
Seasonal Variations of Fine Root Dynamics in Rubber-Flemingia macrophylla Intercropping System in Southwestern China
 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Seasonal Dynamics of Fine Root Biomass
2.2. Total Fine Root Biomass Contribution of Different Species
2.3. Seasonal Dynamics of Available Soil Nutrients
2.3.1. Available Soil Nitrogen (N)
2.3.2. Available Soil Phosphorus (P)
2.3.3. Soil Available Potassium (K)
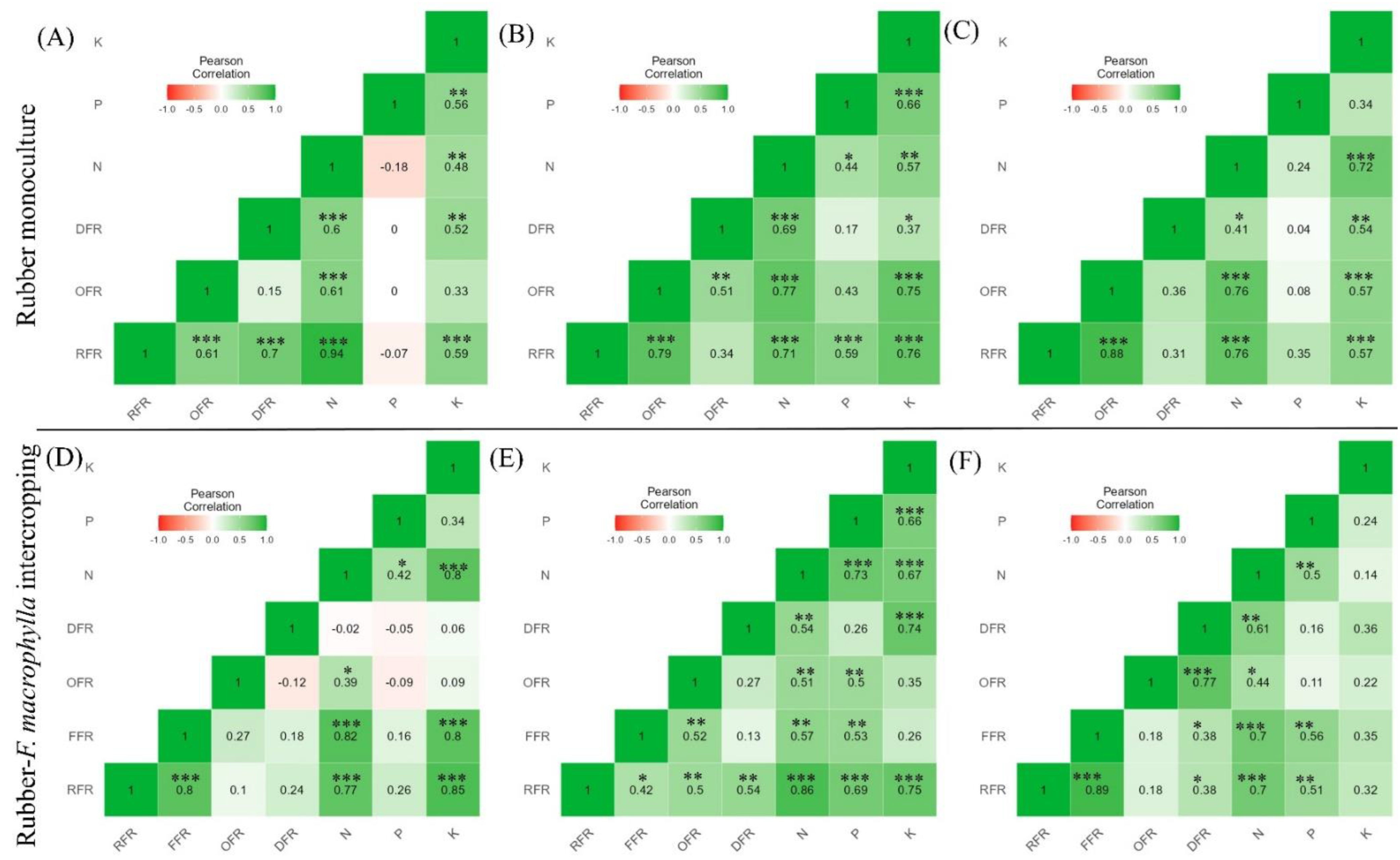
2.4. Effect of Available Soil Nutrients on Fine Root Biomass
3. Discussion
3.1. Fine Root Biomass and Nutrients Dynamics in Two Management Systems
3.2. Effect of Stand Ages
3.3. Response of Fine Roots Biomass towards Soil Available Nutrients
4. Materials and Methods
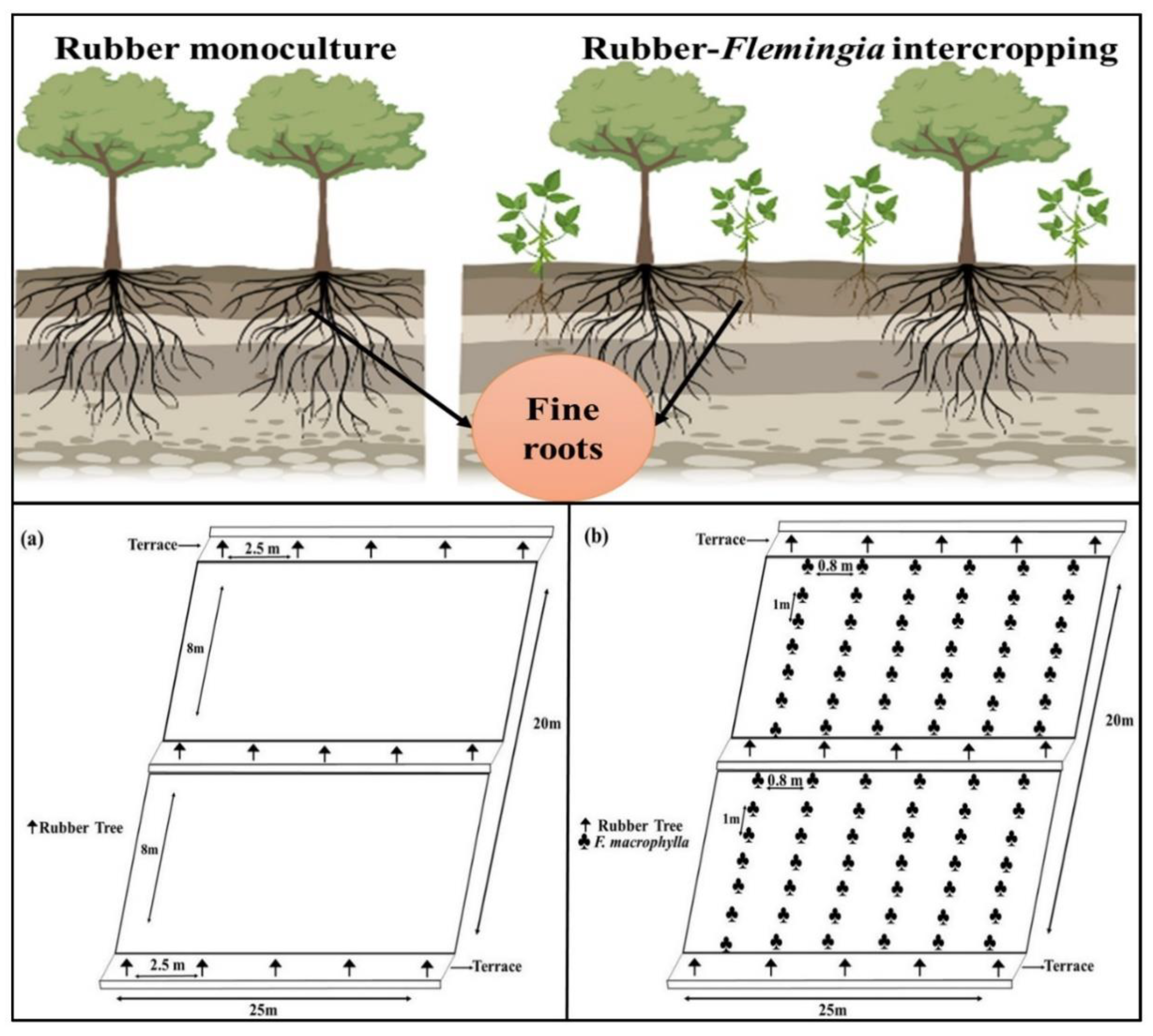
4.1. Study Site
4.2. Collection of Fine Roots Samples
4.3. Estimation of Fine Root Biomass
4.4. Soil Nutrient (C, N, P, and K) Determination
4.5. Data Processing and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, D.; Slik, J.W.; Jeon, Y.-S.; Tomlinson, K.W.; Yang, X.; Wang, J.; Kerfahi, D.; Porazinska, D.L.; Adams, J.M. Tropical forest conversion to rubber plantation affects soil micro-& mesofaunal community & diversity. Sci. Rep. 2019, 9, 5893. [Google Scholar]
- Zhou, X.; Wang, Z.; Yu, B.; Seitz, L. Effects of Large-Scale Rubber Farm on Soil Erosion and River Bed Material in the Upper Mekong Basin. In River Flow; Geology; 2014; pp. 2305–2312. Available online: https://scholar.google.co.uk/scholar?hl=nl&as_sdt=0%2C5&q=Effects+of+large-scale+rubber+farm+on+soil+erosion+and+river+bed+material+in+the+upper+Mekong+Basin&btnG= (accessed on 12 May 2022).
- Cheng, C.M.; Wang, R.S.; Jiang, J.S. Variation of soil fertility and carbon sequestration by planting Hevea brasiliensis in Hainan Island, China. J. Environ. Sci. 2007, 19, 348–352. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, M.; Zhang, Y. Vertical distribution of fine-root area in relation to stand age and environmental factors in black locust (Robinia pseudoacacia) forests of the Chinese Loess Plateau. Can. J. For. Res. 2018, 48, 1148–1158. [Google Scholar] [CrossRef]
- Yang, B.; Meng, X.; Zhu, X.; Zakari, S.; Singh, A.K.; Bibi, F.; Mei, N.; Song, L.; Liu, W. Coffee performs better than amomum as a candidate in the rubber agroforestry system: Insights from water relations. Agric. Water Manag. 2021, 244, 106593. [Google Scholar] [CrossRef]
- Raizada, A.; Jayaprakash, J.; Rathore, A.C.; Tomar, J.M.S. Distribution of fine root biomass of fruit and forest tree species raised on old river bed lands in the north west Himalaya. Trop. Ecol. 2013, 54, 251–261. [Google Scholar]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Chen, H.Y.H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. CRC. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.-D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L.; Hodge, A.; Kuzyakov, Y. Plant and mycorrhizal regulation of rhizodeposition. New Phytol. 2004, 163, 459–480. [Google Scholar] [CrossRef]
- Goebel, M.; Hobbie, S.E.; Bulaj, B.; Zadworny, M.; Archibald, D.D.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Decomposition of the finest root branching orders: Linking belowground dynamics to fine-root function and structure. Ecol. Monogr. 2011, 81, 89–102. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Jagodzinski, A.M.; Ziółkowski, J.; Warnkowska, A.; Prais, H. Tree age effects on fine root biomass and morphology over chronosequences of Fagus sylvatica, Quercus robur and Alnus glutinosa stands. PLoS ONE 2016, 11, e0148668. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, H.; Huang, Y.; Chen, G.; Ye, Y. Potential effects of sea level rise on decomposition and nutrient release of dead fine roots in a Kandelia obovata forest. Estuar. Coast. Shelf Sci. 2022, 268, 107809. [Google Scholar] [CrossRef]
- Nakayama, M.; Tateno, R. In-situ measurement of the effect of canopy tree fine roots on nitrogen availability in forest soil. Rhizosphere 2021, 18, 100324. [Google Scholar] [CrossRef]
- He, L.; Jia, Z.; Li, Q.; Zhang, Y.; Wu, R.; Dai, J.; Gao, Y. Fine root dynamic characteristics and effect on plantation’s carbon sequestration of three Salix shrub plantations in Tibetan Plateau alpine sandy land. Ecol. Evol. 2021, 11, 2645–2659. [Google Scholar] [CrossRef]
- Fox, J.; Castella, J.-C.; Ziegler, A.D.; Westley, S.B. Rubber Plantations Expand in Mountainous Southeast Asia: What Are the Consequences for the Environment? East-West Center: Honolulu, HI, USA, 2014. [Google Scholar]
- Kumar, K.A. Cost and return analysis of natural rubber plantations in Pathanamthitta District in Kerala. MUDRA J. Financ. Account. 2020, 7, 98–110. [Google Scholar]
- Raza, M.A.; Feng, L.Y.; van der Werf, W.; Iqbal, N.; Khan, I.; Khan, A.; Din, A.M.U.; Naeem, M.; Meraj, T.A.; Hassan, M.J. Optimum strip width increases dry matter, nutrient accumulation, and seed yield of intercrops under the relay intercropping system. Food Energy Secur. 2020, 9, e199. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.A.; Gul, H.; Wang, J.; Yasin, H.S.; Qin, R.; Khalid, M.H.B.; Naeem, M.; Feng, L.Y.; Iqbal, N.; Gitari, H. Land productivity and water use efficiency of maize-soybean strip intercropping systems in semi-arid areas: A case study in Punjab Province, Pakistan. J. Clean. Prod. 2021, 308, 127282. [Google Scholar] [CrossRef]
- Raza, M.A.; Feng, L.Y.; van der Werf, W.; Iqbal, N.; Khan, I.; Hassan, M.J.; Ansar, M.; Chen, Y.K.; Xi, Z.J.; Shi, J.Y. Optimum leaf defoliation: A new agronomic approach for increasing nutrient uptake and land equivalent ratio of maize soybean relay intercropping system. Field Crop. Res. 2019, 244, 107647. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, W.; Jiang, X.J.; Wang, P.; Li, W. Effects of land-use changes on runoff and sediment yield: Implications for soil conservation and forest management in Xishuangbanna, Southwest China. Land Degrad. Dev. 2018, 29, 2962–2974. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Chen, C. Below-ground interspecific competition for water in a rubber agroforestry system may enhance water utilization in plants. Sci. Rep. 2016, 6, 19502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Jin, Y.; Hu, Y.; Tang, J.; Xiong, Q.; Xu, M.; Bibi, F.; Beng, K.C. Drivers of soil bacterial community structure and diversity in tropical agroforestry systems. Agric. Ecosyst. Environ. 2019, 278, 24–34. [Google Scholar] [CrossRef]
- Lalnunzira, C.; Tripathi, S.K.; Lalnunzira, C.; Tripathi, S.K. Leaf and root production, decomposition and carbon and nitrogen fluxes during stand development in tropical moist forests, north-east India. Soil Res. 2018, 56, 306–317. [Google Scholar] [CrossRef]
- Bautista-Cruz, A.; Del Castillo, R.F. Soil changes during secondary succession in a tropical montane cloud forest area. Soil Sci. Soc. Am. J. 2005, 69, 906–914. [Google Scholar] [CrossRef]
- Powers, J.S.; Marín-Spiotta, E. Ecosystem processes and biogeochemical cycles in secondary tropical forest succession. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 497–519. [Google Scholar] [CrossRef] [Green Version]
- Eaton, J.M.; Lawrence, D. Loss of carbon sequestration potential after several decades of shifting cultivation in the Southern Yucatán. For. Ecol. Manag. 2009, 258, 949–958. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Wen, D.; Yu, K. Soil potential labile but not occluded phosphorus forms increase with forest succession. Biol. Fertil. Soils 2016, 52, 41–51. [Google Scholar] [CrossRef]
- Bibi, F.; Tomlinson, K.W.; Liu, C.; Liu, C.; Jin, Y.; Tang, J. Fine Root Production and Soil Available Nutrients in Rubber Monoculture versus Rubber–Flemingia macrophylla Agroforestry. Forests 2022, 13, 830. [Google Scholar] [CrossRef]
- Meinen, C.; Hertel, D.; Leuschner, C. Biomass and morphology of fine roots in temperate broad-leaved forests differing in tree species diversity: Is there evidence of below-ground overyielding? Oecologia 2009, 161, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Domisch, S.; Amatulli, G.; Jetz, W. Near-global freshwater-specific environmental variables for biodiversity analyses in 1 km resolution. Sci. Data 2015, 2, 150073. [Google Scholar] [CrossRef] [Green Version]
- Jacob, A.; Hertel, D.; Leuschner, C. On the significance of belowground overyielding in temperate mixed forests: Separating species identity and species diversity effects. Oikos 2013, 122, 463–473. [Google Scholar] [CrossRef]
- Cavard, X.; Bergeron, Y.; Chen, H.Y.H.; Paré, D. Mixed-species effect on tree aboveground carbon pools in the east-central boreal forests. Can. J. For. Res. 2010, 40, 37–47. [Google Scholar] [CrossRef]
- Dybzinski, R.; Fargione, J.E.; Zak, D.R.; Fornara, D.; Tilman, D. Soil fertility increases with plant species diversity in a long-term biodiversity experiment. Oecologia 2008, 158, 85–93. [Google Scholar] [CrossRef]
- Wu, J.; Zeng, H.; Zhao, F.; Chen, C.; Jiang, X.; Zhu, X.; Wang, P.; Wu, Z.; Liu, W. The nutrient status of plant roots reveals competition intensities in rubber agroforestry systems. Forests 2020, 11, 1163. [Google Scholar] [CrossRef]
- Huston, M.A.; DeAngelis, D.L. Competition and coexistence: The effects of resource transport and supply rates. Am. Nat. 1994, 144, 954–977. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Schaefer, D.A.; Chan, O.C.; Zou, X. Decomposition differences of labile carbon from litter to soil in a tropical rain forest and rubber plantation of Xishuangbanna, southwest China. Eur. J. Soil Biol. 2013, 55, 55–61. [Google Scholar] [CrossRef]
- Liu, C.; Jin, Y.; Liu, C.; Tang, J.; Wang, Q.; Xu, M. Phosphorous fractions in soils of rubber-based agroforestry systems: Influence of season, management and stand age. Sci. Total Environ. 2018, 616, 1576–1588. [Google Scholar] [CrossRef]
- Wang, F.J. The Content of Nitrogen Fixed by Flemingia macrophylla and Its Effects on the Growth of Rubber and Concentration of Soil Total Nitrogen in Rubber–Flemingia macrophylla Intercropped Systems in Different Stand Ages. Master’s Thesis, University of Chinese Academy of Sciences, Beijing, China, 2015. [Google Scholar]
- Pei, Y.; Lei, P.; Xiang, W.; Ouyang, S.; Xu, Y. Effect of stand age on fine root biomass, production and morphology in Chinese fir plantations in subtropical China. Sustainability 2018, 10, 2280. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Barrantes, O.J.; Horning, A.L.; Smemo, K.A.; Blackwood, C.B.; Smemo, K.A.; Blackwood, C.B.; Valverde-Barrantes, O.J.; Horning, A.L. Phylogenetically structured traits in root systems influence arbuscular mycorrhizal colonization in woody angiosperms. Plant Soil 2016, 404, 1–12. [Google Scholar] [CrossRef]
- Singha, L.P.; Pandey, P. Glutathione and glutathione-S-transferase activity in Jatropha curcas in association with pyrene degrader Pseudomonas aeruginosa PDB1 in rhizosphere, for alleviation of stress induced by polyaromatic hydrocarbon for effective rhizoremediation. Ecol. Eng. 2017, 102, 422–432. [Google Scholar] [CrossRef]
- Liu, B.; Li, H.; Zhu, B.; Koide, R.T.; Eissenstat, D.M.; Guo, D. Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species. New Phytol. 2015, 208, 125–136. [Google Scholar] [CrossRef]
- Singha, D.; Brearley, F.Q.; Tripathi, S.K. Fine root and soil nitrogen dynamics during stand development following shifting agriculture in Northeast India. Forests 2020, 11, 1236. [Google Scholar] [CrossRef]
- Raveendra, S.; Nissanka, S.P.; Somasundaram, D.; Atapattu, A.J.; Mensah, S. Coconut-gliricidia mixed cropping systems improve soil nutrients in dry and wet regions of Sri Lanka. Agrofor. Syst. 2021, 95, 307–319. [Google Scholar] [CrossRef]
- Helmisaari, H.-S. Nutrient cycling in Pinus sylvestris stands in eastern Finland. Plant Soil 1995, 168, 327–336. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct. Ecol. 2007, 21, 191–201. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Prieto, P.; Estiarte, M. Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant Soil 2008, 306, 261–271. [Google Scholar] [CrossRef]
- Walley, F.L.; Van Kessel, C.; Pennock, D.J. Landscape-scale variability of N mineralization in forest soils. Soil Biol. Biochem. 1996, 28, 383–391. [Google Scholar] [CrossRef]
- Assefa, D.; Rewald, B.; Sandén, H.; Godbold, D.L. Fine root dynamics in afromontane forest and adjacent land uses in the northwest ethiopian highlands. Forests 2017, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Li, B.; Zhong, Y.; Chen, J. Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences. Hydrobiology 2004, 528, 99–106. [Google Scholar] [CrossRef]
- Contador, T.; Kennedy, J.H.; Rozzi, R.; Villarroel, J.O. Sharp altitudinal gradients in Magellanic Sub-Antarctic streams: Patterns along a fluvial system in the Cape Horn Biosphere Reserve (55°S). Polar Biol. 2015, 38, 1853–1866. [Google Scholar] [CrossRef]
- Merchant, A.; Tausz, M.; Arndt, S.K.; Adams, M.A. Cyclitols and carbohydrates in leaves and roots of 13 Eucalyptus species suggest contrasting physiological responses to water deficit. Plant. Cell Environ. 2006, 29, 2017–2029. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.T.S.; Miranda, I.S.; Vasconcelos, S.S. Effects of water and nutrient availability on fine root growth in eastern Amazonian forest regrowth, Brazil. New Phytol. 2010, 187, 622–630. [Google Scholar] [CrossRef]
- Malhotra, S.K.; Maheswarappa, H.P.; Selvamani, V.; Chowdappa, P. Diagnosis and management of soil fertility constraints in coconut (Cocos nucifera): A review. Indian J. Agric. Sci. 2017, 87, 711–726. [Google Scholar]
- Zeng, H.; Wu, J.; Zhu, X.; Singh, A.K.; Chen, C.; Liu, W. Jungle rubber facilitates the restoration of degraded soil of an existing rubber plantation. J. Environ. Manag. 2021, 281, 111959. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Barrantes, O.J.; Raich, J.W.; Russell, A.E. Fine-root mass, growth and nitrogen content for six tropical tree species. Plant Soil 2007, 290, 357–370. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, J.; Gong, L. The morphological and chemical properties of fine roots respond to nitrogen addition in a temperate Schrenk’s spruce (Picea schrenkiana) forest. Sci. Rep. 2021, 11, 3839. [Google Scholar] [CrossRef]
- Ravenek, J.M.; Bessler, H.; Engels, C.; Scherer-Lorenzen, M.; Gessler, A.; Gockele, A.; de Luca, E.; Temperton, V.M.; Ebeling, A.; Roscher, C.; et al. Long-term study of root biomass in a biodiversity experiment reveals shifts in diversity effects over time. Oikos 2014, 123, 1528–1536. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Y.; Yang, J.; Lu, Z. Influence of tree functional diversity and stand environment on fine root biomass and necromass in four types of evergreen broad-leaved forests. Glob. Ecol. Conserv. 2020, 21, e00832. [Google Scholar] [CrossRef]
- Espeleta, J.F.; Clark, D.A. Multi-scale variation in fine-root biomass in a tropical rain forest: A seven-year study. Ecol. Monogr. 2007, 77, 377–404. [Google Scholar] [CrossRef] [Green Version]
- Van der Sande, M.T.; Arets, E.J.M.M.; Peña-Claros, M.; Hoosbeek, M.R.; Cáceres-Siani, Y.; van der Hout, P.; Poorter, L. Soil fertility and species traits, but not diversity, drive productivity and biomass stocks in a Guyanese tropical rainforest. Funct. Ecol. 2018, 32, 461–474. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.D.G.; Klieve, A.V. Does the complexity of the rumen microbial ecology preclude methane mitigation? Anim. Feed Sci. Technol. 2011, 166–167, 248–253. [Google Scholar] [CrossRef]
- Gower, S.T.; Vogt, K.A.; Grier, C.C. Carbon dynamics of Rocky Mountain douglas-fir: Influence of water and nutrient availability. Ecol. Monogr. 1992, 62, 43–65. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J. The potential effects of nitrogen deposition on fine-root production in forest ecosystems. New Phytol. 2000, 147, 131–139. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. A global analysis of fine root production as affected by soil nitrogen and phosphorus. Proc. R. Soc. B Biol. Sci. 2012, 279, 3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, Z.; Huang, C.; Li, M.; Bibi, F.; Zhou, S.; Nakamura, A. Ant assemblage composition explains high predation pressure on artificial caterpillars during early night. Ecol. Entomol. 2020, 45, 547–554. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Zhang, Y.-P.; Grace, J.; Sha, L.-Q.; Jin, Y.; Zhou, L.-G.; Lin, Y.-X.; Zhou, R.-W.; Gao, J.-B.; Song, Q.-H. Soil organic matter as affected by the conversion of natural tropical rainforest to monoculture rubber plantations under acric ferralsols. Catena 2020, 195, 104753. [Google Scholar] [CrossRef]
- Li, H.; Ma, Y.; Liu, W.; Liu, W. Soil changes induced by rubber and tea plantation establishment: Comparison with tropical rain forest soil in Xishuangbanna, SW China. Environ. Manag. 2012, 50, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, Q.-W.; Jin, Y.; Tang, J.; Lin, F.; Olatunji, O.A. Perennial cover crop biomass contributes to regulating soil P availability more than rhizosphere P-mobilizing capacity in rubber-based agroforestry systems. Geoderma 2021, 401, 115218. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Ere, L.; Reich, P.B.; Bergeron, Y.; Par, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Liao, Y.; McCormack, M.L.; Fan, H.; Wang, H.; Wu, J.; Tu, J.; Liu, W.; Guo, D. Relation of fine root distribution to soil C in a Cunninghamia lanceolata plantation in subtropical China. Plant Soil 2014, 381, 225–234. [Google Scholar] [CrossRef]
- Bouma, T.J.; Bryla, D.R. On the assessment of root and soil respiration for soils of different textures: Interactions with soil moisture contents and soil CO2 concentrations. Plant Soil 2000, 227, 215–221. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, W.; Shangguan, Z. Impact of long-term N additions upon coupling between soil microbial community structure and activity, and nutrient-use efficiencies. Soil Biol. Biochem. 2015, 91, 151–159. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility; CAB International: Wallingford, UK, 1989; ISBN 0851986358. [Google Scholar]
- Atapattu, A.J.; Xia, S.; Cao, M.; Zhang, W.; Mishra, S.; Yang, X. Can Dominant Canopy Species Leaf Litter Determine Soil Nutrient Heterogeneity? A Case Study in a Tropical Rainforest in Southwest China. J. Soil Sci. Plant Nutr. 2020, 20, 2479–2489. [Google Scholar] [CrossRef]
- Liu, C.-A.; Nie, Y.; Rao, X.; Tang, J.-W.; Siddique, K.H.M. The effects of introducing Flemingia macrophylla to rubber plantations on soil water content and exchangeable cations. Catena 2019, 172, 480–487. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Effects | Fine Roots Biomass (Mg·ha−1) | Available Nitrogen (mg·kg−1) | Available Phosphorus (mg·kg−1) | Available Potassium (mg·kg−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Factors | df | SSq | F Value | p Value | SSq | F Value | p Value | SSq | F Value | p Value | SSq | F Value | p Value |
| Season (S) | 4 | 2.13 | 5.03 | 0.001 | 5.43 | 70.59 | 0.001 | 17.42 | 12.94 | 0.001 | 0.62 | 5.02 | 0.001 |
| Management (M) | 1 | 0.25 | 2.39 | 1.00 | 0.00 | 0.16 | 0.695 | 24.14 | 71.72 | 0.001 | 0.93 | 30.09 | 0.001 |
| Age (A) | 2 | 0.5 | 2.38 | 1.00 | 0.98 | 25.38 | 0.001 | 114.64 | 170.27 | 0.001 | 0.46 | 7.49 | 0.01 |
| Depths (D) | 5 | 191.89 | 362.47 | 0.001 | 28.94 | 300.82 | 0.001 | 95.8 | 56.92 | 0.001 | 7.28 | 47.22 | 0.001 |
| S × D | 20 | 4.87 | 2.3 | 0.001 | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| A × D | 10 | 2.87 | 2.71 | 0.003 | -- | -- | -- | 17.16 | 5.1 | 0.001 | -- | -- | -- |
| S × M | 4 | 0.4 | 0.95 | 0.433 | 0.25 | 3.29 | 0.05 | 13.75 | 10.21 | 0.001 | 0.79 | 6.38 | 0.001 |
| S × A | 8 | 1.19 | 1.4 | 0.196 | 1.44 | 9.34 | 0.001 | 35.84 | 77.82 | 0.001 | 0.96 | 3.9 | 0.001 |
| M × A | 2 | 0.16 | 0.77 | 1.00 | 0.1 | 2.6 | 0.102 | 52.39 | 77.82 | 0.001 | 0.67 | 10.84 | 0.001 |
| M × D | 5 | 0.54 | 1.03 | 0.402 | -- | -- | -- | 1.14 | 0.67 | 0.643 | -- | -- | -- |
| S × M × A | 8 | 0.93 | 1.1 | 0.362 | 1.031 | 6.69 | 0.001 | 17.74 | 6.59 | 0.001 | 1.43 | 5.81 | 0.001 |
| R2 | -- | -- | -- | -- | |||||||||
| R2 m | -- | 0.76 | -- | -- | 0.78 | -- | -- | 0.85 | -- | -- | 0.64 | -- | -- |
| R2 c | -- | 0.82 | -- | -- | 0.79 | -- | -- | 0.87 | -- | -- | 0.77 | -- | -- |
| Fine Roots | Age | M.S | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|---|---|
| Rubber FR | 12 | Mono | 0.52 ± 0.07 a | 0.50 ± 0.09 a | 0.53 ± 0.08 a | 0.53 ± 0.10 a | 0.58 ± 0.07 a |
| Mix | 0.36 ± 0.05 ab | 0.37 ± 0.06 ab | 0.37 ± 0.05 a | 0.42 ± 0.10 a | 0.48 ± 0.10 a | ||
| 15 | Mono | 0.34 ± 0.04 ab | 0.38 ± 0.06 ab | 0.32 ± 0.05 a | 0.43 ± 0.10 a | 0.45 ± 0.08 a | |
| Mix | 0.33 ± 0.04 ab | 0.24 ± 0.03 b | 0.37 ± 0.06 a | 0.37 ± 0.06 a | 0.40 ± 0.08 a | ||
| 24 | Mono | 0.40 ± 0.06 ab | 0.47 ± 0.09 ab | 0.41 ± 0.09 a | 0.43 ± 0.10 a | 0.51 ± 0.10 a | |
| Mix | 0.30 ± 0.04 b | 0.30 ± 0.05 ab | 0.27 ± 0.06 a | 0.36 ± 0.09 a | 0.32 ± 0.07 a | ||
| F. macrophylla FR | 12 | Mix | 0.08 ± 0.01 a | 0.06 ± 0.01 ab | 0.06 ± 0.01 a | 0.06 ± 0.01 a | 0.09 ± 0.01 a |
| 15 | Mix | 0.06 ± 0.01 a | 0.12 ± 0.02 a | 0.04 ± 0.01 ab | 0.05 ± 0.01 ab | 0.06 ± 0.01 a | |
| 24 | Mix | 0.02 ± 0.01 b | 0.02 ± 0.01 b | 0.02 ± 0.01 bc | 0.02 ± 0.01 bc | 0.02 ± 0.01 ab | |
| Other FR | 12 | Mono | 0.09 ± 0.02 a | 0.04 ± 0.02 a | 0.02 ± 0.01 ab | 0.01 ± 0.00 ab | 0.02 ± 0.01 a |
| Mix | 0.04 ± 0.01 a | 0.00 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 a | ||
| 15 | Mono | 0.06 ± 0.02 a | 0.05 ± 0.02 a | 0.05 ± 0.02 a | 0.06 ± 0.03 a | 0.04 ± 0.02 a | |
| Mix | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.01 ± 0.01 a | ||
| 24 | Mono | 0.01 ± 0.01 a | 0.01 ± 0.01 a | 0.01 ± 0.01 ab | 0.01 ± 0.00 ab | 0.01 ± 0.01 a | |
| Mix | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.00 ± 0.00 a | ||
| Dead FR (Necromass) | 12 | Mono | 0.00 ± 0.00 c | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.03 ± 0.01 b | 0.01 ± 0.00 a |
| Mix | 0.01 ± 0.00 c | 0.00 ± 0.00 a | 0.02 ± 0.00 a | 0.12 ± 0.05 a | 0.04 ± 0.01 a | ||
| 15 | Mono | 0.15 ± 0.04 bc | 0.00 ± 0.00 a | 0.03 ± 0.01 a | 0.02 ± 0.01 b | 0.07 ± 0.02 a | |
| Mix | 0.10 ± 0.02 bc | 0.00 ± 0.00 a | 0.03 ± 0.01 a | 0.09 ± 0.03 b | 0.04 ± 0.01 a | ||
| 24 | Mono | 0.35 ± 0.07 a | 0.01 ± 0.01 a | 0.03 ± 0.01 a | 0.05 ± 0.02 b | 0.07 ± 0.02 a | |
| Mix | 0.18 ± 0.04 b | 0.02 ± 0.01 a | 0.01 ± 0.00 a | 0.02 ± 0.01 b | 0.06 ± 0.02 a |
| Nutrients | Age | M.S | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|---|---|
| N (mg·kg−1) | 12 | Mono | 80.4 ± 5.1 c | 69.4 ± 7.1 bc | 68.8 ± 7.3 b | 71.2 ± 5.7 c | 71.6 ± 5.9 ab |
| Mix | 82.6 ± 4.6 c | 64.0 ± 7.1 c | 77.7 ± 7.2 b | 58.6 ± 4.7 d | 78.0 ± 8.9 a | ||
| 15 | Mono | 101.7 ± 4.3 ab | 77.6 ± 7.1 ab | 76.3 ± 8.3 b | 73.9 ± 7.7 bc | 76.4 ± 8.8 a | |
| Mix | 94.4 ± 5.0 b | 85.6 ± 9.1 a | 91.5 ± 9.3 a | 84.7 ± 8.7 a | 77.0 ± 7.9 a | ||
| 24 | Mono | 101.3 ± 3.8 ab | 80.1 ± 8.1 ab | 80.6 ± 9.3 ab | 82.2 ± 9.7 ab | 72.8 ± 6.9 ab | |
| Mix | 104.5 ± 5.2 a | 81.8 ± 7.1 a | 76.4 ± 7.3 b | 76.1 ± 6.7 abc | 62.9 ± 4.9 b | ||
| P (mg·kg−1) | 12 | Mono | 38.8 ± 4.4 a | 50.1 ± 8.3 a | 49.5 ± 8.4 a | 49.7 ± 4.0 a | 41.3 ± 9.4 a |
| Mix | 2.0 ± 0.2 b | 1.4 ± 0.1 b | 9.8 ± 2.4 b | 1.3 ± 0.9 b | 2.5 ± 0.8 b | ||
| 15 | Mono | 1.3 ± 0.1 b | 0.7 ± 0.1 b | 4.5 ± 1.4 b | 1.2 ± 1.0 b | 3.4 ± 0.9 b | |
| Mix | 1.6 ± 0.1 b | 1.6 ± 0.1 b | 1.4 ± 0.6 b | 0.7 ± 0.4 b | 1.6 ± 0.7 b | ||
| 24 | Mono | 1.4 ± 0.1 b | 0.8 ± 0.1 b | 0.8 ± 0.4 b | 12.3 ± 1.2 b | 0.7 ± 0.5 b | |
| Mix | 1.4 ± 0.1 b | 0.8 ± 0.1 b | 2.7 ± 1.0 b | 0.8 ± 0.4 b | 1.0 ± 0.5 b | ||
| K (mg·kg−1) | 12 | Mono | 170.3 ± 9.6 a | 211.6 ± 9.2 a | 219.7 ± 12.6 a | 208.8 ± 14.2 a | 188.7 ± 13.9 a |
| Mix | 86.5 ± 5.8 d | 94.3 ± 8.1 bc | 94.3 ± 8.8 bc | 87.4 ± 9.2 b | 102.8 ± 9.2 c | ||
| 15 | Mono | 104.5 ± 6.6 bc | 112.8 ± 8.1 b | 124.1 ± 10.9 b | 116.1 ± 12.1 b | 136.5 ± 11.9 b | |
| Mix | 111.2 ± 9.4 bc | 88.0 ± 7.2 c | 97.7 ± 8.1 bc | 97.8 ± 9.8 b | 108.4 ± 9.9 bc | ||
| 24 | Mono | 112.8 ± 6.5 b | 93.2 ± 7.1 bc | 106.6 ± 9.6 bc | 113.7 ± 11.2 b | 110.3 ± 10.9 bc | |
| Mix | 95.5 ± 9.9 cd | 105.5 ± 8.2 bc | 87.8 ± 7.6 c | 82.0 ± 8.2 b | 104.7 ± 9.9 bc |
| Age (Years) | M.S | TC (g·kg−1) | TN (g·kg−1) | TP (g·kg−1) | TK (g·kg−1) | Soil pH |
|---|---|---|---|---|---|---|
| 12 | Mono | 9.4 ± 1.5 ab | 0.3 ± 0.0 a | 0.7 ± 0.1 b | 20.1 ± 1.6 b | 5.8 |
| 12 | Mix | 7.9 ± 1.2 b | 1.1 ± 0.1 ab | 0.3 ± 0.0 a | 17.5 ± 1.0 c | 5.9 |
| 15 | Mono | 9.8 ± 0.5 ab | 0.3 ± 0.0 a | 0.3 ± 0.0 c | 16.2 ± 0.9 c | 5.0 |
| 15 | Mix | 8.9 ± 0.5 ab | 1.2 ± 0.0 a | 0.3 ± 0.0 a | 15.2 ± 0.6 d | 5.0 |
| 24 | Mono | 10.6 ± 0.7 a | 1.3 ± 0.1 a | 0.3 ± 0.0 c | 22.3 ± 3.7 a | 5.0 |
| 24 | Mix | 10.8 ± 1.2 a | 1.2 ± 0.1 a | 0.3 ± 0.0 a | 17.3 ± 1.6 c | 5.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bibi, F.; Balasubramanian, D.; Ilyas, M.; Sher, J.; Samoon, H.A.; Bin Khalid, M.H.; Alharby, H.F.; Majrashi, A.; Alghamdi, S.A.; Hakeem, K.R.; et al. Seasonal Variations of Fine Root Dynamics in Rubber-Flemingia macrophylla Intercropping System in Southwestern China. Plants 2022, 11, 2682. https://doi.org/10.3390/plants11202682
Bibi F, Balasubramanian D, Ilyas M, Sher J, Samoon HA, Bin Khalid MH, Alharby HF, Majrashi A, Alghamdi SA, Hakeem KR, et al. Seasonal Variations of Fine Root Dynamics in Rubber-Flemingia macrophylla Intercropping System in Southwestern China. Plants. 2022; 11(20):2682. https://doi.org/10.3390/plants11202682
Chicago/Turabian StyleBibi, Farkhanda, Durairaj Balasubramanian, Muhammad Ilyas, Jan Sher, Hamz Ali Samoon, Muhammad Hayder Bin Khalid, Hesham F. Alharby, Ali Majrashi, Sameera A. Alghamdi, Khalid Rehman Hakeem, and et al. 2022. "Seasonal Variations of Fine Root Dynamics in Rubber-Flemingia macrophylla Intercropping System in Southwestern China" Plants 11, no. 20: 2682. https://doi.org/10.3390/plants11202682
APA StyleBibi, F., Balasubramanian, D., Ilyas, M., Sher, J., Samoon, H. A., Bin Khalid, M. H., Alharby, H. F., Majrashi, A., Alghamdi, S. A., Hakeem, K. R., Shah, M., & Rather, S. A. (2022). Seasonal Variations of Fine Root Dynamics in Rubber-Flemingia macrophylla Intercropping System in Southwestern China. Plants, 11(20), 2682. https://doi.org/10.3390/plants11202682