.; Izquierdo-Pulido, M.; Lamuela-Raventós, R.M. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao). J. Mass Spectrom. 2003, 38, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, A.; Pati, S.; Minervini, F.; D’Antuono, I.; Linsalata, V.; Lattanzio, V. Verbascoside, isoverbascoside, and their derivatives recovered from olive mill wastewater as possible food antioxidants. J. Agric. Food Chem. 2012, 60, 1822–1829. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of cell viability by the lactate dehydrogenase assay. Cold Spring Harb. Protoc. 2018, 2018, pdb.prot095497. [Google Scholar] [CrossRef]
- Wu, P.; Song, Z.; Wang, X.; Li, Y.; Li, Y.; Cui, J.; Tuerhong, M.; Jin, D.-Q.; Abudukeremu, M.; Lee, D. Bioactive triterpenoids from Lantana camara showing anti-inflammatory activities in vitro and in vivo. Bioorg. Chem. 2020, 101, 104004. [Google Scholar] [CrossRef]
- Silva, T.; Suffredini, I.; Ricci, E.; Fernandes, S.; Gonçalves, V., Jr.; Romoff, P.; Lago, J.; Bernardi, M. Antinociceptive and anti-inflammatory effects of Lantana camara L. extract in mice. Rev. Bras. Plantas Med. 2015, 17, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Das Sarma, M.; Patra, A.; Hazra, B. Anti-inflammatory and anticancer compounds isolated from Ventilago madraspatana Gaertn., Rubia cordifolia Linn. and Lantana camara Linn. J. Pharm. Pharmacol. 2010, 62, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Yuting, C.; Rongliang, Z.; Zhongjian, J.; Yong, J. Flavonoids as superoxide scavengers and antioxidants. Free Radic. Biol. Med. 1990, 9, 19–21. [Google Scholar] [CrossRef]
- Xu, G.-H.; Kim, Y.-H.; Choo, S.-J.; Ryoo, I.-J.; Yoo, J.-K.; Ahn, J.-S.; Yoo, I.-D. Chemical constituents from the leaves of Ilex paraguariensis inhibit human neutrophil elastase. Arch. Pharmacal Res. 2009, 32, 1215–1220. [Google Scholar] [CrossRef]
- Deng, S.; Dai, G.; Chen, S.; Nie, Z.; Zhou, J.; Fang, H.; Peng, H.J.B. Dexamethasone induces osteoblast apoptosis through ROS-PI3K/AKT/GSK3β signaling pathway. Biomed. Pharmacother. 2019, 110, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Guo, Y.; Stacey, W.C.; Harwalkar, J.; Fretthold, J.; Hitomi, M.; Stacey, D.W. Glycogen synthase kinase 3 has a limited role in cell cycle regulation of cyclin D1 levels. BMC Cell Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Li, X.; Ho, C.-T.; Lin, X.; Zhang, Y.; Li, B.; Chen, Z. Cocoa tea (Camellia ptilophylla) induces mitochondria-dependent apoptosis in HCT116 cells via ROS generation and PI3K/Akt signaling pathway. Food Res. Int. 2020, 129, 108854. [Google Scholar] [CrossRef]
- MS-Convert. Available online: http://proteowizard.sourceforge.net/download.html (accessed on 1 June 2021).
- Burits, M.; Bucar, F. Antioxidant activity of Nigella sativa essential oil. Phytother. Res. 2000, 14, 323–328. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yang, S.-C.; Chung, P.-J.; Ho, C.-M.; Kuo, C.-Y.; Hung, M.-F.; Huang, Y.-T.; Chang, W.-Y.; Chang, Y.-W.; Chan, K.-H.; Hwang, T.-L. Propofol inhibits superoxide production, elastase release, and chemotaxis in formyl peptide–activated human neutrophils by blocking formyl peptide receptor 1. J. Immunol. 2013, 190, 6511–6519. [Google Scholar] [CrossRef] [Green Version]
- Bøyum, A.; Løvhaug, D.; Tresland, L.; Nordlie, E. Separation of leucocytes: Improved cell purity by fine adjustments of gradient medium density and osmolality. Scand. J. Immunol. 1991, 34, 697–712. [Google Scholar] [CrossRef]
- Korinek, M.; Hsieh, P.-S.; Chen, Y.-L.; Hsieh, P.-W.; Chang, S.-H.; Wu, Y.-H.; Hwang, T.-L. Randialic acid B and tomentosolic acid block formyl peptide receptor 1 in human neutrophils and attenuate psoriasis-like inflammation in vivo. Biochem. Pharmacol. 2021, 190, 114596. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological defense mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investig. 1973, 52, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Liaw, C.-C.; Chen, Y.-H.; Chang, W.-Y.; Chung, P.-J.; Hwang, T.-L. A novel immunomodulatory effect of ugonin U in human neutrophils via stimulation of phospholipase C. Free Radic. Biol. Med. 2014, 72, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.-L.; Leu, Y.-L.; Kao, S.-H.; Tang, M.-C.; Chang, H.-L. Viscolin, a new chalcone from Viscum coloratum, inhibits human neutrophil superoxide anion and elastase release via a cAMP-dependent pathway. Free Radic. Biol. Med. 2006, 41, 1433–1441. [Google Scholar] [CrossRef]
- Dash, S.K.; Ghosh, T.; Roy, S.; Chattopadhyay, S.; Das, D. Zinc sulfide nanoparticles selectively induce cytotoxic and genotoxic effects on leukemic cells: Involvement of reactive oxygen species and tumor necrosis factor alpha. J. Appl. Toxicol. 2014, 34, 1130–1144. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Halicka, H.D.; Zhao, H. Analysis of cellular DNA content by flow and laser scanning cytometry. Polyploidization Cancer 2010, 676, 137–147. [Google Scholar]
- Noser, A.A.; Abdelmonsef, A.H.; El-Naggar, M.; Salem, M.M. New Amino Acid Schiff Bases as Anticancer Agents via Potential Mitochondrial Complex I-Associated Hexokinase Inhibition and Targeting AMP-Protein Kinases/mTOR Signaling Pathway. Molecules 2021, 26, 5332. [Google Scholar] [CrossRef]
- Kvastad, L.; Werne Solnestam, B.; Johansson, E.; Nygren, A.; Laddach, N.; Sahlén, P.; Vickovic, S.; Bendigtsen, S.C.; Aaserud, M.; Floer, L.; et al. Single cell analysis of cancer cells using an improved RT-MLPA method has potential for cancer diagnosis and monitoring. Sci. Rep. 2015, 5, 16519. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. Enhanced chemiluminescence (ECL) for routine immunoblotting: An inexpensive alternative to commercially available kits. Spermatogenesis 2011, 1, 121–122. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Rt | Formula | m/z | MS2 | Relative Abundance | Chemical Class | Ref. | |
|---|---|---|---|---|---|---|---|---|---|
| Lc | Lm | ||||||||
| 1. | Palmitamide | 1.69 | C16H33NO | 257.2312 | 239.1328, 201.1358, 187.1268, 103.0043, 88.9936 | 11.84 | 4.58 | FA amide | GNPS |
| 2. | Gallic acid | 2.67 | C7H6O5 | 171.5321 | 152.9517, 85.9147 | 17.85 | 12.44 | Phenolic acid | [31] |
| 3. | α-Humulene | 2.76 | C15H24 | 202.3955 | 155.9594, 141.9675, 127, 9391 | 9.65 | 4.14 | Monocyclic sesquiterpene | GNPS |
| 4. | p-Coumaric acid | 3.73 | C9H8O3 | 166.5324 | 148.9633, 119.9389 | 5.95 | 3.97 | Phenolic acid | [32] |
| 5. | Theveside | 3.88 | C16H22O11 | 391.1981 | 373.0371, 355.0253, 279.0401, 228.9559, 210.9715, 192.9794, 148.935 | 7.15 | 6.09 | Iridoid | [33] |
| 6. | Ferulic acid | 4.82 | C10H10O4 | 193.3430 | 174.9286, 162.9635, 116.9951 | 15.32 | 14.58 | Phenolic acid | [34] |
| 7. | Lamiridoside | 8.01 | C17H26O12 | 425.3458 | 276.1333, 218.0855, 160.0576 | 12.52 | 12.14 | Iridoid | [35] |
| 8. | Momorodol | 9.84 | C26H49O5 | 441.5767 | 292.1333, 234.0912, 176.1231, 172.0572, 160.0817 | 9.09 | 6.65 | Triterpene | [36] |
| 9. | Pomolic acid | 10.01 | C30H48O4 | 473.4977 | 455.1928, 397.1318, 321.1171, 227.0642, 169.0692 | 11.08 | 3.49 | Triterpene | [13,37] |
| 10. | Coprostanone | 10.12 | C27H46O | 387.3893 | 369.0617, 355.1737, 351.2124, 313.1629, 269.1862 | 6.31 | 3.55 | Triterpene | GNPS |
| 11. | Dihydrixyolean-enoic acid (Hederagenin) | 10.12 | C30H49O4 | 473.5281 | 455.1928, 397.1318, 379.2358, 339.2030, 321.1171, 245.1248, 227.0642, 203.1588 | 11.08 | 8.26 | Triterpene | [37,38] |
| 12. | Carminic acid | 10.18 | C22H20O13 | 491.3225 | 315.1465, 300.1916, 159.1022 | 32.62 | 21.71 | Flavonoid | GNPS |
| 13. | Amyrin | 10.42 | C30H50O | 467.4973 | 334.1802, 276.1421, 218.0966, 160.013 | - | 6.39 | Triterpene | [38] |
| 14. | Rutin | 10.77 | C27H30O16 | 611.6093 | 553.2927, 477.2678, 317.1439, 301.124, 271.1095 | 45.86 | 39.98 | Flavonoid | [31] |
| 15. | Lantadene C | 11.32 | C35H54O5 | 553.61 | 535.2781, 525.3694, 495.2407, 477.2509, 401.1971, 301.1317 | 21.51 | 24.82 | Triterpene | [39] |
| 16. | Calceolarioside E | 11.88 | C23H26O11 | 479.4901 | 461.0790, 443.0788, 425.1241, 317.0097, 299.1252, 263.1182, 162.982 | 17.17 | 11.54 | Phenolic acid | [37] |
| 17. | Triterpene glycoside derivative | 12 | - | 589.7351 | 513.2347, 455.2542, 437.2231, 379.2318, 285.1443 | 57.74 | 73.83 | Triterpene glycoside | [39,40] |
| 18. | Triterpene glycoside derivative | 12.4 | - | 727.4208 | 709.4453, 669.3836, 670.4085, 651.4137, 611.3975, 593.3708 | 56.84 | 73.05 | Triterpene glycoside | |
| 19. | Triterpene glycoside derivative | 12.5 | - | 647.7673 | 571.2557, 513.2347, 455.2542, 437.2231, 379.2318, 285.1443 | 73.45 | 100 | Triterpene glycoside | [41] |
| 20. | Triterpene glycoside derivative | 12.8 | - | 705.8645 | 571.2820, 513.2588, 629.2966, 437.2466, 495.2869, 455.2817, 285.1443 | 55.65 | 75.45 | Triterpene glycoside | [39,40] |
| 21. | Triterpene glycoside derivative | 13.4 | - | 785.4552 | 727.4422, 709.4453, 669.3836, 670.4085, 651.4137, 611.3975, 593.3708, | 66.14 | 67.99 | Triterpene glycoside | |
| 22. | Durantoside | 13.13 | C35H40O19 | 763.9281 | 687.3165, 629.3169, 571.0399, 513.3507, 437.2202, 285.1336 | 39.41 | 52.87 | Iridoid | [40] |
| 23. | Dihydroxy-dimethoxyflavone-O-glucopyranoside (Camaroside) | 13.45 | C23H24O11 | 477.4684 | 459.1729, 357.1318, 315.2007, 301.1745 | 47.17 | 36.08 | Flavonoid | [37] |
| 24. | Triterpene glycoside derivative | 14.27 | - | 843.9122 | 785.4807, 767.5064, 727.5031, 709.4918 | 54.35 | 54.97 | Triterpene glycoside | |
| 25. | Triterpene glycoside derivative | 14.83 | - | 901.9307 | 543.5972, 825.6067, 767.5857, 709.5566 | 39.59 | 38.59 | Triterpene glycoside | |
| 26. | Lantanoside | 14.68 | C25H26O12 | 519.6086 | 459.1729, 357.1318, 315.2007, 301.1745, | 10.89 | 10.75 | Flavonoid | [42] |
| 27. | Cirsiliol/ Trihydroxy-dimethoxyflavone | 15.05 | C17H14O7 | 331.4301 | 316.1003, 285.1243, 271.1618, 151.052 | 14.30 | 17.36 | Flavonoid | [43,44] |
| 28. | Hexahydroxyflavone (Gossypetin) | 15.07 | C15H10O8 | 318.6483 | 283.10, 242.90, 183.05, 169, 156.90, 109, 96.92 | 6.03 | 4.86 | Flavonoid | [37] |
| 29. | Caffeic acid | 16.26 | C9H9O4 | 181.5156 | 162.9605, 135.0327, 107.0433, 59.0440 | 23.17 | 16.72 | Phenolic acid | [44] |
| 30. | Copaenol | 16.44 | C15H25O | 221.7963 | 203.0473, 175.0733, 161.0492 | 13.81 | 5.39 | Sesquiterpene | [45] |
| 31. | Catechin | 16.81 | C15H15O6 | 291.6731 | 273.0806, 255.1193, 217.0495, 147.0402 | 12.79 | 9.06 | Flavonoid | [46] |
| 32. | Methyl-hydroxylantanolate | 16.84 | C31H49O | 500.7488 | 482.2634, 469.2448, 401.2497, 317.1457 | 5.21 | 3.48 | Triterpeme | [47] |
| 33. | Ursangilic acid | 17.06 | C36H54O6 | 583.6300 | 565.2075, 485.2668, 467.3068, 449.361 | 6.46 | 8.04 | Triterpene | [40,48] |
| 34. | Benzalkonium chloride | 17.65 | C21H38N+ | 304.8513 | 212.1338, 90.9176 | 3.6 | - | Ammonium Compound | GNPS |
| 35. | Dihydroxy-dimethoxyflavone (Pectolinarigenin) | 18.05 | C17H14O6 | 315.5381 | 300.0404, 282.0015, 269.0966, 121.0294 | 26.17 | 22.87 | Flavonoid | [37,49] |
| 36. | Dihydroxy-trimethoxyflavone | 18.20 | C18H17O7 | 345.4969 | 330.1257, 313.1737, 285.16, 151.0042 | 17.83 | 4.92 | Flavonoid | [50] |
| 37. | Lantaninilic acid/Lantoic acid | 18.50 | C30H46O5 | 487.6703 | 469.2092, 451.2662, 433.2741, 405.2914, 259.1011 | 18.75 | 10.55 | Triterpene | [37,51] |
| 38. | Camarin | 18.62 | C30H46O4 | 471.865 | 451.2739, 433.3931, 423.2796, 405.3678, 395.294, 313.2488, 271.192 | 46.93 | 15.27 | Triterpene | [52,53] |
| 39. | Stigmasterol acetate | 18.77 | C31H50O2 | 454.2024 | 328.1359, 299.1358. 270.1251, 241.083, 211.9450, 182.9712 | 16.83 | 18.26 | Triterpene | [35] |
| 40. | Triterpene glycoside derivative | 18.99 | - | 927.9218 | 851.6035, 793.5519, 735.5273, 677.4890, 635.4446 | 28.27 | 34.97 | Triterpene | |
| 41. | Pomonic acid | 19.24 | C30H46O4 | 469.6669 | 451.2739, 395.294, 313.2488 | 20.68 | 17.92 | Triterpene | [37] |
| 42. | Triterpene glycoside derivative | 19.80 | - | 985.9156 | 909.6604, 851.6151, 793.5680, 735.5282, 677.4925 | 35.37 | 43.41 | Triterpene | |
| 43. | Lantadene A | 20.02 | C35H52O5 | 552.7491 | 524.3465, 506.4606, 478.4638, 316.2659 | 26.88 | 31.96 | Triterpene | [54] |
| 44. | Lantanone | 20.14 | C32H48O5 | 512.3184 | 482.4986, 425.2136, 357.3943, 328.3304, 299.2942, 270.1839 | 5.81 | 47.77 | Triterpene | [37] |
| 45. | Lablaboside derivative | 20.51 | - | 1043.9023 | 941.7347, 793.5754, 647.5658, 473.4372, 389.2796, 331.2671 | 37.08 | 45.57 | Triterpene glycoside | |
| 46. | Lantadene D | 21.21 | C34H52O5 | 541.2581 | 511.6045, 425.2182, 357.3688, 328.3118, 299.2821, 270.2123, 241.122 | 6.53 | 56.70 | Triterpene | [39] |
| 47. | Lablaboside A | 21.26 | C54H87O23 | 1102.8948 | 941.7347, 793.5754, 647.5658, 473.4372, 389.2796, 331.2671 | 37.39 | 47.76 | Triterpene glycosides | [41] |
| 48. | Icterogenin/Lantacin | 21.64 | C35H52O6 | 570.2877 | 551.2625, 451.2778, 405.2828, 357.3562, 299.2313, 241.1121 | 59.43 | 67.93 | Triterpene | [39,40,52] |
| 49. | Osmanthuside B | 22.33 | C29H36O13 | 592.267 | 574.2937, 524.5654, 447.2928, 389.3079, 331.2425, 273.1571 | 30.85 | 32.81 | Phenolic acid | [55] |
| 50. | Hydroxyoleanonic acid/Lantabetulic acid | 22.78 | C30H48O4 | 470.0257 | 452.2639, 434.261, 396.2762, 307.0825 | - | 17.92 | Triterpene | [37,42,56] |
| 51. | Isonuomioside A | 23.01 | C28H34O15 | 610.2413 | 591.4889, 531.5561, 447.2902, 389.2357, 339. 1718, 243.0484 | 49.29 | 48.15 | Phenolic acid | [42] |
| 52. | Cistanoside C | 23.31 | C30H38O15 | 639.2013 | 621.2913, 552.4019, 505.355, 447.2967 | 44.41 | 44.59 | Phenolic acid | [55] |
| 53. | Lipedoside A | 25.42 | C29H36O14 | 609.6940 | 591.3207, 559.3793, 531.3948, 515.344 | 49.29 | 48.15 | Phenolic acid | [55] |
| 54. | Apigenin-6,8-di-C-glycoside (Vicenin 2) | 25.48 | C27H30O15 | 594.9509 | 576.4872, 534.3077, 474.3816, 642.376, 317.2311, 236.2413 | 100 | 54.74 | Flavonoid | [55] |
| 55. | Camarinic acid | 26.57 | C35H62O3 | 529.1427 | 283.2026, 256.3454, 246.3058, 242.3309, 163.1626, 149.1549 | 15.61 | 6.3 | Triterpene | [57] |
| 56. | Pheophorbide A | 26.59 | C35H36N4O5 | 593.9152 | 533.391, 473.4104, 461.4372, 433.4519 | 30.61 | 54.74 | Chlorophyll derivative | [35] |
| 57. | Pectolinarigenin-O-rutinoside (Pectolinarin) | 27.88 | C29H34O15 | 623.886 | 605.3220, 545.3717, 459.3893, 395.3564, 367.3008 | 14.18 | 9.35 | Flavonoid | [37] |
| 58. | Verbascoside/Forsythoside A | 27.89 | C29H36O15 | 624.8313 | 606.2446, 546.3597, 397.3595, 284.3636, 266.3377 | 9.16 | 6.87 | Phenolic acid | [52,58] |
| 59. | Vanillic acid | 31.12 | C12H6O4 | 169.8434 | 150.9395, 140.9568, 123.0144, 108.9833 | 16.99 | 12.36 | Phenolic acid | [44] |
| Extract | Cell Viability (%) |
|---|---|
| L. camara | 95.88 ± 4.60 |
| L. montevidensis | 97.11 ± 2.89 |
| Extract | IC50 a | Inhibition% (1 μg/mL) | Inhibition% (3 μg/mL) | Inhibition% (10 μg/mL) |
|---|---|---|---|---|
| L. camara | 1.57 ± 0.19 μg/mL | 24.80 ± 4.53 ** | 91.94 ± 4. 90 *** | 100.49 ± 1.14 *** |
| L. montevidensis | 1.31 ± 0.14 μg/mL | 29.90 ± 4.28 *** | 97.70 ± 0.26 *** | 100.92 ± 0.29 *** |
| LY294002 b | 2.41 ± 0.26 μM |
| Extract | IC50 a | Inhibition% (1 μg/mL) | Inhibition% (3 μg/mL) | Inhibition% (10 μg/mL) |
|---|---|---|---|---|
| L. camara | 2.40 ± 0.16 μg/mL | 9.95 ± 2.32 * | 64.22 ± 6.33 *** | 109.24 ± 5.15 *** |
| L. montevidensis | 1.90 ± 0.07 μg/mL | 16.68 ± 0.04 *** | 80.82 ± 4.68 *** | 115.84 ± 2.02 *** |
| LY294002 b | 3.18 ± 0.57 μM |
| Gene | Forward Primer (/5–/3) | Reverse Primer (/5–/3) |
|---|---|---|
| p53 | TAACAGTTCCTGCATGGGCGGC | AGGACAGGCACAAACACGCACC |
| GSK-3β | CCGACTAACACCACTGGAAGCT | AGGATGGTAGCCAGAGGTGGAT |
| PI3K | GCTCTCTCACTGCATACATTGT | AGTCACAGCTGTATTGGTCG |
| GAPDH | TGTGTCCGTCGTGGATCTGA | CCTGCTTCACCACCTTCTTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Din, M.I.G.; Fahmy, N.M.; Wu, F.; Salem, M.M.; Khattab, O.M.; El-Seedi, H.R.; Korinek, M.; Hwang, T.-L.; Osman, A.K.; El-Shazly, M.; et al. Comparative LC–LTQ–MS–MS Analysis of the Leaf Extracts of Lantana camara and Lantana montevidensis Growing in Egypt with Insights into Their Antioxidant, Anti-Inflammatory, and Cytotoxic Activities. Plants 2022, 11, 1699. https://doi.org/10.3390/plants11131699
El-Din MIG, Fahmy NM, Wu F, Salem MM, Khattab OM, El-Seedi HR, Korinek M, Hwang T-L, Osman AK, El-Shazly M, et al. Comparative LC–LTQ–MS–MS Analysis of the Leaf Extracts of Lantana camara and Lantana montevidensis Growing in Egypt with Insights into Their Antioxidant, Anti-Inflammatory, and Cytotoxic Activities. Plants. 2022; 11(13):1699. https://doi.org/10.3390/plants11131699
Chicago/Turabian StyleEl-Din, Mariam I. Gamal, Nouran M. Fahmy, Fulin Wu, Maha M. Salem, Omar M. Khattab, Hesham R. El-Seedi, Michal Korinek, Tsong-Long Hwang, Ahmed K. Osman, Mohamed El-Shazly, and et al. 2022. "Comparative LC–LTQ–MS–MS Analysis of the Leaf Extracts of Lantana camara and Lantana montevidensis Growing in Egypt with Insights into Their Antioxidant, Anti-Inflammatory, and Cytotoxic Activities" Plants 11, no. 13: 1699. https://doi.org/10.3390/plants11131699
APA StyleEl-Din, M. I. G., Fahmy, N. M., Wu, F., Salem, M. M., Khattab, O. M., El-Seedi, H. R., Korinek, M., Hwang, T. -L., Osman, A. K., El-Shazly, M., & Fayez, S. (2022). Comparative LC–LTQ–MS–MS Analysis of the Leaf Extracts of Lantana camara and Lantana montevidensis Growing in Egypt with Insights into Their Antioxidant, Anti-Inflammatory, and Cytotoxic Activities. Plants, 11(13), 1699. https://doi.org/10.3390/plants11131699