
Zinc Oxide Nanoparticles: Physiological and Biochemical Responses in Barley (Hordeum vulgare L.)

 ,
,  ,
,  ,
,  ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
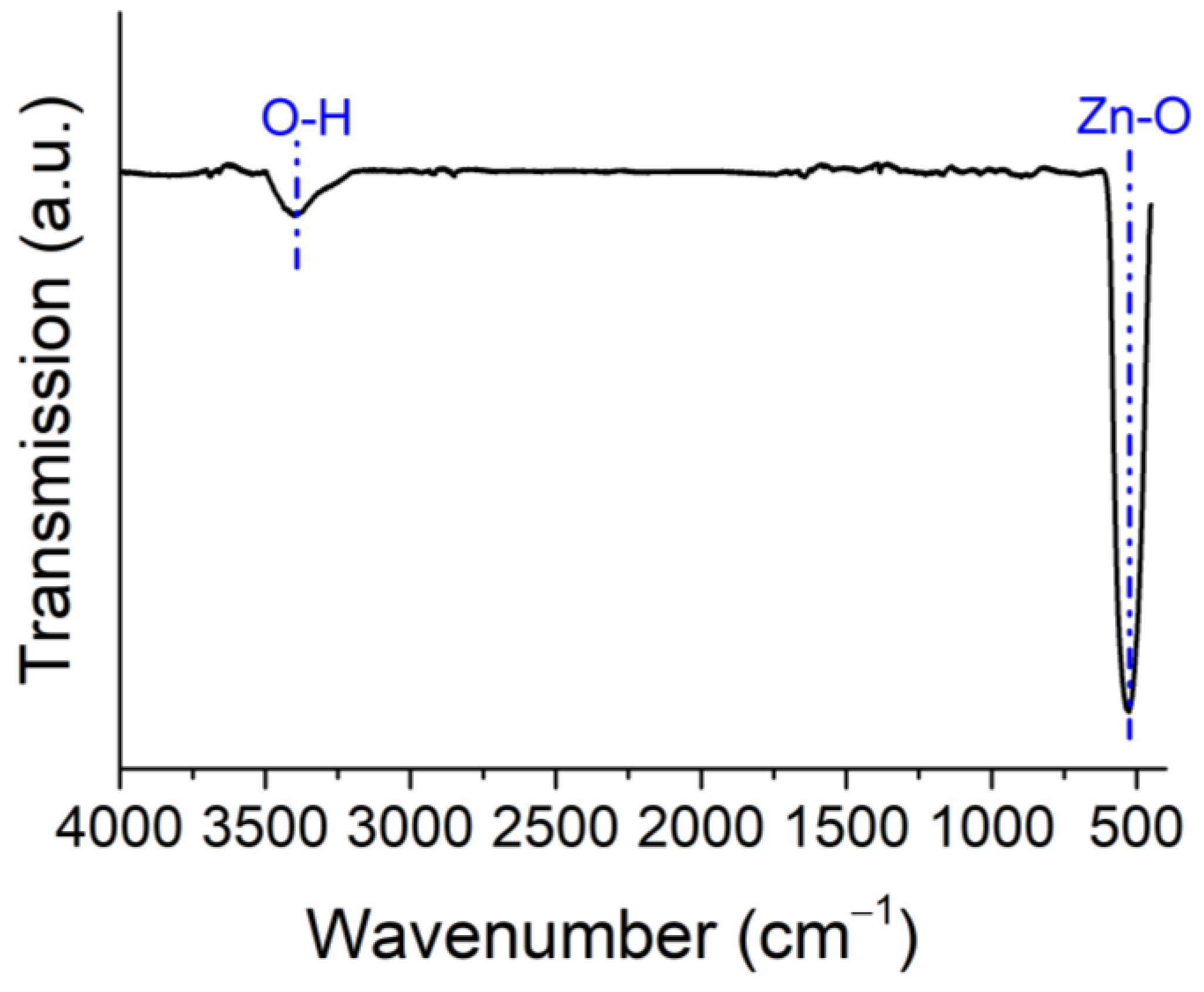
2.1. Structural and Morphology Characterization of ZnO Nanoparticles
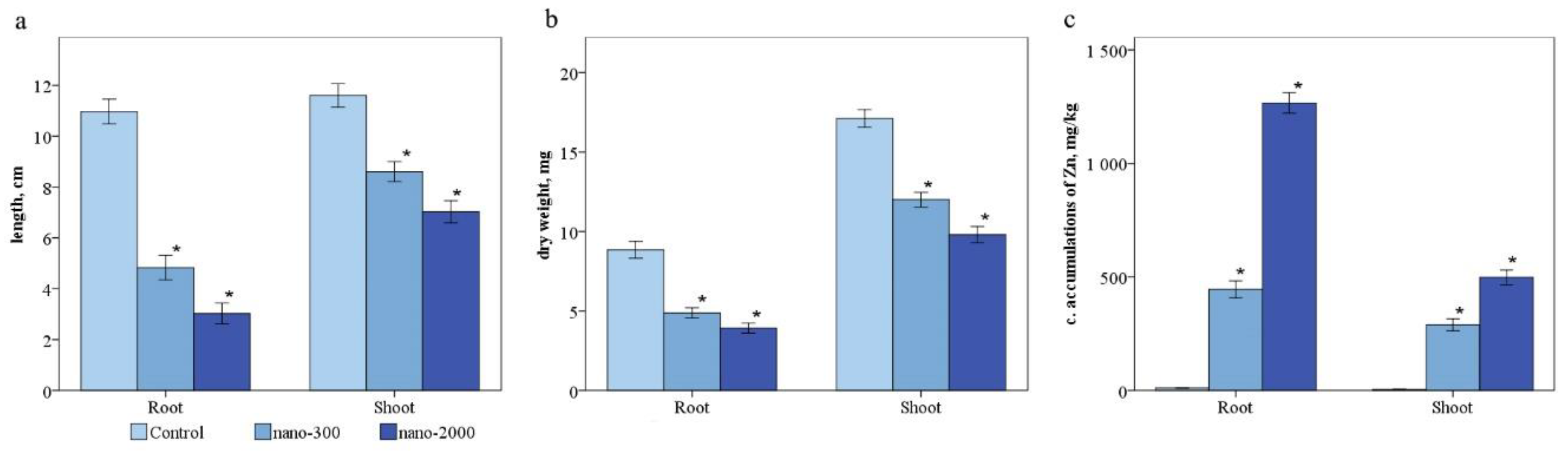
2.2. Morphobiometric Parameters and Accumulation of Zn in Plant Tissues
2.3. Effects of ZnO NPs on Biochemical Indices
2.3.1. Malonic Dialdehyde
2.3.2. Protein
2.3.3. δ-Aminolevulinic Acid
2.3.4. Proline
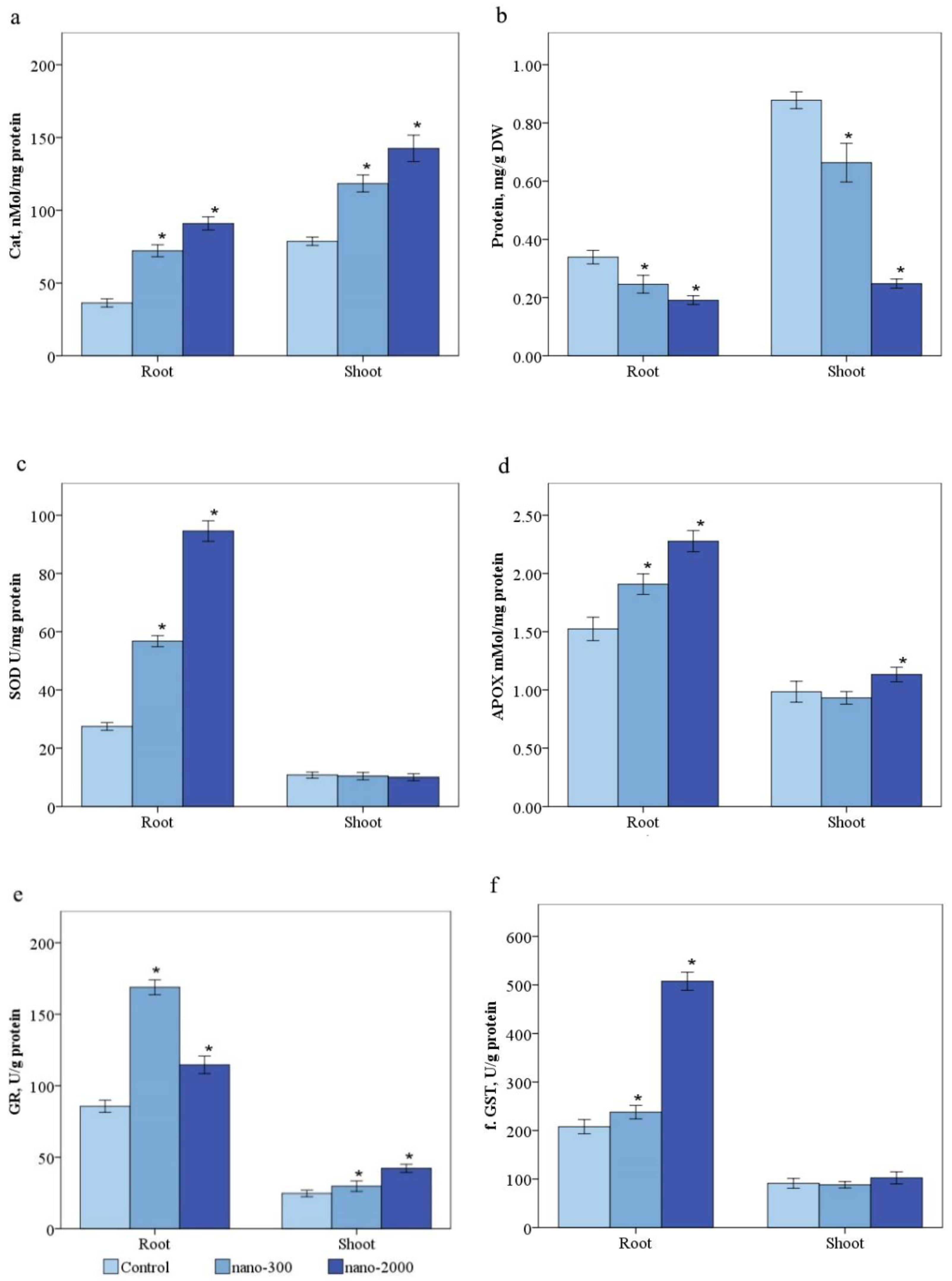
2.3.5. Catalase
2.3.6. Ascorbate Peroxidase
2.3.7. Superoxide Dismutase
2.3.8. Glutathione and Glutathione Reductase
2.3.9. Glutathione-s-Transferase
3. Discussion
4. Materials and Methods
4.1. Preparation, Characterization, and Application of Nanoparticles
4.2. Plant Growth Condition and Sample Collection for Morpho-Physiological and Biochemical Indices
4.3. Determination of Zn in Barley
4.4. Morphobiometric Analysis
4.5. Biochemical Indicators
4.5.1. Malonic Dialdehyde
4.5.2. Proline
4.5.3. δ-Aminolevulinic Acid
4.5.4. Protein
4.5.5. Superoxide Dismutase (EC 1.15.1.1)
4.5.6. Catalase (EC 1.11.1.6)
4.5.7. Glutathione Reductase (EC 1.8.1.7)
4.5.8. Ascorbate Peroxidase (EC 1.11.1.11)
4.5.9. Glutathione-S-Transferase (EC2.5.1.18)
4.5.10. Glutathione Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajput, V.D.; Minkina, T.; Upadhyay, S.K.; Kumari, A.; Ranjan, A.; Mandzhieva, S.; Sushkova, S.; Singh, R.K.; Verma, K.K. Nanotechnology in the Restoration of Polluted Soil. Nanomaterials 2022, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.; Singh, M.; Rajput, V.; Verma, C.; Burachevskaya, M. Nanofertilizers: A sustainable alternative to conventional means. In Nanotechnology for Sustainable Agriculture an Innovative and Eco-Friendly Approach; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Baig, N.; Kammakakam, I.; Falath, W. Nanomaterials: A review of synthesis methods, properties, recent progress, and challenges. Mater. Adv. 2021, 2, 1821–1871. [Google Scholar] [CrossRef]
- Singh, S.; Pradhan, A.; Singh, R.; Singh, A.; Minkina, T.; Rajput, V. Nanoparticles: Physiology, Chemistry and Biochemistry. In The Role of Nanoparticles in Plant Nutrition under Soil Pollution; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Galbraith, D.W. Nanobiotechnology: Silica breaks through in plants. Nat. Nanotechnol. 2007, 2, 272–273. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A.; et al. Effects of zinc-oxide nanoparticles on soil, plants, animals and soil organisms: A review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84. [Google Scholar] [CrossRef]
- Rico, C.M.; Barrios, A.C.; Tan, W.; Rubenecia, R.; Lee, S.C.; Varela-Ramirez, A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Physiological and biochemical response of soil-grown barley (Hordeum vulgare L.) to cerium oxide nanoparticles. Environ. Sci. Pollut. Res. 2015, 22, 10551–10558. [Google Scholar] [CrossRef] [PubMed]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of nanoparticle-induced oxidative stress and toxicity. BioMed Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmielowska-Bąk, J.; Izbiańska, K.; Deckert, J. Products of lipid, protein and RNA oxidation as signals and regulators of gene expression in plants. Front. Plant Sci. 2015, 6, 405. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Rajput, V.D.; Minkina, T.; Kumari, A.; Singh, V.K.; Verma, K.K.; Mandzhieva, S.; Sushkova, S.; Srivastava, S.; Keswani, C. Coping with the challenges of abiotic stress in plants: New dimensions in the field application of nanoparticles. Plants 2021, 10, 1221. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Öztürk, L.; Demir, Y. In vivo and in vitro protective role of proline. Plant Growth Regul. 2002, 38, 259–264. [Google Scholar] [CrossRef]
- Gumpu, M.B.; Sethuraman, S.; Krishnan, U.M.; Rayappan, J.B.B. A review on detection of heavy metal ions in water—An electrochemical approach. Sens. Actuators B Chem. 2015, 213, 515–533. [Google Scholar] [CrossRef]
- Korkmaz, A.; Korkmaz, Y.; Demirkıran, A.R. Enhancing chilling stress tolerance of pepper seedlings by exogenous application of 5-aminolevulinic acid. Environ. Exp. Bot. 2010, 67, 495–501. [Google Scholar] [CrossRef]
- Plaksenkova, I.; Kokina, I.; Petrova, A.; Jermaļonoka, M.; Gerbreders, V.; Krasovska, M. The Impact of Zinc Oxide Nanoparticles on Cytotoxicity, Genotoxicity, and miRNA Expression in Barley (Hordeum vulgare L.) Seedlings. Sci. World J. 2020, 2020, 6649746. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azarin, K.; Usatov, A.; Minkina, T.; Plotnikov, A.; Kasyanova, A.; Fedorenko, A.; Duplii, N.; Vechkanov, E.; Rajput, V.D.; Mandzhieva, S.; et al. Effects of ZnO nanoparticles and its bulk form on growth, antioxidant defense system and expression of oxidative stress related genes in Hordeum vulgare L. Chemosphere 2022, 287, 132167. [Google Scholar] [CrossRef]
- Faizan, M.; Sehar, S.; Rajput, V.D.; Faraz, A.; Afzal, S.; Minkina, T.; Sushkova, S.; Adil, M.F.; Yu, F.; Alatar, A.A.; et al. Modulation of cellular redox status and antioxidant defense system after synergistic application of Zinc oxide nanoparticles and salicylic acid in rice (Oryza sativa) plant under arsenic stress. Plants 2021, 10, 2254. [Google Scholar] [CrossRef] [PubMed]
- Zoufan, P.; Baroonian, M.; Zargar, B. ZnO nanoparticles-induced oxidative stress in Chenopodium murale L, Zn uptake, and accumulation under hydroponic culture. Environ. Sci. Pollut. Res. 2020, 27, 11066–11078. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.S.; Dietz, K.J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Kolesnikov, S.; Timoshenko, A.; Minnikova, T.; Minkina, T.; Rajput, V.D.; Kazeev, K.; Feizi, M.; Fedorenko, E.; Mandzhieva, S.; Sushkova, S. Ecotoxicological assessment of Zn, Cu and Ni based NPs contamination in Arenosols. SAINS TANAH-J. Soil Sci. Agroclimatol. 2021, 18, 9. [Google Scholar] [CrossRef]
- Fedorenko, A.G.; Minkina, T.M.; Chernikova, N.P.; Fedorenko, G.M.; Mandzhieva, S.S.; Rajput, V.D.; Burachevskaya, M.V.; Chaplygin, V.A.; Bauer, T.V.; Sushkova, S.N.; et al. The toxic effect of CuO of different dispersion degrees on the structure and ultrastructure of spring barley cells (Hordeum sativum distichum). Environ. Geochem. Health 2020, 43, 1673–1687. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Khan, M.S.A.; Borah, K.K.; Goswami, Y.; Hakeem, K.R.; Chakrabartty, I. The potential exposure and hazards of metal-based nanoparticles on plants and environment, with special emphasis on ZnO NPs, TiO2 NPs, and AgNPs: A review. Environ. Adv. 2021, 6, 100128. [Google Scholar] [CrossRef]
- Palmer, C.M.; Guerinot, M.L. Facing the challenges of Cu, Fe and Zn homeostasis in plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samimi, S.; Maghsoudnia, N.; Eftekhari, R.B.; Dorkoosh, F. Lipid-based nanoparticles for drug delivery systems. Charact. Biol. Nanomater. Drug Deliv. 2019, 47–76. [Google Scholar] [CrossRef]
- Rajput, V.; Minkina, T.; Sushkova, S.; Behal, A.; Maksimov, A.; Blicharska, E.; Ghazaryan, K.; Movsesyan, H.; Barsova, N. ZnO and CuO nanoparticles: A threat to soil organisms, plants, and human health. Environ. Geochem. Health 2019, 42, 147–158. [Google Scholar] [CrossRef]
- AlShrouf, A. Hydroponics, aeroponic and aquaponic as compared with conventional farming. Am. Sci. Res. J. Eng. Technol. Sci. 2017, 27, 247–255. [Google Scholar]
- Nguyen, N.T.; McInturf, S.A.; Mendoza-Cózatl, D.G. Hydroponics: A Versatile System to Study Nutrient Allocation and Plant Responses to Nutrient Availability and Exposure to Toxic Elements. J. Vis. Exp. 2016, 54317. [Google Scholar] [CrossRef] [Green Version]
- Mousavi Kouhi, S.M.; Lahouti, M.; Ganjeali, A.; Entezari, M.H. Long-term exposure of rapeseed (Brassica napus L.) to ZnO nanoparticles: Anatomical and ultrastructural responses. Environ. Sci. Pollut. Res. Int. 2015, 22, 10733–10743. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Zhang, J.; Jia, L.; Li, Q.; Zhang, T.; Qiao, K.; Ma, S. Zinc induced phytotoxicity mechanism involved in root growth of Triticum aestivum L. Ecotoxicol. Environ. Saf. 2012, 86, 198–203. [Google Scholar] [CrossRef]
- Sagardoy, R.; Morales, F.; López-Millán, A.F.; Abadía, A.; Abadía, J. Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponics. Plant Biol. 2009, 11, 339–350. [Google Scholar] [CrossRef]
- Farghaly, F.A.; Radi, A.A.; Al-Kahtany, F.A.; Hamada, A.M. Impacts of zinc oxide nano and bulk particles on redox-enzymes of the Punica granatum callus. Sci. Rep. 2020, 10, 19722. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Bhadra, S.; Adegoke, A.; Bandyopadhyay, M.; Mukherjee, A. MWCNT uptake in Allium cepa root cells induces cytotoxic and genotoxic responses and results in DNA hyper-methylation. Mutat. Res. 2015, 774, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Wilson, M.; Hogstrand, C.; Maret, W. Picomolar Concentrations of Free Zinc(II) Ions Regulate Receptor Protein-tyrosine Phosphatase β Activity*. J. Biol. Chem. 2012, 287, 9322–9326. [Google Scholar] [CrossRef] [Green Version]
- Srivastav, A.; Ganjewala, D.; Singhal, R.K.; Rajput, V.D.; Minkina, T.; Voloshina, M.; Srivastava, S.; Shrivastava, M. Effect of ZnO Nanoparticles on Growth and Biochemical Responses of Wheat and Maize. Plants 2021, 10, 2556. [Google Scholar] [CrossRef]
- Liu, L.; Nian, H.; Lian, T. Plants and rhizospheric environment: Affected by zinc oxide nanoparticles (ZnO NPs). A review. Plant Physiol. Biochem. 2022, 185, 91–100. [Google Scholar] [CrossRef]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.-Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxidative Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.D.; Cress, W.A. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Naliwajski, M.; Skłodowska, M. The Relationship between the Antioxidant System and Proline Metabolism in the Leaves of Cucumber Plants Acclimated to Salt Stress. Cells 2021, 10, 609. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Nair, R. Plant response Strategies to engineered metal oxide nanoparticles: A review. In Phytotoxicity of Nanoparticles; Faisal, M., Saquib, Q., Alatar, A., Al-Khedhairy, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 377–393. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant Response to Engineered Metal Oxide Nanoparticles. Nanoscale Res. Lett. 2017, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.; Dubey, R.S. Effect of Cadmium on Soluble Sugars and Enzymes of their Metabolism in Rice. Biol. Plant. 2001, 44, 117–123. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [Green Version]
- Grune, T.; Reinheckel, T.; Davies, K.J. Degradation of oxidized proteins in mammalian cells. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1997, 11, 526–534. [Google Scholar] [CrossRef]
- Jung, T.; Höhn, A.; Grune, T. The proteasome and the degradation of oxidized proteins: Part II—Protein oxidation and proteasomal degradation. Redox Biol. 2014, 2, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc oxide nanoparticle-mediated changes in photosynthetic efficiency and antioxidant system of tomato plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Khan, J.; Brennand, D.M.; Bradley, N.; Gao, B.; Bruckdorfer, R.; Jacobs, M. 3-Nitrotyrosine in the proteins of human plasma determined by an ELISA method. Biochem. J. 1998, 330 Pt 2, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Pishkar, L.; Yousefi, S.; Iranbakhsh, A. Foliar application of Zinc oxide nanoparticles alleviates cadmium toxicity in purslane by maintaining nutrients homeostasis and improving the activity of antioxidant enzymes and glyoxalase system. Ecotoxicology 2022, 31, 667–678. [Google Scholar] [CrossRef]
- Cakmak, I. Tansley Review No. 111 Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Salah, S.M.; Yajing, G.; Dongdong, C.; Jie, L.; Aamir, N.; Qijuan, H.; Weimin, H.; Mingyu, N.; Jin, H. Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci. Rep. 2015, 5, 14278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USEPA. Ecological Effects Test Guidelines (OPPTS 850.4200): Seed Germination/Root Elongation Toxicity Test; EPA 712-C-96-132; USEPA: Washington, DC, USA, 1996. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Shemin, D. δ-Aminolevulinic acid dehydrase from Rhodopseudomonas spheroides. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1962; Volume 5, pp. 883–884. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sirota, T.V. Novel approach to the study od adrenaline auto-oxidation and its use for the measurements of superoxide dismutase activity. Vopr. Meditsinskoi Khimii 1999, 45, 263–272. [Google Scholar]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Arora, K.; Ahmad, R.; Srivastava, A.K. Purification and characterization of glutathione reductase (E.C. 1.8.1.7) from bovine filarial worms Setaria cervi. J. Parasit. Dis. 2013, 37, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Shigeoka, S.; Onishi, T.; Nakano, Y.; Kitaoka, S. Characterization and physiological function of glutathione reductase in Euglena gracilis z. Biochem. J. 1987, 242, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voloshina, M.; Rajput, V.D.; Minkina, T.; Vechkanov, E.; Mandzhieva, S.; Mazarji, M.; Churyukina, E.; Plotnikov, A.; Krepakova, M.; Wong, M.H. Zinc Oxide Nanoparticles: Physiological and Biochemical Responses in Barley (Hordeum vulgare L.). Plants 2022, 11, 2759. https://doi.org/10.3390/plants11202759
Voloshina M, Rajput VD, Minkina T, Vechkanov E, Mandzhieva S, Mazarji M, Churyukina E, Plotnikov A, Krepakova M, Wong MH. Zinc Oxide Nanoparticles: Physiological and Biochemical Responses in Barley (Hordeum vulgare L.). Plants. 2022; 11(20):2759. https://doi.org/10.3390/plants11202759
Chicago/Turabian StyleVoloshina, Marina, Vishnu D. Rajput, Tatiana Minkina, Evgeniy Vechkanov, Saglara Mandzhieva, Mahmoud Mazarji, Ella Churyukina, Andrey Plotnikov, Maria Krepakova, and Ming Hung Wong. 2022. "Zinc Oxide Nanoparticles: Physiological and Biochemical Responses in Barley (Hordeum vulgare L.)" Plants 11, no. 20: 2759. https://doi.org/10.3390/plants11202759
APA StyleVoloshina, M., Rajput, V. D., Minkina, T., Vechkanov, E., Mandzhieva, S., Mazarji, M., Churyukina, E., Plotnikov, A., Krepakova, M., & Wong, M. H. (2022). Zinc Oxide Nanoparticles: Physiological and Biochemical Responses in Barley (Hordeum vulgare L.). Plants, 11(20), 2759. https://doi.org/10.3390/plants11202759