
A Model Nutrition Control System in Potato Tissue Culture and Its Influence on Plant Elemental Composition

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Calculations of MS Media N, P, K, Ca, and Mg Concentrations
2.3. Plant Growth Assessments
2.4. Plant Nutrient Composition Analyses
2.5. Statistical Analyses
3. Results and Discussion
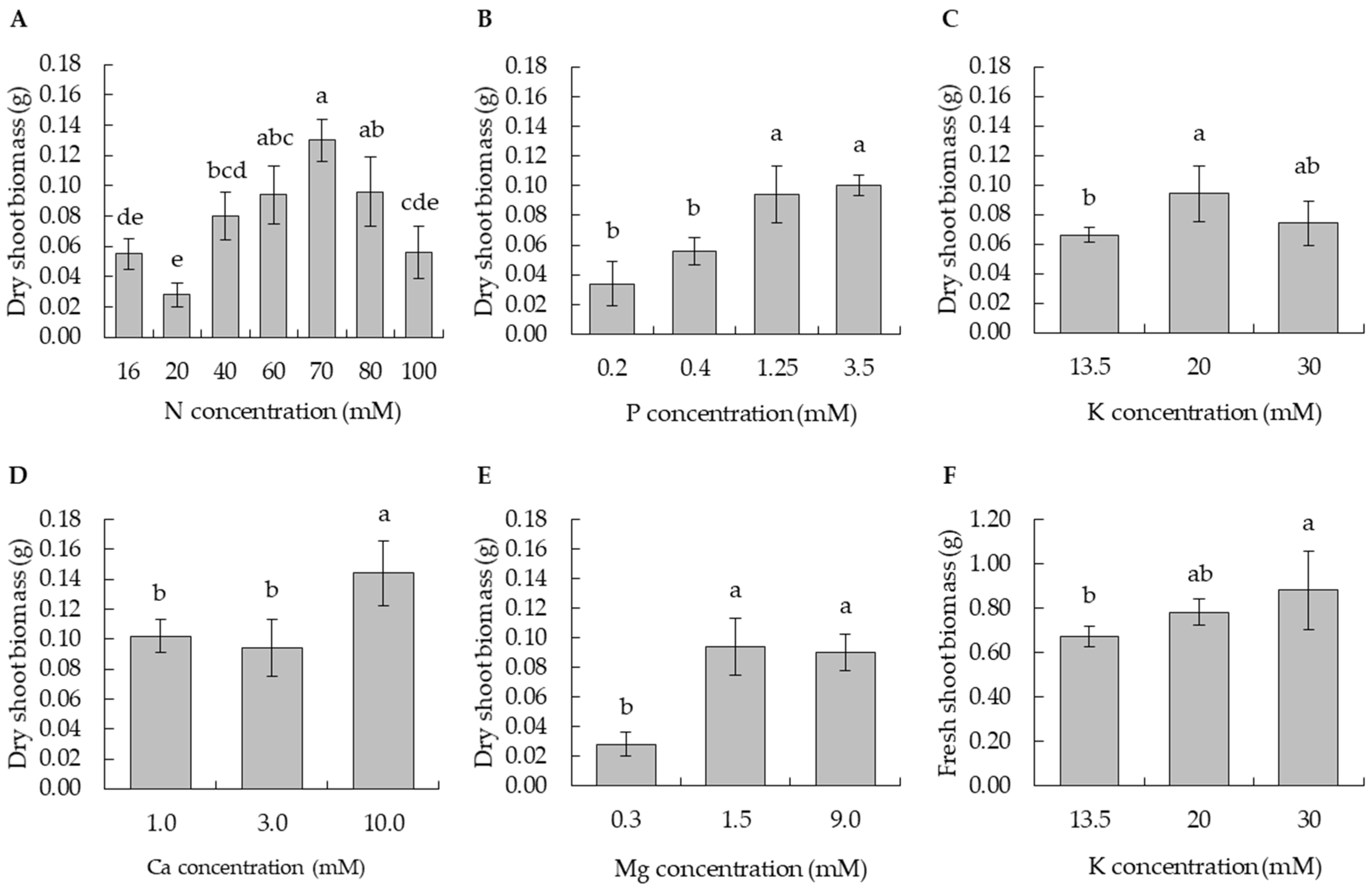
3.1. Definition of N Nutrient Status
3.2. Definition of P Nutrient Status
3.3. Definition of K Nutrient Status
3.4. Definition of Ca Nutrient Status
3.5. Definition of Mg Nutrient Status
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devaux, A.; Goffart, J.P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global food security, contributions from sustainable potato agri-food systems. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Spinger Nature: Cham, Switzerland, 2020; pp. 3–35. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, B.G.; Stark, J.C.; Kelling, K.A. Nutrient Management. In Potato Production Systems, 2nd ed.; Stark, J., Thornton, M., Nolte, P., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 155–202. [Google Scholar] [CrossRef]
- Palta, J.P. Improving potato tuber quality and production by targeted calcium nutrition: The discovery of tuber roots leading to a new concept in potato nutrition. Potato Res. 2010, 53, 267–275. [Google Scholar] [CrossRef]
- Facknath, S.; Lalljee, B. Effect of soil-applied complex fertilizers on an insect-host plant relationship: Liriomyza trifolii on Solanum tuberosum. Entomol. Exp. Appl. 2005, 115, 67–77. [Google Scholar] [CrossRef]
- Butler, J.; Garratt, M.P.D.; Leather, S.R. Fertilizers and insect herbivores: A meta-analysis. Ann. Appl. Biol. 2012, 161, 223–233. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Radouani, A.; Lauer, F.I. Effect of NPK media concentrations on in vitro potato tuberization of cultivars Nicola and Russet Burbank. Am. J. Potato Res. 2015, 92, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.T.; Alizadeh, H.; Leung, D.W.M. Response of potato (Solanum tuberosum L., cv Iwa) nodal explants to low inorganic nitrogen supply in vitro. Biocatal. Agric. Biotechnol. 2021, 38, 102215. [Google Scholar] [CrossRef]
- Upadhyaya, C.P.; Bagri, D.S.; Upadhyaya, D.C.; Pathak, K.; Kawar, P.G. Molecular and biochemical analysis of supplementation of calcium under in vitro condition on tuberization in potato (Solanum tuberosum L.). Biocatal. Agric. Biotechnol. 2016, 7, 210–216. [Google Scholar] [CrossRef]
- Walworth, J.L.; Muniz, J.E. A compendium of tissue nutrient concentrations for field grown potatoes. Am. Potato J. 1993, 70, 579–597. [Google Scholar] [CrossRef]
- Geary, B.; Clark, J.; Hopkins, B.G.; Jolley, V.D. Deficient, adequate and excess nitrogen levels established in hydroponics for biotic and abiotic stress-interaction studies in potato. J. Plant Nutr. 2015, 38, 41–50. [Google Scholar] [CrossRef]
- Bouman, H.; Tiekstra, A. Adaptations of the mineral composition of tissue culture media to the basis of plant elemental composition of hydroponic substrates. In Liquid Culture Systems for In Vitro Plant Propagation, 1st ed.; Hvoslef-Eide, A.K., Preil, W., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 493–505. [Google Scholar] [CrossRef]
- Gonçalves, S.; Correia, P.J.; Martins-Loução, M.A.; Romano, A. A new medium formulation for in vitro rooting of carob tree based on leaf macronutrients concentrations. Biol. Plant. 2005, 49, 277–280. [Google Scholar] [CrossRef]
- Terrer, C.; Tomas, F. Determination of macronutrients to be included in in vitro culture media according to leaf concentrations. J. Hortic. Sci. Biotechnol. 2001, 76, 484–488. [Google Scholar] [CrossRef]
- René, P.J.J.R.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of nutrient antagonism and synergism on yield and fertilizer use efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; kumar, S.; Mohapatra, T. Interactions between macro- and micro-nutrients in plants. Front. Plant Sci. 2021, 12, 665583. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Moreira, A.; Moraes, L.A.C.; Moraes, M.F. Root growth, nutrient uptake, and nutrient-use efficiency by roots of tropical legume cover crops as influenced by phosphorus fertilization. Commun. Soil Sci. Plant Anal. 2014, 45, 555–569. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.O. High-Temperature Oxidation: Dry Ashing. In Handbook of Reference Methods for Plant Analysis, 4th ed.; Kalra, Y.P., Ed.; CRC Press: Boca Raton, FL, USA, 1998; pp. 53–56. [Google Scholar]
- JASP Team. JASP, Version 0.15. [Computer Software]. University of Amsterdam: Amsterdam, The Netherlands, 2021. Available online: https://jasp-stats.org/download/(accessed on 13 January 2022).
- Nuzzo, R.L. An introduction to Bayesian data analysis for correlations. PM&R 2017, 9, 1278–1282. [Google Scholar]
- Schum, A.; Meise, P.; Jansen, G.; Seddig, S.; Ordon, F. Evaluation of nitrogen efficiency associated traits of starch potato cultivars under in vitro conditions. Plant Cell Tissue Organ Cult. 2017, 130, 651–665. [Google Scholar] [CrossRef]
- Cao, W.; Tibbits, T.W. Response of potatoes to nitrogen concentrations differ with nitrogen forms. J. Plant Nutr. 1998, 21, 615–623. [Google Scholar] [CrossRef]
- Roosta, H.R.; Schjoerring, J.K. Effects of ammonium toxicity on nitrogen metabolism and elemental profile of cucumber plants. J. Plant Nutr. 2007, 30, 1933–1951. [Google Scholar] [CrossRef]
- Arteta, T.A.; Hameg, R.; Landin, M.; Gallego, P.P.; Barreal, M.E. Artificial neural networks elucidated the essential role of mineral nutrients versus vitamins and plant growth regulators in achieving healthy micropropagated plants. Plants 2022, 11, 1284. [Google Scholar] [CrossRef]
- Sarkar, D.; Pandey, S.K.; Sud, K.C.; Chanemougasoundharam, A. In vitro characterization of manganese toxicity in relation to phosphorus nutrition in potato. Plant Sci. 2004, 167, 977–986. [Google Scholar] [CrossRef]
- Barben, S.A.; Hopkins, B.G.; Jolley, V.D.; Webb, B.L.; Nichols, B.A. Optimizing phosphorus and zinc concentrations in hydroponic chelator buffered nutrient solution for Russet Burbank potato. J. Plant Nutr. 2010, 33, 557–570. [Google Scholar] [CrossRef]
- Fernandes, A.M.; Soratto, R.P. Nutrition, dry matter accumulation and partitioning and phosphorus use efficiency of potato grown at different phosphorus levels in nutrient solution. Rev. Bras. Cienc. Solo 2012, 36, 1528–1537. [Google Scholar] [CrossRef] [Green Version]
- Kavka, M.; Korn, K.; Hazarika, M.; Bachmann-Pfabe, S.; Uptmoor, R. Potato root and leaf phosphatase activity in response to P deprivation. J. Plant Nutr. Soil Sci. 2021, 184, 668–677. [Google Scholar] [CrossRef]
- Koch, M.; Winkelmann, M.K.; Hasler, M.; Pawelzik, E.; Naumann, M. Root growth in light of changing magnesium distribution and transport between source and sink tissues in potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 8796. [Google Scholar] [CrossRef]
- Rosen, C.J.; Bierman, P.M. Potato yield and tuber set as affected by phosphorus fertilization. Am. J. Potato Res. 2008, 85, 110–120. [Google Scholar] [CrossRef]
- Broadley, M.R.; Bowen, H.C.; Cotterill, H.L.; Hammond, J.P.; Meacham, M.C.; Mead, A.; White, P.J. Phylogenetic variation in the shoot mineral concentration of angiosperms. J. Exp. Bot. 2004, 55, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Abd El Lateef, E.M.; Wali, A.M.; Abd El-Salam, M.S. Synergistic effect of P and K interaction on yield and yield components of mungbean (Vigna radiata (L.) Wilczek) varieties. Bull. Natl. Res. Cent. 2021, 45, 161. [Google Scholar] [CrossRef]
- Koch, M.; Busse, M.; Naumann, M.; Jakli, B.; Smit, I.; Cakmak, I.; Hermans, C.; Pawelzik, E. Differential effects of varied potassium and magnesium nutrition on production and partitioning of photoassimilates in potato plants. Physiol. Plant. 2019, 166, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [Green Version]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Allison, M.F.; Fowler, J.H.; Allen, E.J. Responses of potato (Solanum tuberosum) to potassium fertilizers. J. Agric. Sci. 2001, 136, 407–426. [Google Scholar] [CrossRef] [Green Version]
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of potassium fertilization on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374. [Google Scholar] [CrossRef]
- Xu, X.; Du, X.; Wang, F.; Sha, J.; Chen, Q.; Tian, G.; Zhu, Z.; Ge, S.; Jiang, Y. Effects of potassium levels on plant growth, accumulation and distribution of carbon, and nitrogen metabolism in apple dwarf rootstock seedlings. Front. Plant Sci. 2020, 11, 904. [Google Scholar] [CrossRef]
- ten Hoopen, F.; Cuin, T.A.; Pedas, P.; Hegelund, J.N.; Shabala, S.; Schjoerring, J.K.; Jahn, T.P. Competition between uptake of ammonium and potassium in barley and Arabidopsis roots: Molecular mechanisms and physiological consequences. J. Exp. Bot. 2010, 61, 2303–2315. [Google Scholar] [CrossRef] [Green Version]
- Armengaud, P.; Sulpice, R.; Miller, A.J.; Stitt, M.; Amtmann, A.; Gibon, Y. Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol. 2009, 150, 772–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozgen, S.; Busse, J.S.; Palta, J.P. Influence of root zone calcium on shoot tip necrosis and apical dominance of potato shoot: Simulation of this disorder by ethylene glycol tetra acetic acid and prevention by strontium. HortScience 2011, 46, 1358–1362. [Google Scholar] [CrossRef] [Green Version]
- Busse, J.S.; Ozgen, S.; Palta, J.P. Influence of root zone calcium on subapical necrosis in potato shoot cultures: Localization of injury at the tissue and cellular levels. J. Am. Soc. Hortic. Sci. 2008, 133, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wiren, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef]
- Peng, W.T.; Qi, W.L.; Nie, M.M.; Xiao, Y.B.; Liao, H.; Chen, Z.C. Magnesium supports uptake through regulating NRT2.1/2.2 in soybean. Plant Soil 2020, 457, 97–111. [Google Scholar] [CrossRef]
- Santner, J.; Smolders, E.; Wenzel, W.W.; Degryse, F. First observation of diffusion-limited plant root phosphorus uptake from nutrient solution. Plant Cell Environ. 2012, 35, 1558–1566. [Google Scholar] [CrossRef]
- Liebersbach, H.; Steingrobe, B.; Claassen, N. Roots regulate ion transport in rhizosphere to counteract reduced mobility in dry soil. Plant Soil 2004, 260, 79–88. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Concentration | NH4NO3 | KNO3 | KH2PO4 | KCl | Ca(NO3)2 4H2O | CaCl2 2H2O | MgCl2 6H2O | MgSO4 7H2O | KI | |
|---|---|---|---|---|---|---|---|---|---|---|
| (mM) | (g L−1) | |||||||||
| Standard a | 1.65 | 1.90 | 0.17 | 0.440 | 0.37 | 0.00083 | ||||
| N | 20 | 0.775 | 0.17 | 1.39 | 0.440 | 0.37 | 0.00083 | |||
| 70 | 1.925 | 1.90 | 0.17 | 0.440 | 0.37 | 0.00083 | ||||
| P | 0.4 | 1.65 | 1.90 | 0.058 | 0.062 | 0.440 | 0.37 | 0.00083 | ||
| 3.5 | 1.74 | 1.67 | 0.479 | 0.440 | 0.37 | 0.00083 | ||||
| K | 13.5 | 1.65 | 0.17 | 0.921 | 0.0312 | 0.421 | 0.37 | 0.00083 | ||
| 30 | 1.65 | 1.90 | 0.17 | 0.752 | 0.440 | 0.37 | 0.00083 | |||
| Ca | 1.0 | 1.65 | 1.90 | 0.17 | 0.147 | 0.37 | 0.00083 | |||
| 10.0 | 1.65 | 1.90 | 0.17 | 1.467 | 0.37 | 0.00083 | ||||
| Mg | 1.5 | 1.65 | 1.90 | 0.17 | 0.440 | 0.37 | 0.00083 | |||
| 9.0 | 1.65 | 1.90 | 0.17 | 0.440 | 1.52 | 0.37 | 0.00083 | |||
| Medium Element Concentration | N | P | K | Ca | Mg | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Element | (mM) | (mg Plantlet−1) | ||||||||||||||
| N | 20 | 1.90 | ± | 0.40 c | 0.26 | ± | 0.06 b | 0.73 | ± | 0.17 c | 0.26 | ± | 0.04 c | 0.10 | ± | 0.02 b |
| 40 | 4.36 | ± | 0.71 b | 0.51 | ± | 0.13 a | 3.61 | ± | 0.39 a | 0.44 | ± | 0.06 b | 0.19 | ± | 0.03 a | |
| 60 a | 6.03 | ± | 1.15 ab | 0.59 | ± | 0.13 a | 3.96 | ± | 1.40 a | 0.43 | ± | 0.07 b | 0.19 | ± | 0.06 a | |
| 70 | 7.39 | ± | 0.62 a | 0.45 | ± | 0.03 ab | 3.02 | ± | 0.28 ab | 0.58 | ± | 0.07 a | 0.20 | ± | 0.02 a | |
| 80 | 7.50 | ± | 1.89 a | 0.66 | ± | 0.12 a | 2.53 | ± | 1.08 ab | 0.30 | ± | 0.07 c | 0.15 | ± | 0.05 ab | |
| 100 | 4.98 | ± | 1.43 b | 0.44 | ± | 0.15 ab | 1.77 | ± | 0.38 bc | 0.21 | ± | 0.06 c | 0.10 | ± | 0.03 b | |
| P | 0.2 | 2.22 | ± | 0.85 c | 0.14 | ± | 0.04 c | 1.14 | ± | 0.52 b | 0.14 | ± | 0.07 c | 0.06 | ± | 0.02 c |
| 0.4 | 3.75 | ± | 0.44 b | 0.24 | ± | 0.05 c | 1.83 | ± | 0.29 b | 0.24 | ± | 0.04 c | 0.10 | ± | 0.01 c | |
| 1.25 a | 6.03 | ± | 1.15 a | 0.59 | ± | 0.13 b | 3.96 | ± | 1.40 a | 0.43 | ± | 0.07 b | 0.19 | ± | 0.06 b | |
| 3.5 | 7.36 | ± | 0.59 a | 1.35 | ± | 0.17 a | 4.71 | ± | 0.42 a | 0.62 | ± | 0.05 a | 0.25 | ± | 0.04 a | |
| K | 13.5 | 4.18 | ± | 0.46 b | 0.48 | ± | 0.14 a | 2.53 | ± | 0.73 a | 0.34 | ± | 0.07 a | 0.15 | ± | 0.02 a |
| 20.0 a | 6.03 | ± | 1.15 a | 0.59 | ± | 0.13 a | 3.96 | ± | 1.40 a | 0.43 | ± | 0.07 a | 0.19 | ± | 0.06 a | |
| 30.0 | 4.33 | ± | 0.89 b | 0.57 | ± | 0.08 a | 3.94 | ± | 0.91 a | 0.32 | ± | 0.08 a | 0.15 | ± | 0.03 a | |
| Ca | 1.0 | 6.09 | ± | 0.57 b | 0.52 | ± | 0.03 a | 2.07 | ± | 0.42 b | 0.20 | ± | 0.02 b | 0.15 | ± | 0.01 b |
| 3.0 a | 6.03 | ± | 1.15 b | 0.59 | ± | 0.13 a | 3.96 | ± | 1.40 a | 0.43 | ± | 0.07 b | 0.19 | ± | 0.06 ab | |
| 10.0 | 8.61 | ± | 1.17 a | 0.63 | ± | 0.07 a | 4.57 | ± | 0.49 a | 1.56 | ± | 0.29 a | 0.25 | ± | 0.04 a | |
| Mg | 0.3 | 1.60 | ± | 0.30 c | 0.33 | ± | 0.14 b | 0.40 | ± | 0.15 b | 0.28 | ± | 0.09 b | 0.06 | ± | 0.02 c |
| 1.5 a | 6.03 | ± | 1.15 b | 0.59 | ± | 0.13 a | 3.96 | ± | 1.40 a | 0.43 | ± | 0.07 a | 0.19 | ± | 0.06 b | |
| 9.0 | 8.04 | ± | 1.22 a | 0.59 | ± | 0.07 a | 3.50 | ± | 0.14 a | 0.33 | ± | 0.04 ab | 0.81 | ± | 0.11 a | |
| Medium Element Concentration | N | P | K | Ca | Mg | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Element | (mM) | (g kg−1) | ||||||||||||||
| N | 20 | 64.4 | ± | 3.4 c | 9.8 | ± | 2.6 a | 27.7 | ± | 8.2 b | 9.7 | ± | 1.9 a | 3.7 | ± | 0.8 a |
| 40 | 55.0 | ± | 5.3 d | 6.6 | ± | 1.7 b | 46.0 | ± | 5.9 a | 5.5 | ± | 0.8 b | 2.3 | ± | 0.2 b | |
| 60 a | 64.4 | ± | 3.4 c | 7.0 | ± | 0.9 ab | 45.6 | ± | 4.8 a | 4.3 | ± | 0.5 bc | 2.1 | ± | 0.1 bc | |
| 70 | 57.0 | ± | 3.5 cd | 3.5 | ± | 0.4 c | 23.4 | ± | 3.2 b | 4.4 | ± | 0.6 bc | 1.5 | ± | 0.2 c | |
| 80 | 78.0 | ± | 4.1 b | 6.9 | ± | 0.9 b | 25.6 | ± | 4.4 b | 3.1 | ± | 0.3 c | 1.5 | ± | 0.1 c | |
| 100 | 89.2 | ± | 3.4 a | 8.0 | ± | 1.3 ab | 32.5 | ± | 5.3 b | 3.6 | ± | 0.1 c | 1.7 | ± | 0.1 bc | |
| P | 0.2 | 66.6 | ± | 5.8 ab | 4.3 | ± | 1.4 c | 33.3 | ± | 2.8 b | 4.0 | ± | 0.3 b | 1.8 | ± | 0.2 bc |
| 0.4 | 67.4 | ± | 4.7 ab | 4.4 | ± | 1.2 c | 32.7 | ± | 2.8 b | 4.2 | ± | 0.3 b | 1.7 | ± | 0.2 c | |
| 1.25 a | 64.4 | ± | 3.4 b | 7.0 | ± | 0.9 b | 45.6 | ± | 4.8 a | 4.3 | ± | 0.5 b | 2.1 | ± | 0.1 b | |
| 3.5 | 73.6 | ± | 2.5 a | 13.4 | ± | 1.2 a | 47.4 | ± | 6.3 a | 6.3 | ± | 0.5 a | 2.5 | ± | 0.3 a | |
| K | 13.5 | 63.2 | ± | 3.0 ab | 7.3 | ± | 1.7 a | 38.0 | ± | 8.9 b | 5.2 | ± | 0.8 a | 2.3 | ± | 0.2 a |
| 20.0 a | 64.4 | ± | 3.4 a | 7.0 | ± | 0.9 a | 45.6 | ± | 4.8 ab | 4.3 | ± | 0.5 a | 2.1 | ± | 0.1 a | |
| 30.0 | 58.6 | ± | 2.6 b | 7.9 | ± | 1.2 a | 53.7 | ± | 8.6 a | 4.4 | ± | 0.3 a | 2.1 | ± | 0.1 a | |
| Ca | 1.0 | 59.8 | ± | 2.6 a | 5.2 | ± | 0.5 b | 20.2 | ± | 3.2 c | 1.9 | ± | 0.1 c | 1.5 | ± | 0.1 c |
| 3.0 a | 64.4 | ± | 3.4 a | 7.0 | ± | 0.9 a | 45.6 | ± | 4.8 a | 4.3 | ± | 0.5 b | 2.1 | ± | 0.1 a | |
| 10.0 | 60.0 | ± | 2.5 a | 4.4 | ± | 0.6 b | 32.2 | ± | 5.1 b | 10.8 | ± | 1.0 a | 1.7 | ± | 0.2 b | |
| Mg | 0.3 | 47.2 | ± | 6.4 b | 11.7 | ± | 2.0 a | 14.2 | ± | 3.2 b | 9.8 | ± | 0.7 a | 2.1 | ± | 0.2 b |
| 1.5 a | 64.4 | ± | 3.4 a | 7.0 | ± | 0.9 b | 45.6 | ± | 4.8 a | 4.3 | ± | 0.5 b | 2.1 | ± | 0.1 b | |
| 9.0 | 66.6 | ± | 6.1 a | 6.6 | ± | 0.6 b | 39.4 | ± | 4.4 a | 3.6 | ± | 0.3 b | 9.0 | ± | 0.7 a | |
| Change in Shoot Nutrient Uptake | Shoot Nutrient Uptake | n | Pearson’s r | p Value | BF10 | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|---|---|---|
| N | P | 30 | 0.723 | <0.001 ††† | 7231.98 *** | 0.461 | 0.850 |
| K | 30 | 0.554 | 0.001 †† | 56.03 ** | 0.225 | 0.747 | |
| Ca | 30 | 0.520 | 0.003 †† | 28.12 * | 0.183 | 0.724 | |
| Mg | 30 | 0.541 | 0.002 †† | 42.89 ** | 0.209 | 0.738 | |
| P | N | 20 | 0.870 | <0.001 ††† | 54,778.27 *** | 0.647 | 0.944 |
| K | 20 | 0.825 | <0.001 ††† | 5984.41 *** | 0.552 | 0.922 | |
| Ca | 20 | 0.935 | <0.001 ††† | 11,040,000.00 *** | 0.805 | 0.974 | |
| Mg | 20 | 0.908 | <0.001 ††† | 727,016.73 *** | 0.735 | 0.961 | |
| K | N | 15 | 0.382 | 0.160 | 1.43 | 0.031 | 0.713 |
| P | 15 | 0.856 | <0.001 ††† | 1152.63 *** | 0.537 | 0.947 | |
| Ca | 15 | 0.384 | 0.157 | 1.45 | 0.032 | 0.715 | |
| Mg | 15 | 0.363 | 0.183 | 1.29 | 0.029 | 0.704 | |
| Ca | N | 15 | 0.881 | <0.001 ††† | 3138.88 *** | 0.599 | 0.957 |
| P | 15 | 0.399 | 0.070 | 1.58 | 0.034 | 0.722 | |
| K | 15 | 0.629 | 0.006 †† | 11.24 * | 0.152 | 0.839 | |
| Mg | 15 | 0.743 | <0.001 ††† | 62.26 ** | 0.310 | 0.895 | |
| Mg | N | 15 | 0.812 | <0.001 ††† | 291.83 *** | 0.440 | 0.927 |
| P | 15 | 0.533 | 0.020 † | 4.23 | 0.079 | 0.791 | |
| K | 15 | 0.523 | 0.023 † | 3.87 | 0.073 | 0.786 | |
| Ca | 15 | 0.060 | 0.415 | 0.38 | 0.009 | 0.554 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munthali, C.; Kinoshita, R.; Onishi, K.; Rakotondrafara, A.; Mikami, K.; Koike, M.; Tani, M.; Palta, J.; Aiuchi, D. A Model Nutrition Control System in Potato Tissue Culture and Its Influence on Plant Elemental Composition. Plants 2022, 11, 2718. https://doi.org/10.3390/plants11202718
Munthali C, Kinoshita R, Onishi K, Rakotondrafara A, Mikami K, Koike M, Tani M, Palta J, Aiuchi D. A Model Nutrition Control System in Potato Tissue Culture and Its Influence on Plant Elemental Composition. Plants. 2022; 11(20):2718. https://doi.org/10.3390/plants11202718
Chicago/Turabian StyleMunthali, Chandiona, Rintaro Kinoshita, Kazumitsu Onishi, Aurelie Rakotondrafara, Kakeru Mikami, Masanori Koike, Masayuki Tani, Jiwan Palta, and Daigo Aiuchi. 2022. "A Model Nutrition Control System in Potato Tissue Culture and Its Influence on Plant Elemental Composition" Plants 11, no. 20: 2718. https://doi.org/10.3390/plants11202718
APA StyleMunthali, C., Kinoshita, R., Onishi, K., Rakotondrafara, A., Mikami, K., Koike, M., Tani, M., Palta, J., & Aiuchi, D. (2022). A Model Nutrition Control System in Potato Tissue Culture and Its Influence on Plant Elemental Composition. Plants, 11(20), 2718. https://doi.org/10.3390/plants11202718