


Soil–Plant Relationships in Soybean Cultivated under Crop Rotation after 17 Years of No-Tillage and Occasional Chiseling

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
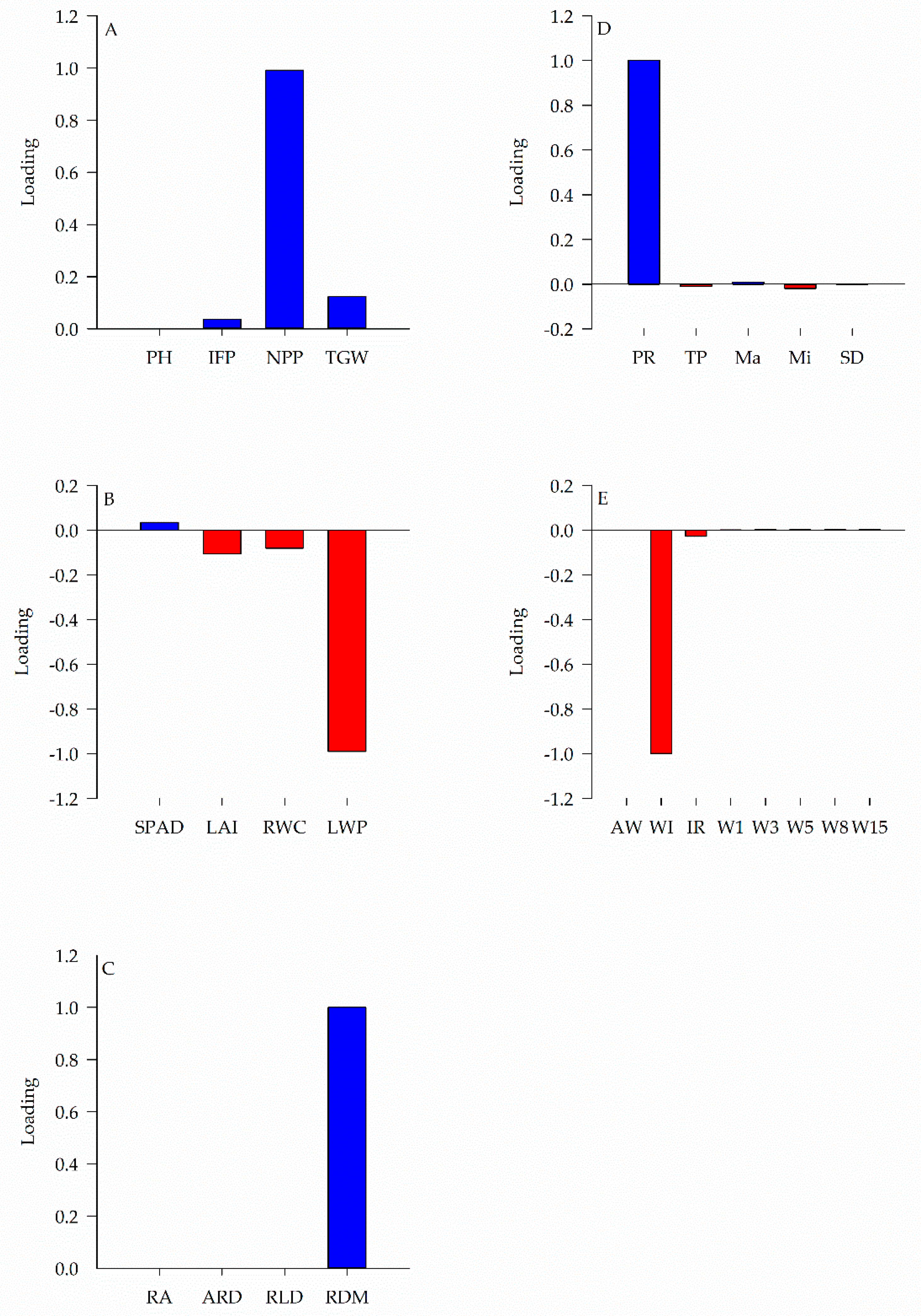
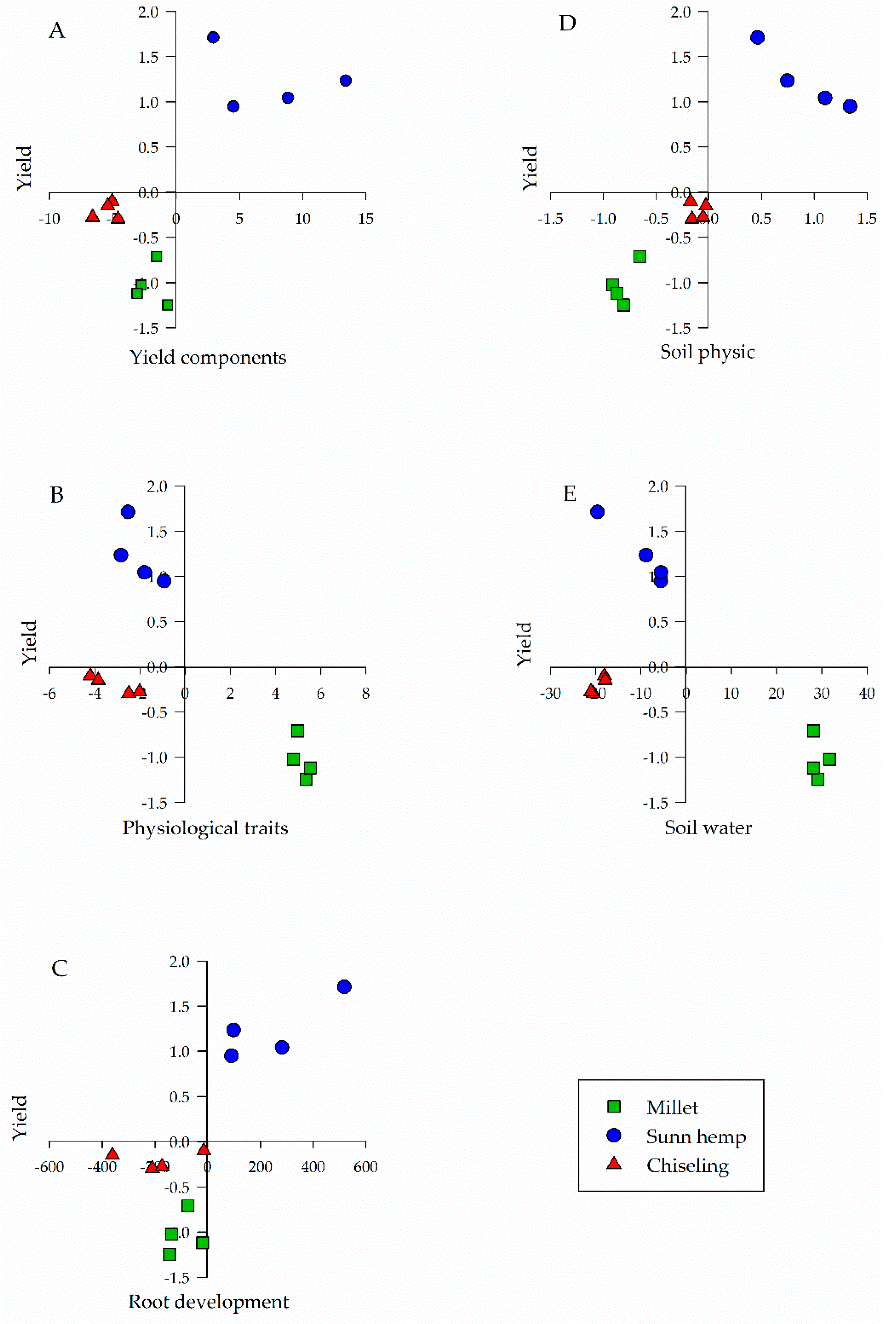
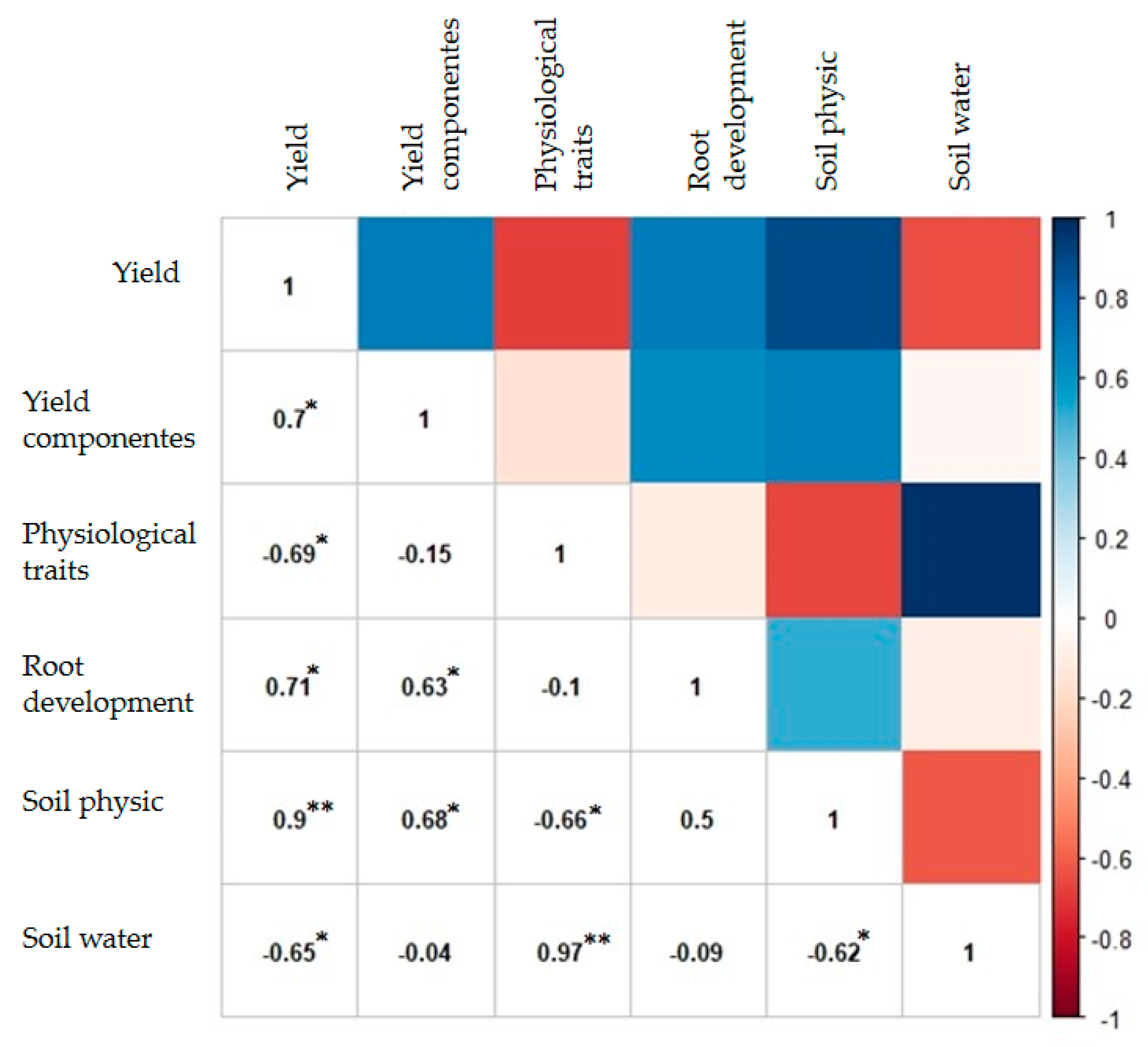
2. Results
3. Discussion
4. Materials and Methods
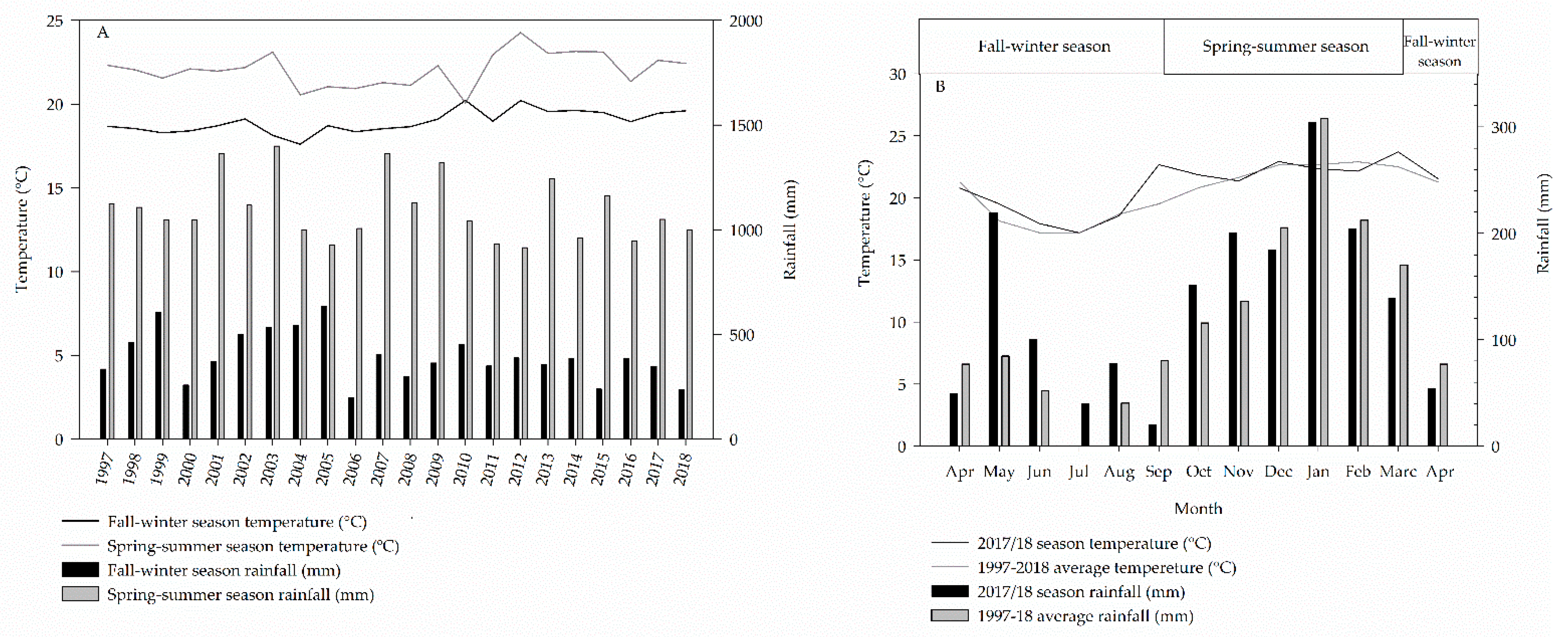
4.1. Site Description
4.2. History and Experimental Design
4.3. Management and Analysis of Soybean Plant
4.3.1. Physiological Traits
4.3.2. Root Development
4.3.3. Yield Components
4.3.4. Yield
4.4. Soil Analysis
4.4.1. Soil Water Storage
4.4.2. Soil Physical Properties
4.4.3. Infiltration and Rate of Water Infiltration into the Soil
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palm, C.; Blanco-Canqui, H.; DeClerck, F.; Gatere, L.; Grace, P. Conservation agriculture and ecosystem services: An overview. Agric. Ecosyst. Environ. 2014, 187, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Canqui, H.; Ruis, S.J. No-tillage and soil physical environment. Geoderma 2018, 326, 164–200. [Google Scholar] [CrossRef]
- Deuschle, D.; Minella, J.P.G.; Hörbe, T.D.A.N.; Londero, A.L.; Schneider, F.J.A. Erosion and hydrological response in no-tillage subjected to crop rotation intensification in southern Brazil. Geoderma 2019, 340, 157–163. [Google Scholar] [CrossRef]
- Ferreira, C.J.B.; Tormena, C.A.; Severiano, E.D.C.; Zotarelli, L.; Betioli Júnior, E. Soil compaction influences soil physical quality and soybean yield under long-term no-tillage. Arch. Agron. Soil Sci. 2021, 67, 383–396. [Google Scholar] [CrossRef]
- Zhang, Z.; Peng, X. Bio-tillage: A new perspective for sustainable agriculture. Soil Tillage Res. 2021, 206, 104844. [Google Scholar] [CrossRef]
- Elkins, C.B. Plant roots as tillage tools. J. Terramechanics 1985, 22, 177–178. [Google Scholar] [CrossRef]
- Chen, G.; Weil, R.R. Penetration of cover crop roots through compacted soils. Plant Soil 2010, 331, 31–43. [Google Scholar] [CrossRef]
- Calonego, J.C.; Raphael, J.P.A.; Rigon, J.P.G.; de Oliveira Neto, L.; Rosolem, C.A. Soil compaction management and soybean yields with cover crops under no-till and occasional chiseling. Eur. J. Agron. 2017, 85, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Cresswell, H.; Kirkegaard, J. Subsoil amelioration by plant-roots—The process and the evidence. Soil Res. 1995, 33, 221. [Google Scholar] [CrossRef]
- Yunusa, I.A.M.; Newton, P.J. Plants for amelioration of subsoil constraints and hydrological control: The primer-plant concept. Plant Soil 2003, 257, 261–281. [Google Scholar] [CrossRef]
- Kautz, T.; Lüsebrink, M.; Pätzold, S.; Vetterlein, D.; Pude, R.; Athmann, M.; Küpper, P.M.; Perkons, U.; Köpke, U. Contribution of anecic earthworms to biopore formation during cultivation of perennial ley crops. Pedobiologia 2014, 57, 47–52. [Google Scholar] [CrossRef]
- Han, E.; Kautz, T.; Perkons, U.; Lüsebrink, M.; Pude, R.; Köpke, U. Quantification of soil biopore density after perennial fodder cropping. Plant Soil 2015, 394, 73–85. [Google Scholar] [CrossRef]
- Naveed, M.; Moldrup, P.; Schaap, M.G.; Tuller, M.; Kulkarni, R.; Vogel, H.-J.; Wollesen de Jonge, L. Prediction of biopore- and matrix-dominated flow from X-ray CT-derived macropore network characteristics. Hydrol. Earth Syst. Sci. 2016, 20, 4017–4030. [Google Scholar] [CrossRef] [Green Version]
- Galdos, M.V.; Pires, L.F.; Cooper, H.V.; Calonego, J.C.; Rosolem, C.A.; Mooney, S.J. Assessing the long-term effects of zero-tillage on the macroporosity of Brazilian soils using X-ray Computed Tomography. Geoderma 2019, 337, 1126–1135. [Google Scholar] [CrossRef]
- Zhang, D.-W.; Xiao, Z.-J.; Zeng, B.-P.; Li, K.; Tang, Y.-L. Insect Behavior and Physiological Adaptation Mechanisms Under Starvation Stress. Front. Physiol. 2019, 10, 163. [Google Scholar] [CrossRef]
- Pfeifer, J.; Kirchgessner, N.; Walter, A. Artificial pores attract barley roots and can reduce artifacts of pot experiments. J. Plant Nutr. Soil Sci. 2014, 177, 903–913. [Google Scholar] [CrossRef]
- Colombi, T.; Braun, S.; Keller, T.; Walter, A. Artificial macropores attract crop roots and enhance plant productivity on compacted soils. Sci. Total Environ. 2017, 574, 1283–1293. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, T.M.; de Moraes Sá, J.C.; Tormena, C.A.; Dranski, A.; Muchalak, A.; Briedis, C.; de Oliveira Ferreira, A.; Giarola, N.F.B.; da Silva, Á.P. Mechanical and biological chiseling impacts on soil organic C stocks, root growth, and crop yield in a long-term no-till system. Soil Tillage Res. 2021, 211, 104993. [Google Scholar] [CrossRef]
- Guedes Filho, O.; da Silva, A.P.; Giarola, N.F.B.; Tormena, C.A. Structural properties of the soil seedbed submitted to mechanical and biological chiseling under no-tillage. Geoderma 2013, 204–205, 94–101. [Google Scholar] [CrossRef]
- Nunes, M.R.; Denardin, J.E.; Pauletto, E.A.; Faganello, A.; Pinto, L.F.S. Effect of soil chiseling on soil structure and root growth for a clayey soil under no-tillage. Geoderma 2015, 259–260, 149–155. [Google Scholar] [CrossRef]
- de Moraes, M.T.; Debiasi, H.; Carlesso, R.; Franchini, J.C.; da Silva, V.R. Critical limits of soil penetration resistance in a rhodic Eutrudox. Rev. Bras. Ciência Do Solo 2014, 38, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Canqui, H.; Wortmann, C.S. Does occasional tillage undo the ecosystem services gained with no-till? A review. Soil Tillage Res. 2020, 198, 104534. [Google Scholar] [CrossRef]
- Abrahão, G.M.; Costa, M.H. Evolution of rain and photoperiod limitations on the soybean growing season in Brazil: The rise (and possible fall) of double-cropping systems. Agric. For. Meteorol. 2018, 256–257, 32–45. [Google Scholar] [CrossRef]
- da Silva, G.F.; Calonego, J.C.; Luperini, B.C.O.; Chamma, L.; Alves, E.R.; Rodrigues, S.A.; Putti, F.F.; da Silva, V.M.; de Almeida Silva, M. Soil—Plant Relationships in Soybean Cultivated under Conventional Tillage and Long-Term No-Tillage. Agronomy 2022, 12, 697. [Google Scholar] [CrossRef]
- Hubert, F.; Hallaire, V.; Sardini, P.; Caner, L.; Heddadj, D. Pore morphology changes under tillage and no-tillage practices. Geoderma 2007, 142, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Osunbitan, J.A.; Oyedele, D.J.; Adekalu, K.O. Tillage effects on bulk density, hydraulic conductivity and strength of a loamy sand soil in southwestern Nigeria. Soil Tillage Res. 2005, 82, 57–64. [Google Scholar] [CrossRef]
- Deiss, L.; Kleina, G.B.; Moraes, A.; Franzluebbers, A.J.; Motta, A.C.V.; Dieckow, J.; Sandini, I.E.; Anghinoni, I.; Carvalho, P.C.F. Soil chemical properties under no-tillage as affected by agricultural trophic complexity. Eur. J. Soil Sci. 2019, 71, 1090–1105. [Google Scholar] [CrossRef]
- Sokolowski, A.C.; Prack McCormick, B.; De Grazia, J.; Wolski, J.E.; Rodríguez, H.A.; Rodríguez-Frers, E.P.; Gagey, M.C.; Debelis, S.P.; Paladino, I.R.; Barrios, M.B. Tillage and no-tillage effects on physical and chemical properties of an Argiaquoll soil under long-term crop rotation in Buenos Aires, Argentina. Int. Soil Water Conserv. Res. 2020, 8, 185–194. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, N.; Yan, J.; Zhang, S. Effects of Soft Rock and Biochar Applications on Millet (Setaria italica L.) Crop Performance in Sandy Soil. Agronomy 2020, 10, 669. [Google Scholar] [CrossRef]
- Smith, M.R.; Reis Hodecker, B.E.; Fuentes, D.; Merchant, A. Investigating Nutrient Supply Effects on Plant Growth and Seed Nutrient Content in Common Bean. Plants 2022, 11, 737. [Google Scholar] [CrossRef]
- Raven, J.A.; Smith, F.A. Nitrogen assimilation and transport in vascular land plants in relation to intracellular ph regulation. New Phytol. 1976, 76, 415–431. [Google Scholar] [CrossRef]
- Xiao, X.; Fan, M.; Wang, E.; Chen, W.; Wei, G. Interactions of plant growth-promoting rhizobacteria and soil factors in two leguminous plants. Appl. Microbiol. Biotechnol. 2017, 101, 8485–8497. [Google Scholar] [CrossRef] [PubMed]
- Daigh, A.L.M.; Dick, W.A.; Helmers, M.J.; Lal, R.; Lauer, J.G.; Nafziger, E.; Pederson, C.H.; Strock, J.; Villamil, M.; Mukherjee, A.; et al. Yields and yield stability of no-till and chisel-plow fields in the Midwestern US Corn Belt. Field Crops Res. 2018, 218, 243–253. [Google Scholar] [CrossRef]
- Zhao, C.; Gao, J.; Huang, Y.; Wang, G.; Zhang, M. Effects of Vegetation Stems on Hydraulics of Overland Flow Under Varying Water Discharges. Land Degrad. Dev. 2016, 27, 748–757. [Google Scholar] [CrossRef]
- Steponavičienė, V.; Bogužas, V.; Sinkevičienė, A.; Skinulienė, L.; Vaisvalavičius, R.; Sinkevičius, A. Soil Water Capacity, Pore Size Distribution, and CO2 Emission in Different Soil Tillage Systems and Straw Retention. Plants 2022, 11, 614. [Google Scholar] [CrossRef]
- Pucciariello, C.; Perata, P. The Oxidative Paradox in Low Oxygen Stress in Plants. Antioxidants 2021, 10, 332. [Google Scholar] [CrossRef]
- Siczek, A.; Lipiec, J. Soybean nodulation and nitrogen fixation in response to soil compaction and surface straw mulching. Soil Tillage Res. 2011, 114, 50–56. [Google Scholar] [CrossRef]
- Marcelo, A.V.; Corá, J.E.; Fernandes, C.; Martins, M.D.R.; Jorge, R.F. Crop sequences in no-tillage system: Effects on soil fertility and soybean, maize and rice yield. Rev. Bras. Ciência Do Solo 2009, 33, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Calonego, J.C.; Rosolem, C.A. Soybean root growth and yield in rotation with cover crops under chiseling and no-till. Eur. J. Agron. 2010, 33, 242–249. [Google Scholar] [CrossRef]
- Hama, J.R.; Kolpin, D.W.; LeFevre, G.H.; Hubbard, L.E.; Powers, M.M.; Strobel, B.W. Exposure and transport of alkaloids and phytoestrogens from soybeans to agricultural soils and streams in the midwestern United States. Environ. Sci. Technol. 2021, 55, 11029–11039. [Google Scholar] [CrossRef]
- Henry, C.; John, G.P.; Pan, R.; Bartlett, M.K.; Fletcher, L.R.; Scoffoni, C.; Sack, L. A stomatal safety-efficiency trade-off constrains responses to leaf dehydration. Nat. Commun. 2019, 10, 3398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- do Rosário Rosa, V.; dos Santos, A.L.F.; da Silva, A.A.; Sab, M.P.V.; Germino, G.H.; Cardoso, F.B.; de Almeida Silva, M. Increased soybean tolerance to water deficiency through biostimulant based on fulvic acids and Ascophyllum nodosum (L.) seaweed extract. Plant Physiol. Biochem. 2021, 158, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, N.A.; Moses, V.; Prakash, C. The impact of possible climate changes on developing countries: The needs for plants tolerant to abiotic stresses. GM Crops Food 2014, 5, 77–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basche, A.D.; Delonge, M.S. Comparing infiltration rates in soils managed with conventional and alternative farming methods: A meta-analysis. PLoS One 2019, 14, e0215702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Andrade Bonetti, J.; Anghinoni, I.; de Moraes, M.T.; Fink, J.R. Resilience of soils with different texture, mineralogy and organic matter under long-term conservation systems. Soil Tillage Res. 2017, 174, 104–112. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014; Volume 12.
- Raij, B.V.; Andrade, J.C.; Cantarella, H.; Quaggio, J.A. Análise Química Para Avaliação Da Fertilidade De Solos Tropicais; IAC: Campinas, Brazil, 2001. [Google Scholar]
- EMBRAPA. Manual de Métodos de Análise de Solo, 3rd ed.; Teixeira, P.C., Donagemma, G.K., Fontana, A., Teixeira, W.G.T., Eds.; Embrapa: Brasílis, Brazil, 2017. [Google Scholar]
- Escobedo, J.F.; Gomes, E.N.; Oliveira, A.P.; Soares, J. Modeling hourly and daily fractions of UV, PAR and NIR to global solar radiation under various sky conditions at Botucatu, Brazil. Appl. Energy 2009, 86, 299–309. [Google Scholar] [CrossRef]
- Watson, D.J. Comparative Physiological Studies on the Growth of Field Crops: I. Variation in Net Assimilation Rate and Leaf Area between Species and Varieties, and within and between Years. Ann. Bot. 1947, 11, 41–76. [Google Scholar] [CrossRef]
- Watson, D.J. Comparative Physiological Studies on the Growth of Field Crops: II. The Effect of Varying Nutrient Supply on Net Assimilation Rate and Leaf Area. Ann. Bot. 1947, 11, 375–407. [Google Scholar] [CrossRef]
- Jamaux, I.; Steinmetz, A.; Belhassen, E. Looking for molecular and physiological markers of osmotic adjustment in sunflower. New Phytol. 1997, 137, 117–127. [Google Scholar] [CrossRef]
- Ministério da Agricultura, Pecuária e Abastecimento. Regras Para Análise de Sementes; Ministério da Agricultura, Pecuária e Abastecimento: Brasília, DF, Brasil, 2009.
- Blake, G.R.; Hartge, K.H. Bulk Density. In Methods of Soil Analysis: Physical and Mineralogical Methods; Klute, A., Ed.; America Society of Agronomy: Madison, WI, USA, 1986; pp. 363–375. [Google Scholar]
- Smith, K.A.; Mullins, C.E. Soil Analysis: Physical Methods; Marcel Dekker: New York, NY, USA, 1991. [Google Scholar]
- Reeve, M.J.; Carter, A.D. Water release characteristic. In Soil Analysis: Physical Methods; Smith, K.A., Mullins, C.E., Eds.; Marcel Dekker: New York, NY, USA, 1991; pp. 111–160. [Google Scholar]
- Bernardo, S.; Mantovani, E.C.; Da Silva, D.D.; Soares, A.A. Manual de Irrigação, 9th ed.; UFV: Viçosa, Brazil, 2019. [Google Scholar]
- Hair, J.F., Jr.; Black, W.C.; Babin, B.J.; Anderson, R.E.; Tatham, R.L. Análise Multivariada de Dados, 6th ed.; Bookman: Porto Alegre, Brazil, 2009. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Variables | Explanation Percentage (%) |
|---|---|
| Yield components | 71.62 |
| Physiological traits | 75.17 |
| Root development | 99.99 |
| Soil physics | 98.80 |
| Soil water | 97.70 |
| Spring Management | pH | P | OM | H + Al | Ca | Mg | K | Al | Sand | Silt | Clay |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CaCl2 | mg dm−3 | mmolc dm−3 | g kg−1 | ||||||||
| Millet | 4.6 | 47.5 | 27.1 | 54.8 | 7.4 | 2.3 | 5.5 | 8.8 | 110 | 245 | 645 |
| Sunn hemp | 5.0 | 43.0 | 30.9 | 46.3 | 8.2 | 3.3 | 4.9 | 4.0 | |||
| Chiseling | 5.1 | 42.3 | 26.5 | 41.3 | 8.1 | 3.3 | 6.7 | 3.0 | |||
| Year | Fall-Winter * | Spring | Summer |
|---|---|---|---|
| From 2003 to 2018 | Triticale | Sunn Hemp | Soybean |
| Millet | |||
| Fallow/chiseling ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, G.F.d.; Matusevicius, A.P.O.; Calonego, J.C.; Chamma, L.; Luperini, B.C.O.; Alves, M.d.S.; Leite, H.M.F.; Pinto, E.d.J.; Silva, M.d.A.; Putti, F.F. Soil–Plant Relationships in Soybean Cultivated under Crop Rotation after 17 Years of No-Tillage and Occasional Chiseling. Plants 2022, 11, 2657. https://doi.org/10.3390/plants11192657
Silva GFd, Matusevicius APO, Calonego JC, Chamma L, Luperini BCO, Alves MdS, Leite HMF, Pinto EdJ, Silva MdA, Putti FF. Soil–Plant Relationships in Soybean Cultivated under Crop Rotation after 17 Years of No-Tillage and Occasional Chiseling. Plants. 2022; 11(19):2657. https://doi.org/10.3390/plants11192657
Chicago/Turabian StyleSilva, Gustavo Ferreira da, Ana Paula Oliveira Matusevicius, Juliano Carlos Calonego, Larissa Chamma, Bruno Cesar Ottoboni Luperini, Michely da Silva Alves, Hugo Mota Ferreira Leite, Elizabete de Jesus Pinto, Marcelo de Almeida Silva, and Fernando Ferrari Putti. 2022. "Soil–Plant Relationships in Soybean Cultivated under Crop Rotation after 17 Years of No-Tillage and Occasional Chiseling" Plants 11, no. 19: 2657. https://doi.org/10.3390/plants11192657
APA StyleSilva, G. F. d., Matusevicius, A. P. O., Calonego, J. C., Chamma, L., Luperini, B. C. O., Alves, M. d. S., Leite, H. M. F., Pinto, E. d. J., Silva, M. d. A., & Putti, F. F. (2022). Soil–Plant Relationships in Soybean Cultivated under Crop Rotation after 17 Years of No-Tillage and Occasional Chiseling. Plants, 11(19), 2657. https://doi.org/10.3390/plants11192657