
Soil Fertility Clock—Crop Rotation as a Paradigm in Nitrogen Fertilizer Productivity Control

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction—The Battle for Yield
- (1)
- Is the increase in nitrogen use efficiency (NUE) the real challenge for the increase in yields?
- (2)
- How is the effect of nitrogen-supporting nutrients (N–SNs) on efficiency of Nf manifested?
- (3)
- What is the required level of N–SNs in the soil, in order to maximize the Nf yield-forming effect?
2. A New Paradigm of Nitrogen Use Efficiency Control—The Basis of the Concept
- (1)
- The necessary condition is the actual yield, which is a function of the amount of available N in the rooting zone of the currently grown crop.
- (2)
- The sufficient condition is the yield-forming functions of nutrients other than N to support the action of N by increasing its uptake and its use by the currently grown plant.
- (1)
- A crop plant in a well-defined geographic area, provided stable environmental and nutritional conditions, can reach Yattmax.
- (2)
- The key production factor is N, present in the soil or/and supplied to the plant as fertilizer (natural, manure; mineral, Nf).
- (3)
- All other nutrients, called nitrogen supporting nutrients (N-SNs), affect the Yattmax, in relation to their relative deficiency in available form in the plant rooting zone.
- Actual yield:
- 2.
- Nitrogen Efficiency (EN):
3. Nitrogen—A Unique and Critical Factor in Plant Production
4. Nitrogen-Supporting Nutrients
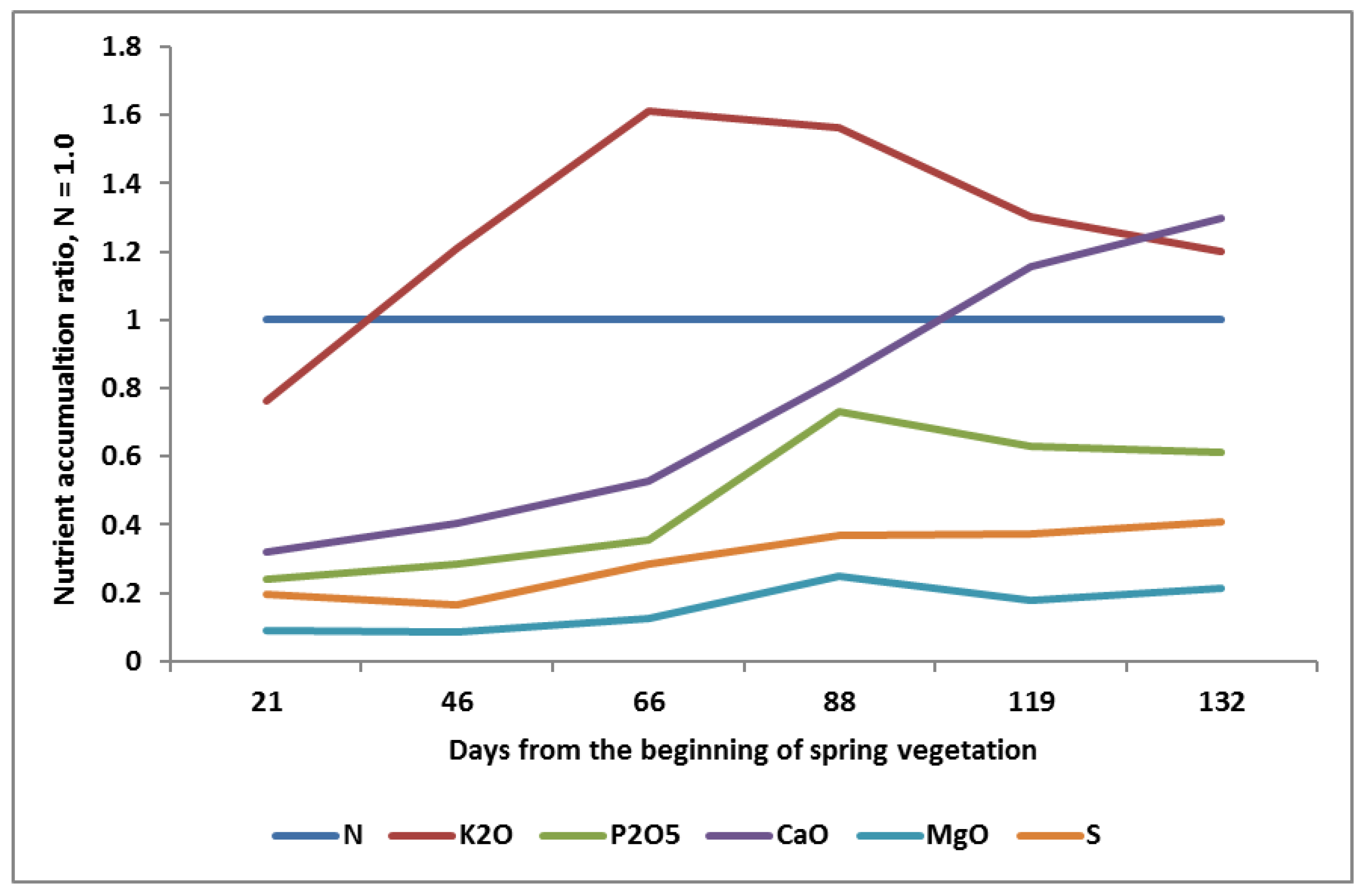
- (1)
- The higher accumulation of K over N during the whole season of WOSR growth:
- Starting with the rosette stage in Spring;
- Achieving the maximum K uptake over N at the full flowering stage (K2O:N as 1:1.6);
- Declining from flowering to maturity (K2O:N as 1:1).
- (2)
- Slow early growth in P uptake, continued up to the inflorescence stage (BBCH 50), followed by rapid ingrowth, lasting until the full flowering stage (BBCH 65), and then smoothly decreasing up to maturity.
- (3)
- A similar pattern for Mg as for P, but at a much lower level.
- (4)
- A spectacular pattern of Ca accumulation. Its uptake increases sharply at inflorescence, reaching a maximum at the end of the pod growth stage.

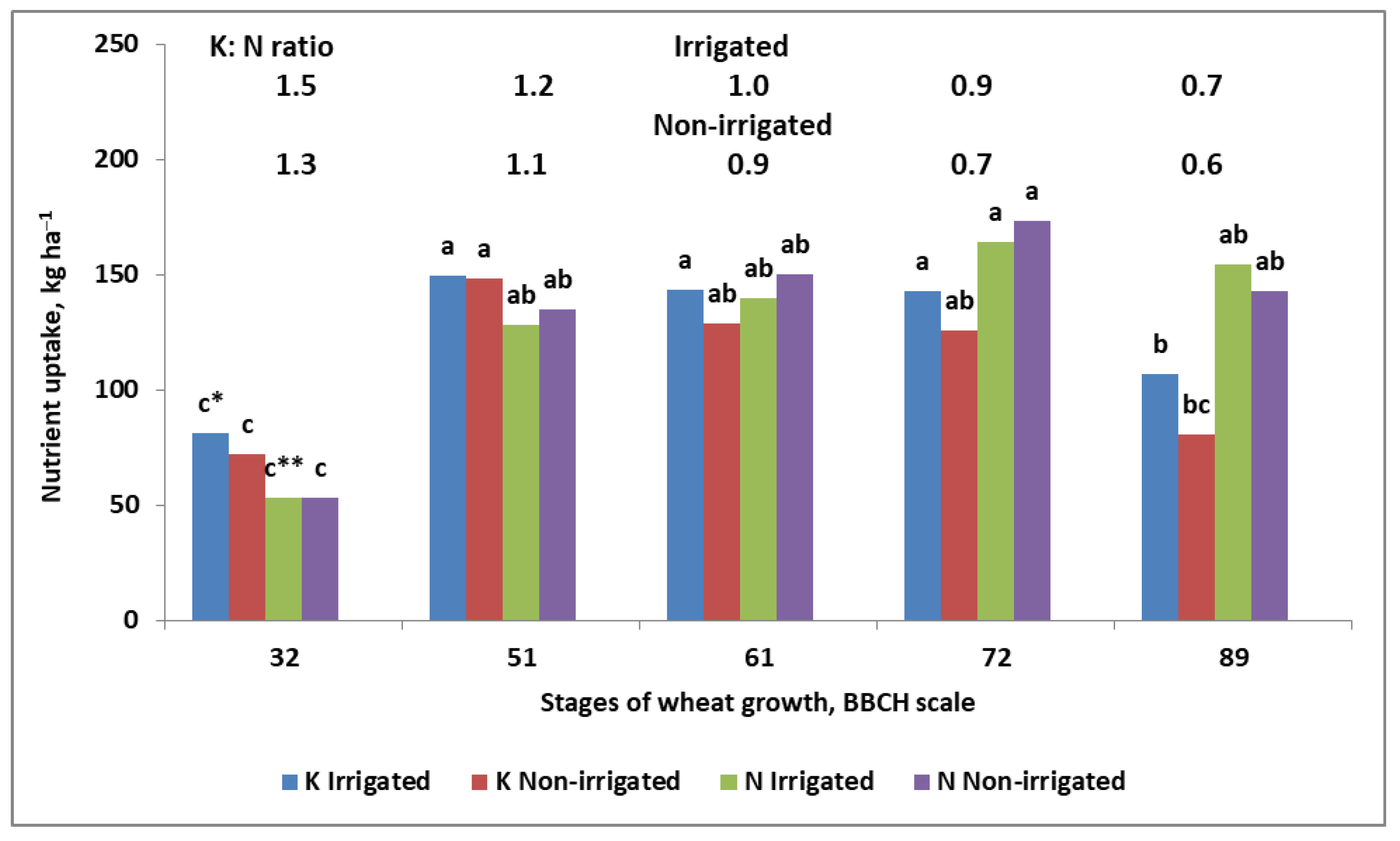
5. Potassium
- (1)
- Higher demand for K in the linear phase of growth;
- (2)
- Much smaller root system, especially root length density.
6. Phosphorus
7. Efficient Nitrogen Management—The Soil Fertility Clock Concept
7.1. Definition of the Concept
7.2. Maximum Attainable Yield—A Farm Production Goal
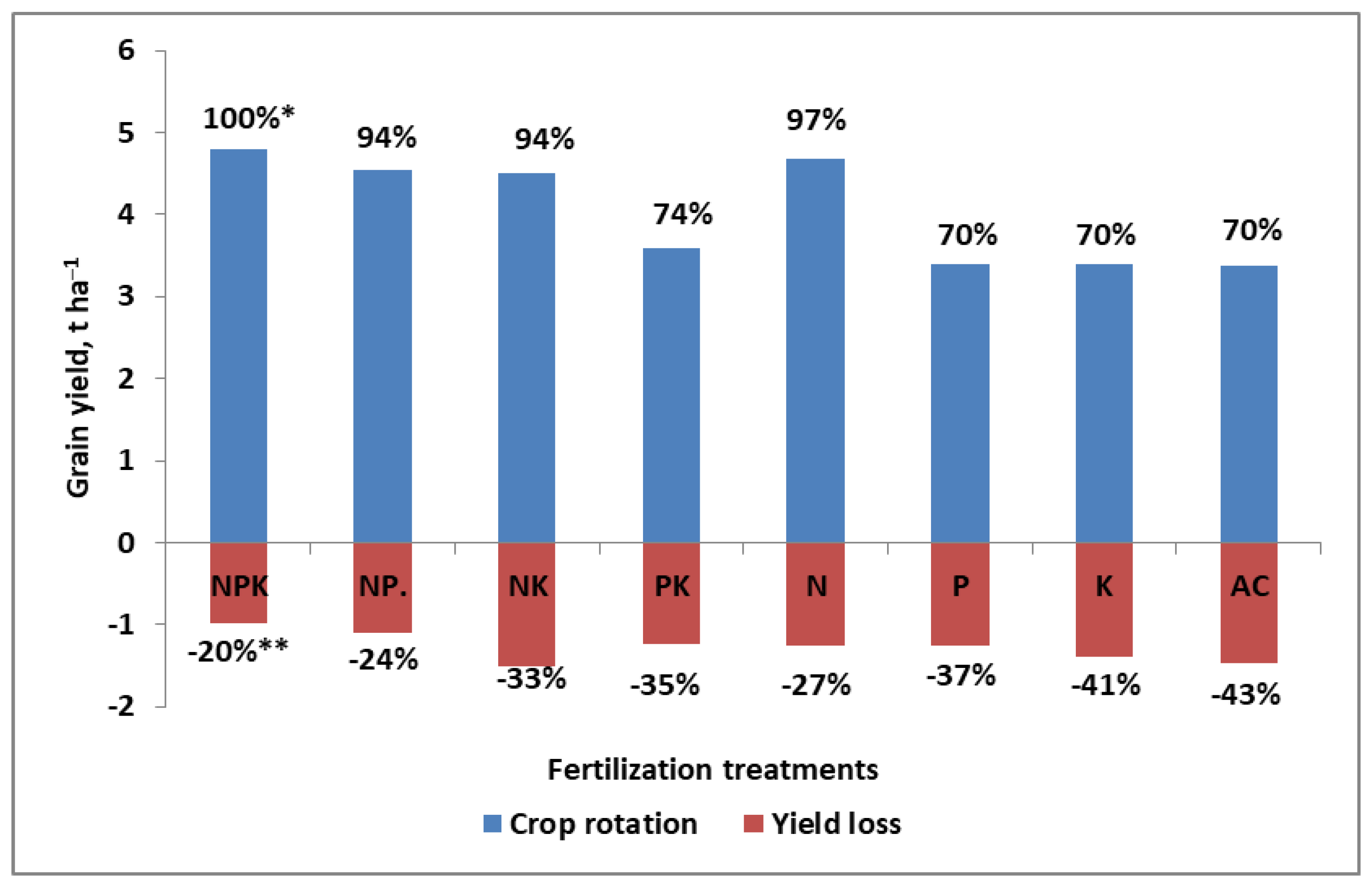
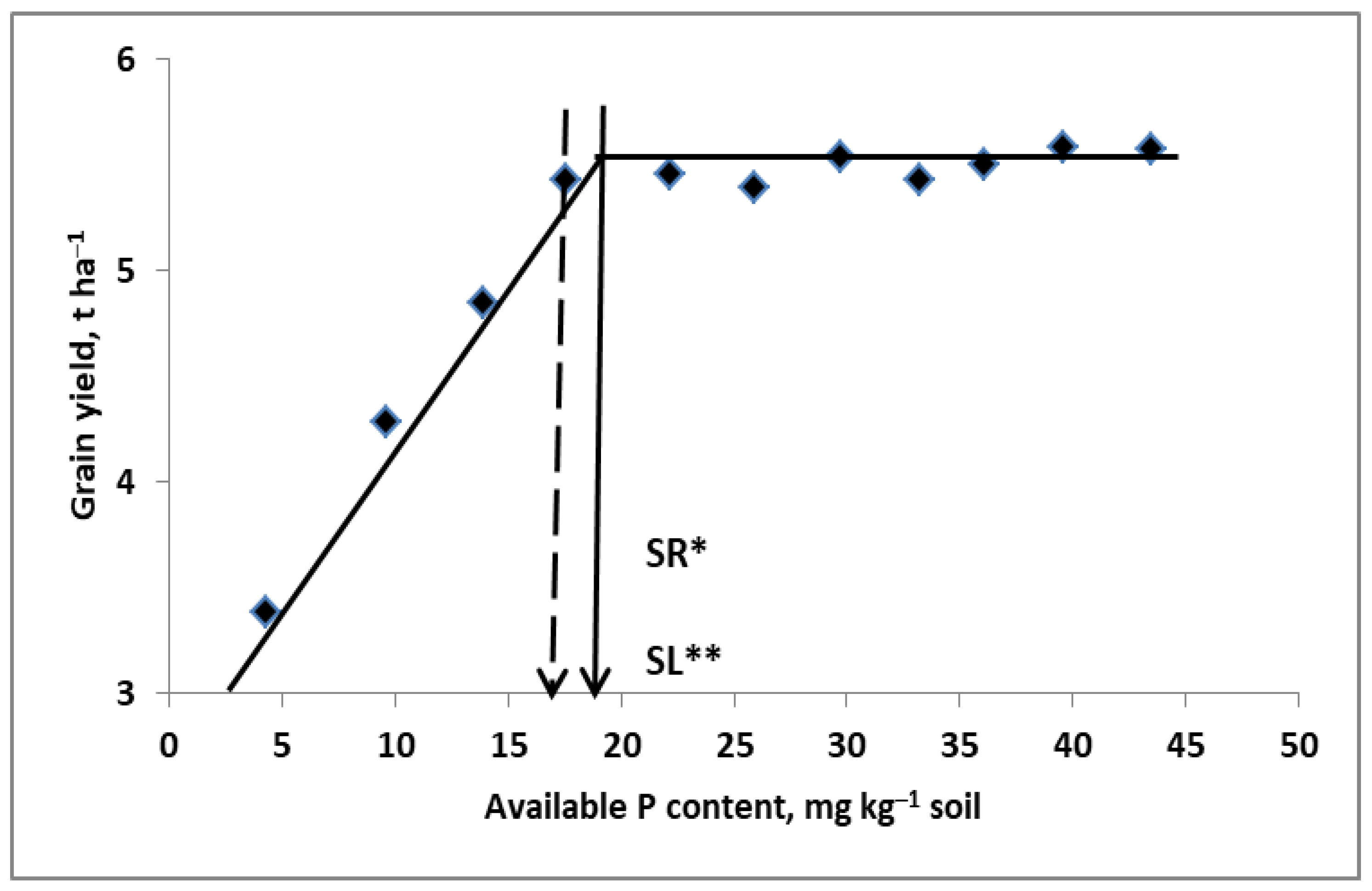
- (1)
- The content of available K (in the medium range) was enough to achieve the highest grain yield.
- (2)
- The interaction of K and N was observed, regardless of the course of the weather.
- (3)
- If the available K in the soil (being in the high range) is excessive, the yield will decrease.

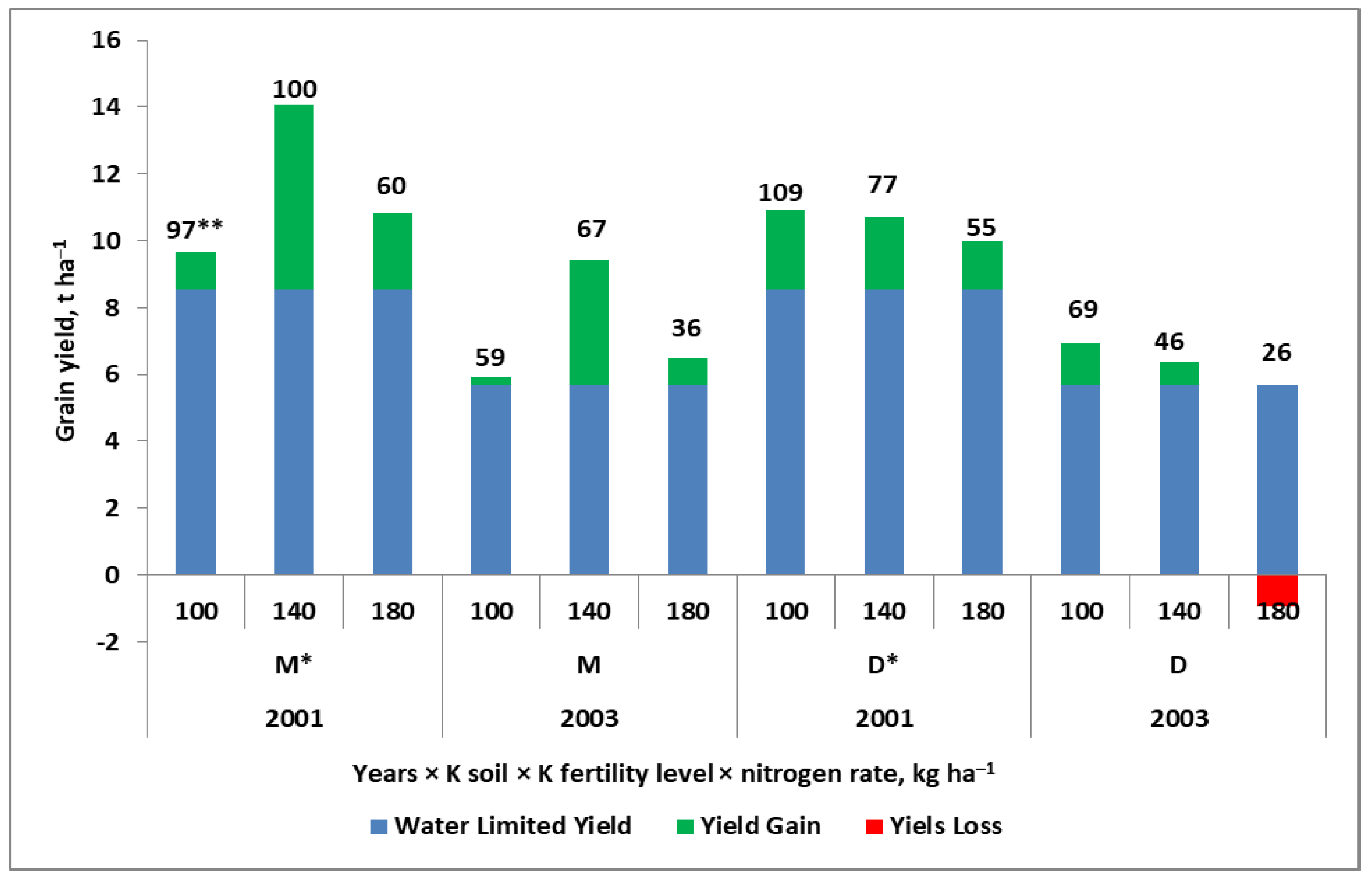
7.3. Factors Affecting N Fertilizer Use Efficiency
- (1)
- Farm organization and management;
- (2)
- Agronomic factors (e.g., cropping sequence ≈ crop rotation, soil tillage, seed bed preparation, cultivar, sowing date, harvest date);
- (3)
- Plant protection treatments, preventing yield reduction due to pathogens and pests;
- (4)
- Fertilizing treatments aimed at the correction of soil fertility;
- (5)
- Fertilizing treatments aimed at the in-season correction of the nutritional status of the grown plant.
- (1)
- The use of natural sources of N available on the field and farm. This leads to a reduction in the need for Nf.
- (2)
- Biological subsoil amelioration by the strong root systems of dicots. Expected agronomic effects lead to:
- Mobilization of the soil nutrient resources (root exudates, mycorrhiza);
- Increased soil water capacity, resulting in better infiltration of rainwater;
- Increased exploration of the subsoil (i.e., growth of cereal roots in the root pores of dicots).
- (3)
- A narrower C:N ratio in manure and residues of legumes. Both sources of organic matter have a positive effect on humus formation and content in the soil.
- (4)
- The exploitation of soil nutrient resources within and in the soil profile is more sustainable, both qualitatively and quantitatively.
8. An Efficient System for Management of N-SNs—Principles of the Soil Fertility Clock
8.1. State of K and P Fertility Level and Food Production
8.2. Management of Soil Fertility—Oriented to Cropping Sequence
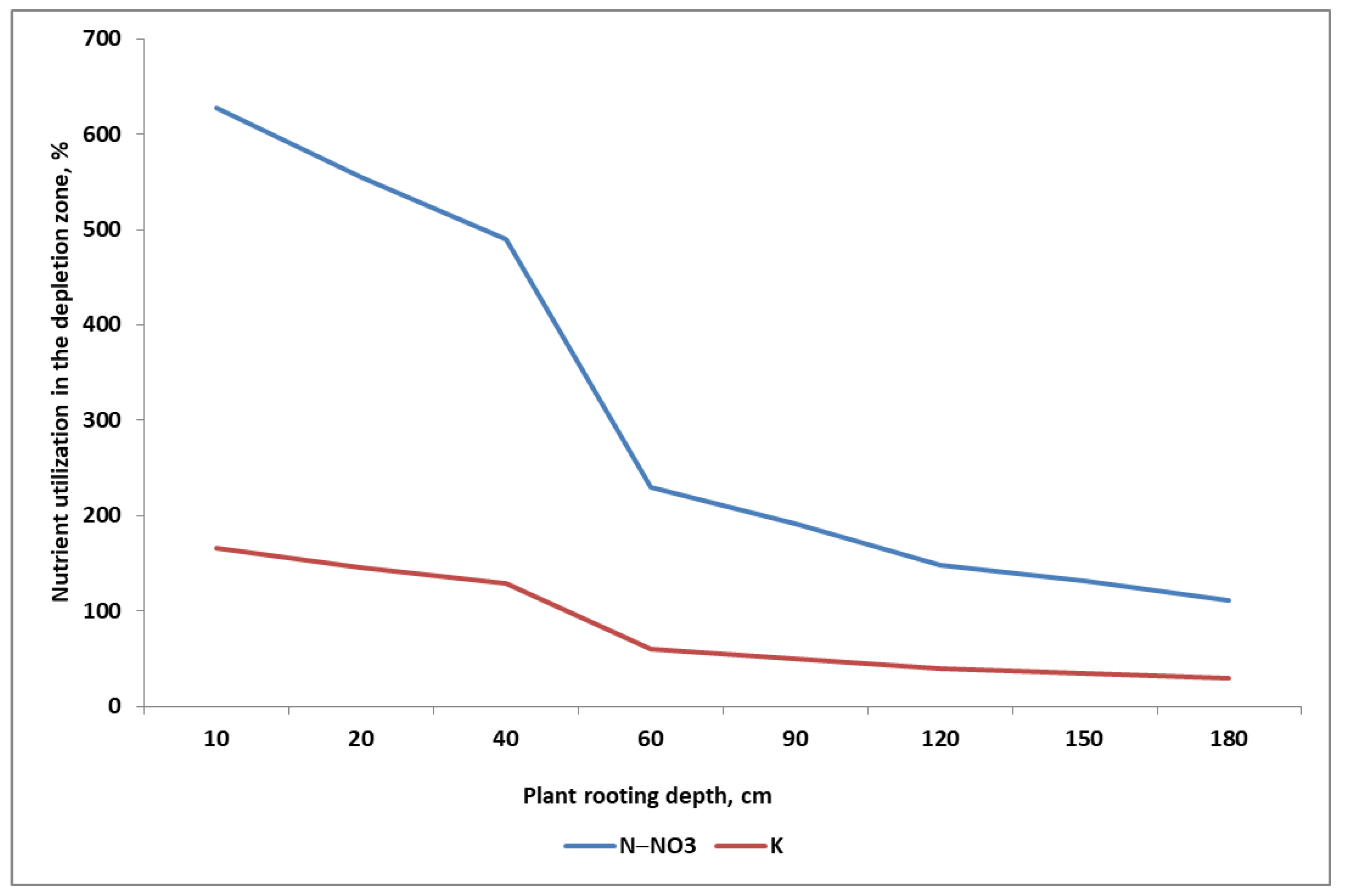
- (1)
- The nutrients are exploited from the whole root zone of the currently grown crop plant;
- (2)
- The faster uptake of nitrates than K+ by the fibrous roots of sugar beet means that a significant part of the low-mobility nutrients in the soil will not be taken up (i.e., not used up in the growing season).

- During the growing season;
- In the course of crop rotation.
- (1)
- Annual crop plants should be cultivated in a fixed sequence (i.e., the crop rotation).
- (2)
- Cereals have, as a rule, lower requirements for K, but higher requirements for P, compared to non-cereal crops.
- (3)
- Dicotyledonous plants have higher requirements for K than cereals.
- (4)
- The architecture of the root system of cereals is, as a rule, extensive compared to dicotyledonous plants. Consequently, cereals are less sensitive to the level of P and K fertility.
- (5)
- The distribution of low-mobility nutrients varies with depth.
- (6)
- Plants during the growing season differ significantly in the critical stages of nutrient requirement:
- Seed crops show critical periods, in terms of P requirements, during the vegetative (minor one) and reproductive (main one) periods of growth;
- All crops are sensitive to K during the linear phase of growth.
- (7)
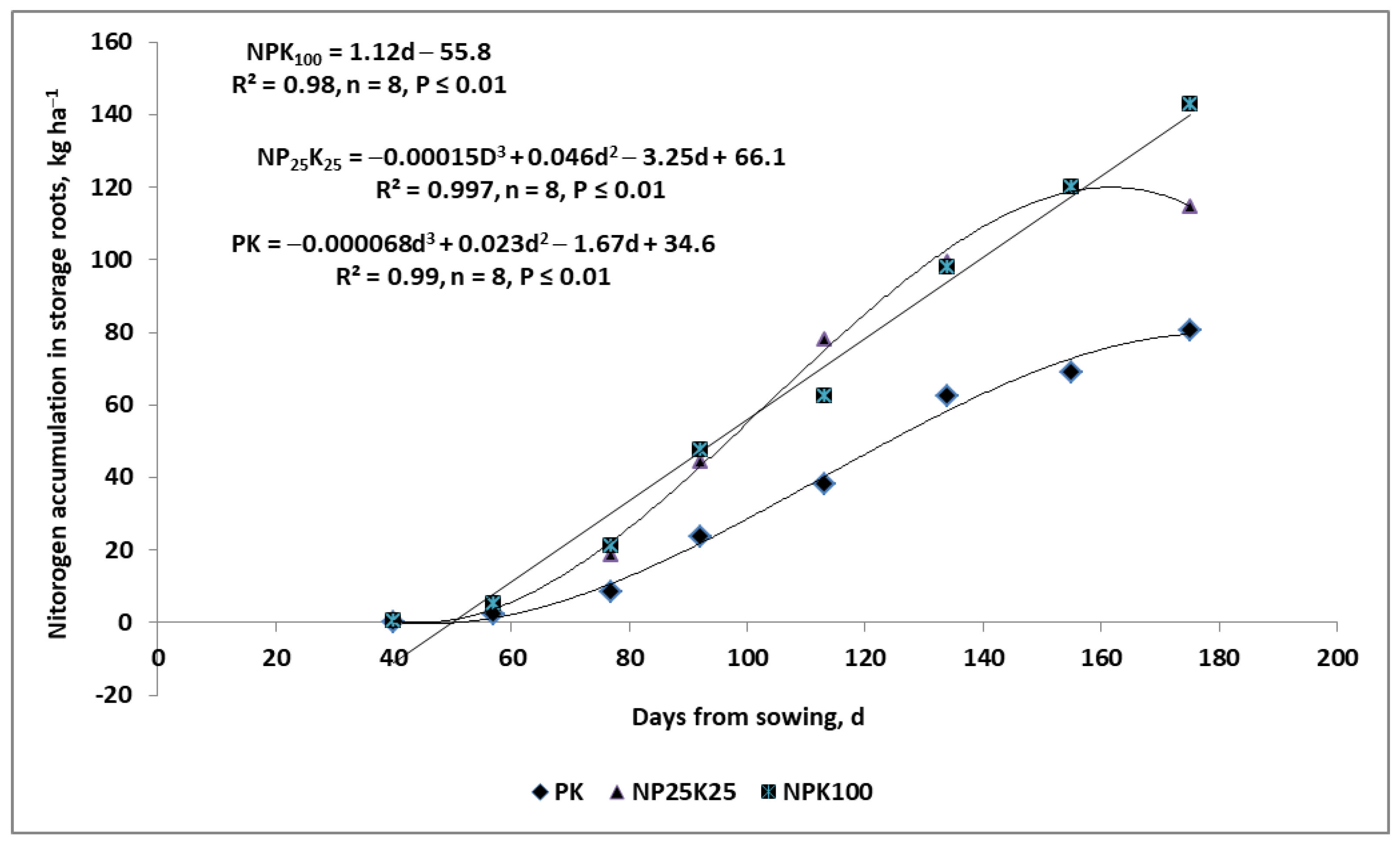
- The critical period for N requirements by a seed crop is related to stages of seed/grain density formation.
- (8)
- The key yield-forming function of P in all crops is to accelerate the early rate of the plant growth.
- (9)
- The exploitation of P resources by seed crops, accumulated in vegetative parts before flowering, depends on seed/grain density.
- (10)
- The yield-forming function of K in
- Seed crops is to strengthen N action;
- Dicotyledonous crops is acceleration of the early rate of the plant growth (mostly up to the rosette stage).
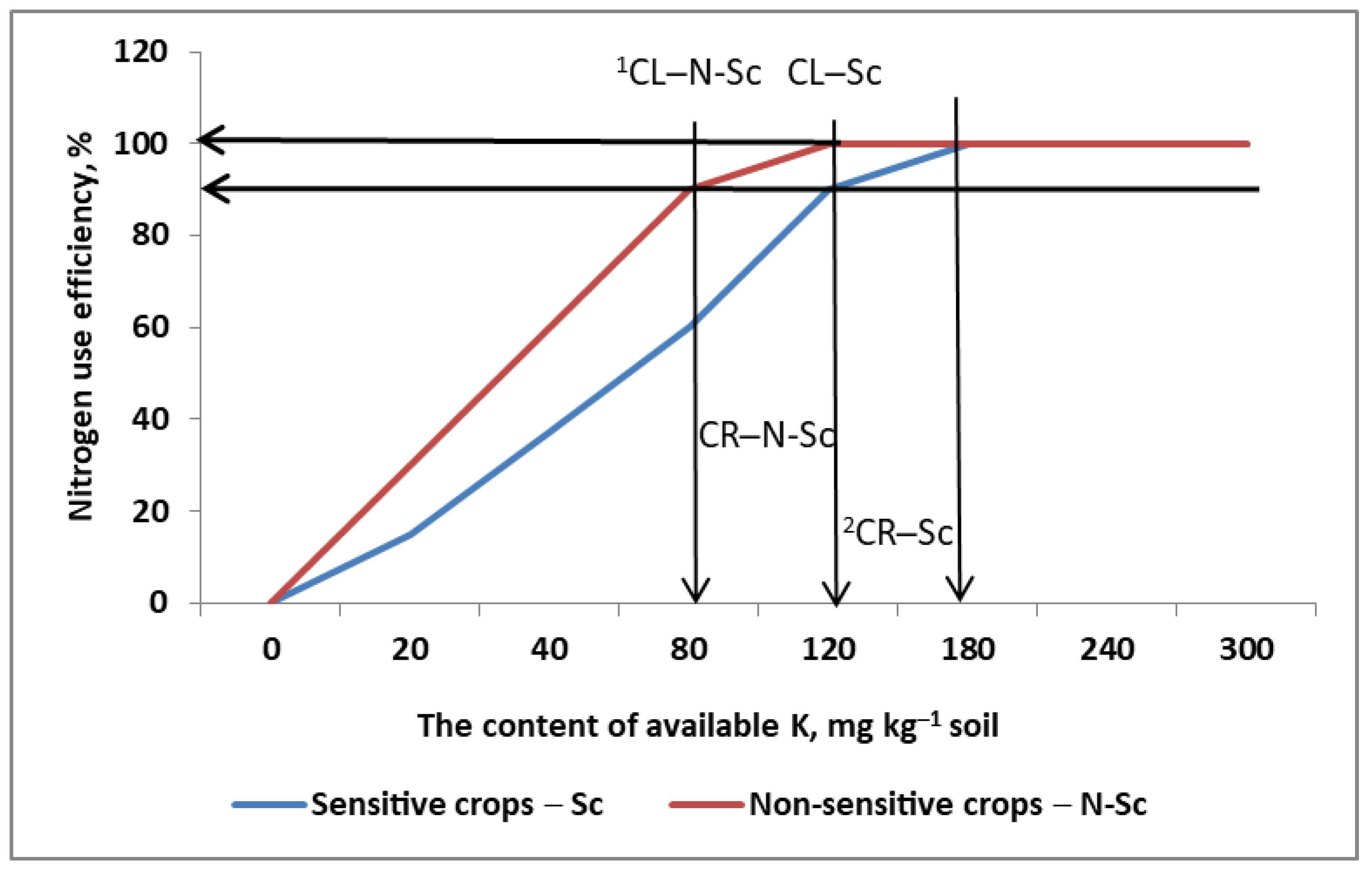
- (1)
- Critical soil fertility is the value or range of a soil nutrient’s content that is sufficient to provide it in the appropriate amount to the plant most sensitive to its supply in a given crop rotation.
- (2)
- Other, non-sensitive plants in the given crop rotation create the necessary time-frame for recovery of its original critical content.
- (3)
- The content of a specific nutrient cannot be a limiting factor in N uptake and utilization for any crop grown. Its fractional use efficiency, regardless of the actual plant in crop rotation, is ≤1.0.
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. BioScience 2017, 67, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Beltran-Peña, A.; Rosa, L.; D’Odorico, P. Global food self-sufficiency in the 21st century under sustainable intensification of agriculture. Environ. Res. Lett. 2020, 15, 095004. [Google Scholar] [CrossRef]
- Röös, E.; Bajželj, B.; Smith, P.; Patel, M.; Little, D.; Garnett, T. Greedy or needy? Land use and climate impacts of food in 2050 under different livestock futures. Glob. Environ. Chang. 2017, 47, 1–12. [Google Scholar] [CrossRef]
- Conijn, J.G.; Bindraban, P.S.; Schröder, J.J.; Jongschaap, R.E.E. Can our global food system meet food demand within planetary boundries? Agric. Ecosys. Environ. 2018, 251, 244–256. [Google Scholar] [CrossRef]
- FAO. Impact of the Ukraine-Russia Conflict on Global Food Security and Related Matters under the Mandate of the Food and Agriculture Organization of the United Nation (FAO). CL 170/6. May 2022. Available online: https://www.fao.org/3/nj164en/nj164en.pdf (accessed on 20 August 2022).
- Strubenhoff, H. The War in Ukraine Triggered a Global Food Shortage. 2022. Available online: www.brookings.edu/blog/future-development/2022/06/14/the-war-in-ukraine-triggered-a-global-food-shortage/ (accessed on 20 August 2022).
- Alexander, P.; Brown, C.; Ameth, A.; Finnigan, J. Human appropriation for land and food: The role of diet. Glob. Environ. Chang. 2016, 41, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Gomiero, T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.; Coe, M.; Walker, W.; Verchot, L.; Vandecar, K. The unseen effects of deforestation: Biophysical effects on climate. Front. For. Glob. Chang. 2022, 5, 756115. [Google Scholar] [CrossRef]
- Silver, W.L.; Perez, T.; Mayer, A.; Jones, A.R. The role of soil in the contribution of food and feed. Philos. Trans. R. Soc. B 2021, 376, 20200181. [Google Scholar] [CrossRef]
- Jaggard, K.W.; Qi, A.; Ober, E.S. Possible changes to arable crop yields by 2050. Philos. Trans. R. Soc. B 2010, 365, 2835–2851. [Google Scholar] [CrossRef] [Green Version]
- Harvey, M.; Pilgrim, S. The new competition for land: Food, Energy, and climate change. Food Policy 2011, 36, 40–50. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- The Royal Society. Reaping the Benefits: Science and the Sustainable Intensification of Global Agriculture; RS Policy Document 11/09; The Royal Policy Center: London, UK, 2009; p. 86. [Google Scholar]
- Łukowiak, R.; Grzebisz, W.; Ceglarek, J.; Podolski, A.; Kaźmierowski, C.; Piekarczyk, J. Spatial variability of yield and nitrogen indicators—A crop rotation approach. Agronomy 2020, 10, 1959. [Google Scholar] [CrossRef]
- Pradhan, P.; Fischer, G.; van Velthuizen, H.; Reusser, D.E.; Kropp, J.P. Closing yield gaps: How sustainable can we be? PLoS ONE 2015, 10, e0129487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houlton, B.Z.; Almaraz, M.; Aneja, V.; Austin, V.T.; Bai, E.; Cassman, K.G.; Compton, J.E.; Davidson, E.A.; Erisman, J.W.; Galloway, J.N.; et al. A world co-benefits: Solving the global nitrogen challenge. Earths Future 2019, 7, 865–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Schnitkey, G.; Paulson, N.; Swanson, K.; Colussi, J.; Baltz, J.; Zulauf, C. Nitrogen fertilizer prices and supply in light of the Ukraine-Russia conflict. Farmdoc Dly. 2022, 12, 45. Available online: https://farmdocdaily.illinois.edu/2022/04/nitrogen-fertilizer-prices-and-supply-in-light-of-the-ukraine-russia-conflict.html (accessed on 20 August 2022).
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenza, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef]
- Stewart, W.M.; Roberts, T.L. Food security and the role of fertilizer in supporting it. Procedia Eng. 2012, 46, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Poutanewn, K.S.; Karlund, A.O.; Gomez-Gallego, C.; Johansson, D.P.; Scheers, N.M.; Marklinder, I.M.; Eriksen, A.K.; Silventoinen, P.C.; Nordlund, E.; Sozer, N.; et al. Grains—A major source of sustainable protein for health. Nutr. Rev. 2022, 80, 1648–1663. [Google Scholar] [CrossRef]
- Kong, L.G.; Xie, Y.; Hu, L.; Feng, B.; Li, S.D. Remobilization of vegetative nitrogen to developing grain in wheat (Triticum aestivum L.). Field Crops Res. 2016, 196, 134–144. [Google Scholar] [CrossRef]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Preanthesis biomass accumulation and plant organs defines yield components in wheat. Eur. J. Agron. 2016, 81, 15–26. [Google Scholar] [CrossRef]
- Klepper, B.; Rickman, R.W.; Waldman, S.; Chevalier, P. The physiological life cycle of wheat: Its use in breeding and crop management. Euphytica 1998, 100, 341–347. [Google Scholar] [CrossRef]
- Slafer, G.A.; Elia, M.; Savin, R.; Garcia, G.A.; Terrile, I.I.; Ferrante, A.; Miralles, D.J.; González, F.G. Fruiting efficiency: An alternative trait to further rise in wheat yield. Food Energy Secur. 2015, 4, 92–109. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global maize production, consumption and trade: Trends and R&D implications. Food Secur. 2022, 5, 345–360. [Google Scholar] [CrossRef]
- Subedi, K.; Ma, B. Nitrogen uptake and partitioning in stay-green and leafy maize hybrids. Crops Sci. 2005, 45, 740–747. [Google Scholar] [CrossRef]
- Grzebisz, W.; Wrońska, M.; Diatta, J.B.; Szczepaniak, W. Effect of zinc foliar application at early stage of maize growth on the patterns of nutrients and dry matter accumulation by the canopy. Part II. Nitrogen uptake and dry matter accumulation patterns. J. Elementol. 2008, 13, 29–39. [Google Scholar]
- Andre, C.M.; Legay, S.; Iammarino, C.; Ziebel, J.; Guignard, C.; Larondelle, Y.; Hausman, J.-F.; Evers, D.; Miranda, L.M. The potato in the human diet: A complex matrix with potential health benefits. Potato Res. 2014, 57, 201–214. [Google Scholar] [CrossRef]
- Jackson, S.D. Multiple signaling pathway control tuber induction in potato. Plant Physiol. 1999, 119, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Engels, C.; Marschner, H. Allocation of photosynthate to individual tubers of Solanum tuberosum L. III. Relationship between growth rate of individual tubers, tuber weight and stolon growth prior to tuber initiation. J. Exp. Bot. 1986, 37, 1813–1822. [Google Scholar] [CrossRef]
- Grzebisz, W.; Potarzycki, J. The in-season nitrogen concentration in the potato tuber as the yield driver. Agron. J. 2020, 112, 1287–1308. [Google Scholar] [CrossRef]
- Lewis, N.S.; Nocera, D.G. Powering the planet: Chemical challenges in solar energy utilization. Proc. Natl. Acad. Sci. USA 2006, 103, 15729–15735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cammarano, D.; Zha, H.; Wilson, L.; Li, Y.; Batchelor, W.D.; Miao, Y. A remote sensing–based approach to management zone delineation in small scale farming systems. Agronomy 2020, 10, 1767. [Google Scholar] [CrossRef]
- Licker, R.; Johnston, M.; Foley, J.A.; Barford, C.; Kucharik, C.J.; Monfreda, C.; Ramankutty, N. Mind the gap: How do climate and agricultural management explain the „yield gap” of croplands around the world? Glob. Ecol. Biogeogr. 2010, 19, 769–782. [Google Scholar] [CrossRef]
- Iizumi, T.; Ramankutty, N. How do weather and climate influence cropping area and intensity? Glob. Food Secur. 2015, 4, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Rabbinge, R. The ecological background of food production. In Crop Production and Sustainable Agriculture; John Wiley & Sons: Chichester, NY, USA, 1993; pp. 2–29. [Google Scholar]
- Rubio, G.; Zhu, J.; Lynch, J. A critical test of the prevailing theories of plant response to nutrient availability. Am. J. Bot. 2003, 90, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Grafton, R.Q.; Williams, J.; Jiang, Q. Food and water gaps to 2050: Preliminary results from the global food and water systems (GFWS) platform. Food Secur. 2015, 7, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, T.R.; Rufty, T.W. Nitrogen and water resources commonly limit crop yield increases, not necessarily plant genetics. Glob. Food Secur. 2012, 1, 94–98. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Elsevier Ltd.: Amsterdam, The Netherlands; Academic Press: London, UK, 1995; p. 899. [Google Scholar]
- Reid, R.; Hayes, J. Mechanisms and control of nutrient uptake in plants. Int. Rev. Cytol. 2003, 229, 73–114. [Google Scholar]
- Kant, S.; Seneweera, S.; Rodin, J.; Materne, M.; Burch, D.; Rothstein, S.J.; Spangenberg, G. Improving yield potential in crop under elevated CO2: Integrating the photosynthetic and nitrogen utilization efficiencies. Front. Plant Sci. 2012, 3, 162. [Google Scholar] [CrossRef] [Green Version]
- Parry, A.J.; Andralojc, P.J.; Scales, J.C.; Salvucci, M.E.; Carmo-Silva, A.E.; Alonso, H.; Whitney, S.M. Rubisco activity and regulation as targets for crop improvement. J. Exp. Bot. 2013, 6493, 717–730. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Milo, R. The global mass and average rate of rubisco. Proc. Natl. Acad. Sci. USA 2019, 116, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giehl, R.F.H.; Gruber, B.D.; von Wirén, N. It’s time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, J.; Smalle, J.A. Auxin/cytokinin antagonistic control of the shoot/root growth ratio and its relevance for adaptation to growth and nutrient deficiency stress. Int. J. Mol. Sci. 2022, 23, 1933. [Google Scholar] [CrossRef] [PubMed]
- Guan, P. Dancing with hormones: A current perspective of nitrate signaling and regulation in Arabidopsis. Front. Plant Sci. 2017, 8, 1697. [Google Scholar] [CrossRef]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanism of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Chen, D.; Schnurbusch, T. Plant and floret growth at distinct developmental stages during the stem elongation phase in wheat. Front. Plant. Sci. 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Potarzycki, J. Influence of balanced fertilization on nutritional status of maize at anthesis. Fertil. Fertil. 2010, 39, 90–108. [Google Scholar]
- Ciampitti, I.A.; Camberato, J.J.; Murrel, S.T.; Vyn, T.J. Maize nutrient accumulation and partitioning in response to plant density and nitrogen rate. I. Macronutrients. Agron. J. 2013, 10593, 7830. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L. Agriculture, plant physiology, and human population growth: Past, present, and future. Theor. Exp. Plant Physiol. 2013, 25, 167–181. [Google Scholar] [CrossRef] [Green Version]
- White, C.; Sylvester-Bradley, R.; Berry, P.M. Root length densities of UK wheat and oilseed rape crops with implications for water capture and yield. J. Exp. Bot. 2015, 66, 2293–2303. [Google Scholar] [CrossRef] [Green Version]
- Fradgley, N.; Evans, G.; Biernaskie, J.M.; Cockram, J.; Marr, E.C.; Oliver, A.G.; Ober, E.; Jones, H. Effects of breeding history an crop management on the root architecture of wheat. Plant Soil 2020, 452, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Milach, S.C.K.; Federizzi, L.C. Dwarfing genes in plant improvement. Adv. Agron. 2001, 73, 35–63. [Google Scholar]
- Lynch, J.L. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kell, D.B. Breeding crop plants with deep roots: Their role in sustainable carbon, nutrient and water sequestration. Ann. Bot. 2011, 108, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. (Ed.) Marchnerr’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; p. 672. [Google Scholar]
- Benton Jones, J., Jr. Agronomic Handbook: Management of Crops, Soils and Their Fertility; CRC Press LLC: Boca Raton, FL, USA, 2003; 450p. [Google Scholar]
- Wang, Y.; Wu, W.-H. Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency. Curr. Opin. Plant Biol. 2015, 24, 46–52. [Google Scholar] [CrossRef]
- White, P.J.; Brown, P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Lyu, S.; Wei, X.; Chen, J.; Wang, X.; Pan, D. Titanium as a beneficial element for crop production. Front. Plant Sci. 2017, 8, 597. [Google Scholar] [CrossRef] [Green Version]
- Blecharczyk, A. The reaction of winter rye and spring barley to the plant crop rotation system and fertilization in a long-term experiment. Rocz. Akad. Rol. Pozn. 2002, 236, 128, (In Polish with English Summary). [Google Scholar]
- Buczko, U.; van Laak, M.; Eichler-Löbermann, B.; Gans, W.; Merbach, I.; Panten, K.; Peiter, E.; Reitz, T.; Spiegel, H.; von Tucher, S. Evaluation of the yield response to phosphorus fertilization based on meta-analysis of long term experiments. Ambio 2018, 47, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Johnston, A.E.; Poulton, P.R. The importnance of long-term experiments in agriculture: Their management to ensure continued crop production and soil fertility: The Rothamsted experience. Eur. J. Soil Sci. 2018, 69, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Janssen, B.H. Crop Yields and NPK Use Efficiency of a Long-Term Experiment on Former Seas Bottom in The Netherlands; Wgageningen University: Wageningen, The Netherlands, 2017; p. 155. ISBN 978-94-6343-130-9. [Google Scholar]
- Plant Health Association California. Western Fertilizer Handbook, 10th ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2022; 371p. [Google Scholar]
- Barłóg, P.; Grzebisz, W.; Diatta, J.B. Effect of timing and nitrogen fertilizers on nutrients content and uptake by winter oilseed rape. Part II. Dynamics of nutrient uptake. Chem. Agric. 2005, 6, 113–123. [Google Scholar]
- Barraclough, P.B. Root growth, macro-nutrient uptake dynamics and soil fertility requirements of a high-yielding winter oilseed rape crop. Plant Soil 1989, 119, 59–70. [Google Scholar] [CrossRef]
- Hirzel, J.; Undurraga, P. Nutritional Management of cereals cropped under irrigation conditions. In Crop Production; Goyal, A., Asil, M., Eds.; InTech: London, UK, 2013; pp. 99–130. [Google Scholar]
- Grzebisz, W. Technology of Crop Plants Fertilization–Yield Physiologu. Part II. Cereals and Maize; PWRiL: Poznań, Poland, 2012; 279p. (In Polish) [Google Scholar]
- Grzebisz, W.; Diatta, J. Constrains and solutions to maintain soil productivity, a case study from Central Europe. In Soil Fertility Improvement and Integrated Nutrient Management—A Global Perspective; Whalen, J., Ed.; InTech: London, UK, 2012; pp. 159–183. [Google Scholar]
- Bender, R.R.; Haegele, J.W.; Below, F.E. Nutrient uptake, partitioning, and remobilization in modern soybean varieties. Agron. J. 2015, 107, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrel, T.S.; Mikkelsen, R.L.; Sulewski, G.; Norton, R.; Thompson, M.L. Improving Potassium Recommendations for Agricultural Crops; Springer Nature Switzerland AG: Cham, Switzerland, 2021; 455p. [Google Scholar]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Marschner, H.; Kirkby, E.A.; Cakmak, J. Effect of mineral nutrition on shoot-root partitioning of photo-assimilates and cycling of mineral nutrients. J. Exp. Bot. 1996, 47, 1255–1263. [Google Scholar] [CrossRef]
- Yawson, D.O.; Amoach, K.K.; Asare, P.A.; Techie-Menson, J.; Afutu, E.; Atiah, K.; Sanieri, A.K.; Sare-Larbi, S.; Adu, M.O. A meta-analysis of modifications of root system traits of crop plants to potassium (K) deprivation. In Plant Roots; Yildirim, E., Turan, M., Ekinci, M., Eds.; InTech: London, UK, 2021; 27p. [Google Scholar]
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of potassium fertilization on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374. [Google Scholar] [CrossRef]
- Hu, Q.Q.; Shu, J.-Q.; Li, W.-M.; Wang, G.-Z. Role of auxin and nitrate signaling in the development of root system architecture. Front. Plant Sci. 2021, 12, 690363. [Google Scholar] [CrossRef]
- Duan, J.; Wu, Y.; Zhou, Y.; Ren, X.; Shao, Y.; Feng, W.; Zhu, Y.; Wang, Y.; Guo, T. Grain number response to pre–anthesis dry matter and nitrogen in improving wheat yield in the Huang–Huai Plain. Sci. Rep. 2018, 8, 7126. [Google Scholar]
- White, P.J.; Bell, M.J.; Djalovic, I.; Hinsinger, P.; Rengel, Z. Potassium use efficiency of plants. In Improving Potassium Recommendations for Agricultural Crops; Murrel, T.S., Mikkelsen, R.L., Sulewski, G., Norton, R., Thompson, M.L., Eds.; Springer Nature Switrzerland AG: Cham, Switzerland, 2021; pp. 119–146. [Google Scholar]
- Noordwijk, M. Functional interpretation of root densities in the field for nutrient and water uptake. Root Ecol. Pract. Appl. 1983, 207–226. Available online: https://edepot.wur.nl/261865 (accessed on 25 August 2022).
- Fan, J.; McConkey, B.; Wang, H.; Janzen, H. Root distribution by depth for temperate agricultural crops. Field Crops Res. 2016, 189, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Kuchenbuch, R.; Claassesn, N.; Jungk, A. Potassium availability in relation to soil moisture. II. Calculation by means of a mathemantical model simulation model. Plant Soil 1986, 95, 233–243. [Google Scholar] [CrossRef]
- Grzebisz, W.; Szczepaniak, W.; Potarzycki, J.; Łukowiak, R. Sustainable management of soil potassium—A crop rotation oriented concept. In Soil Fertility; Issaka, R.N., Ed.; InTech: London, UK, 2012; 38p. [Google Scholar]
- Finck, A. Dünger und Düngung; VCH Verlag-AG: Basel, Switzerland, 1992; 488p. [Google Scholar]
- Grzebisz, W.; Łukowiak, R. Nitrogen gap amelioration is a core for sustainable intensification of agriculture—A concept. Agronomy 2021, 11, 419. [Google Scholar] [CrossRef]
- Pathak, H.; Aggarwal, P.K.; Roetter, R.; Kalra, N.; Bandyopadhaya, S.K.; Prasad, S.; Van Keulen, H. Modelling the quantitative evaluation of soil nutrient supply, nutrient use efficiency, and fertilizer requirements of wheat in India. Nutr. Cycl. Agroecosys. 2003, 65, 105–113. [Google Scholar] [CrossRef]
- Grzebisz, W.; Szczepaniak, W.; Grześ, S. Sources of nutrients for high-yielding winter oilseed rape (Brassica napus L.) during post-flowering growth. Agronomy 2020, 10, 626. [Google Scholar] [CrossRef]
- Barraclough, P.B. The growth and activity of winter wheat roots in the field: Nutrient uptakes of high-yielding crops. J. Agric. Sci. Camb. 1986, 106, 45–52. [Google Scholar] [CrossRef]
- Rogers, C.W.; Dari, B.; Hu, G.; Mikkelsen, R. Dry matter production, nutrient accumulation, and nutrient partitioning of barley. J. Plant Nutr. Soil Sci. 2019, 182, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Würschum, T.; Leiser, W.L.; Langner, S.M.; Tucker, M.R.; Longin, C.F.H. Phenotypic and genetic analysis of spike and kernel characteristics in wheat reveals long-term genetic trends of grain yield components. Theor. Appl. Genet. 2018, 131, 2071–2084. [Google Scholar] [CrossRef]
- Ali, I.; Khan, A.A.; Munsif, F.; He, L.; Khan, A.; Ullah, S.; Saeed, W.; Iqbal, A.; Adnan, M.; Ligeng, J. Optimizing rates and application time of potassium fertilizer for improving growth, grain nutrient content and yield of wheat. Open Agric. 2019, 4, 500–508. [Google Scholar] [CrossRef]
- Kang, W.; Fan, M.; Ma, Z.; Shi, X.; Zheng, H. Luxury absorption of potassium by potato plants. Am. J. Potato Res. 2014, 573–578. [Google Scholar] [CrossRef]
- Barłóg, P.; Grzebisz, W.; Pepliński, K.; Szczepaniak, W. Sugar beet response to balanced nitrogen fertilization with phosphorus and potassium. Part I. Dynamics of beet yield development. Bulg. J. Agric. Sci. 2013, 19, 1311–1318. [Google Scholar]
- Szczepaniak, W.; Grzebisz, W.; Potarzycki, J. An assessment of the effect of potassium fertilizing systems on maize nutritional status in critical stages of growth by plant analysis. J. Elem. 2014, 19, 533–548. [Google Scholar] [CrossRef] [Green Version]
- White, P.J. Improving potassium acquisition and utilization by crop plants. Plant Nutr. Soil Sci. 2013, 176, 305–316. [Google Scholar] [CrossRef]
- Pan, Y.; Lu, Z.; Lu, J.; Li, X.; Cong, R. Effects of low sink demand on leaf photosynthesis under potassium deficiency. Plant Physiol. Bioch. 2017, 113, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, W.M.; Beverly, R.B. The dilution effect in plant nutrition studies. Adv. Agron. 1981, 34, 197–224. [Google Scholar]
- Westermann, D.; James, D.; Tindall, T.; Hurst, R. Nitrogen and potassium fertilization of potatoes, sugars and starch. Am. Potato J. 1994, 71, 433–453. [Google Scholar] [CrossRef]
- Grzebisz, W.; Čermák, P.; Rroco, E.; Szczepaniak, W.; Potarzycki, J. Potassium impact on nitrogen use efficiency in potato—A case study from the Central-East Europe. Plant Soil Environ. 2017, 63, 422–427. [Google Scholar]
- Grzebisz, W.; Szczepaniak, W.; Bocianowski, J. Potassium fertilization as a driver of sustainable management of nitrogen in potato (Solanum tuberosum, L.). Field Crops Res. 2020, 254, 107824. [Google Scholar] [CrossRef]
- Withers, P.J.A.; Sylvester-Bradley, R.; Jones, D.L.; Healey, J.R.; Talboys, P.J. Feed the crop not the soil: Rethinking phosphorus management in the food chain. Environ. Sci. Technol. 2014, 48, 6523–6530. [Google Scholar] [CrossRef] [Green Version]
- Ajmeria, I.; Hodgman, T.C.; Lu, C. An integrative systems perspective on plant phosphate research. Genes 2019, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Veneklass, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.-R.; Shane, M.W.; White, P.J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Ros, C.; Bell, R.W.; White, P.P. Effect of seed phosphorus and soil phosphorus applications on early growth of rice (Oryza sativa L.). Soil Sci. Plant Nutr. 1997, 4393, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Shuja, M.N.; Zahoor, M.; Quadri, I. Phytic acid: How have we come. Afr. J. Biotech. 2010, 9, 1551–1554. [Google Scholar]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. P uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, C.; Bucher, M. Molecular mechanisms of phosphate transport in plants. Plant 2002, 216, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach, 2nd ed.; Wiley and Sons: New York, NY, USA, 1995; 432p. [Google Scholar]
- Kvakić, M.; Pellerin, S.; Ciais, P.; Achat, D.L.; Augusto, L.; Deneroy, P.; Gerber, J.S.; Goll, D.; Mollier, A.; Mueller, N.D.; et al. Quantifying the limitation to world cereal production due to soil phosphorus status. Glob. Biogeoch. Cycles 2018, 32, 143–157. [Google Scholar] [CrossRef]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root structure and functioning for efficient acquisition of phosphorus: Matching morphological and physiological traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef] [Green Version]
- Duarah, I.; Deka, M.; Saikia, N.; Deka Boruachh, H.P. Phosphate solubizers enhance NPK fertilizer use efficiency in rice and legume cultivation. Biotech 2011, 1, 227–238. [Google Scholar]
- Liu, D. Root development responses to phosphorus nutrition. J. Integr. Plant Biol. 2021, 1065–1090. [Google Scholar] [CrossRef]
- Penn, C.J.; Camberato, J.J. A critical review on soil chemical processes that control how soil pH affects phosphorus availability to plants. Agriculture 2019, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Krouk, G.; Kiba, T. Nitrogen and phosphorus interactions in plants: From agronomic to physiological and molecular insight. Curr. Opin. Plant Biol. 2020, 57, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.; Simonne, E.; Obreza, T.; Liu, G. Crop response to low phosphorus bioavailability with focus on tomato. Agronomy 2020, 10, 617. [Google Scholar] [CrossRef]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2012, 112, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Potarzycki, J.; Grzebisz, W. Trends in phosphorus concentration in potato organs during the growing season. J. Elem. 2019, 24, 935–952. [Google Scholar] [CrossRef]
- Grant, C.A.; Flaten, D.N.; Tomasiewicz, D.J.; Sheppard, S.C. The importance of early season phosphorus nutrition. Can. J. Plant Sci. 2001, 81, 211–224. [Google Scholar] [CrossRef]
- Szczepaniak, W.; Barłog, P.; Grzebisz, W.; Pepliński, K. Effect of differentiated fertilizing systems on nitrogen accumulation patterns during the growing season—Sugar beet as an example. J. Elem. 2012, 17, 669–688. [Google Scholar] [CrossRef]
- Barłóg, P.; Grzebisz, W.; Szczepaniak, W.; Pepliński, K. Sugar beet response to balanced nitrogen fertilization with phosphorus and potassium. Part II. Dynamics of beet quality. Bulg. J. Agric. Sci. 2013, 20, 1326–1333. [Google Scholar]
- Grzebisz, W.; Szczepaniak, W.; Barłóg, P.; Przygocka-Cyna, K.; Potarzycki, J. Phosphorus sources for winter oilseed rape (Brassica napus L.)—Magnesium sulfate management impact on P use efficiency. Archiv. Agron. Soil Sci. 2018, 12, 1646–1662. [Google Scholar] [CrossRef]
- Zhang, W.; Li, H.; Zhang, J.; Shen, J.; Brown, H.; Wang, E. Contrasting patterns of accumulation, partitioning and remobilization of biomass and phosphorus in a maize cultivar. Crop J. 2022, 10, 254–261. [Google Scholar] [CrossRef]
- Julia, C.; Wissuwa, M.; Kretzschmar, T.; Jeong, K.; Rose, T. Phosphorus uptake, partitioning and redistribution during grain filling in rice. Ann. Bot. 2016, 118, 1151–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Shen, J.; Liao, H. Acquisition or utilization, which is more critical for enhancing phosphorus efficiency in modern crops. Plant Sci. 2010, 179, 302–306. [Google Scholar] [CrossRef]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.-M.; Yvin, J.-C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front. Plant Sci. 2015, 6, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Mazlouzi, M.; Morel, C.; Robert, T.; Chesseron, C.; Salon, C.; Cornu, J.-Y.; Molier, A. The dynamics of Phosphorus uptake and remobilization during the grain development period in durum wheat plants. Plants 2022, 11, 1006. [Google Scholar] [CrossRef]
- Łukowiak, R.; Grzebisz, W.; Sassenrath, G.F. New insights into phosphorus management in agriculture—A crop rotation approach. Sci. Total Environ. 2016, 15, 1062–1077. [Google Scholar] [CrossRef]
- Hu, Z.; Ding, Z.; Al-Yasi, H.M.; Ali, E.F.; Eissa, M.A.; Abou-Elwafa, S.F.; Sayed, M.A.; Said, M.T.; Said, A.A.; Ibrahim, K.A.M.; et al. Modelling of phosphorus nutrition to obtain maximum yield, high P use efficiency and low P-risk for wheat grown in sandy calcareous soils. Agronomy 2021, 11, 1950. [Google Scholar] [CrossRef]
- Bergmann, W. Nutritional Disorders of Plants; Verlag Gustav Fisher: Jena, Germany, 1992; 741p. [Google Scholar]
- Spiertz, J.H.J. Nitrogen, sustainable agriculture and food security. A review. Agron. Sustain. Dev. 2010, 30, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef]
- Weih, M.; Hamnér, K.; Pourazari, F. Analyzing plant nutrient uptake and utilization efficiencies: Comparison between crops and approaches. Plant Soil 2018, 430, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, M.; Darch, T.; Haslam, R. Phosphorus use efficiency and fertilizers: Future opportunities for improvements. Front. Agric. Sci. Eng. 2019, 6, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Evans, L.T.; Fischer, R.A. Yield potential: Its definition, measurement, and significance. Crop Sci. Soc. Am. 1999, 39, 1544–1551. [Google Scholar] [CrossRef]
- Hay, R.; Porter, J. The Physiology of Crop Yield, 2nd ed.; Blackwell Publishing Ltd.: Oxford, UK, 2006; 314p. [Google Scholar]
- Agnus, J.F.; van Herwaarden, A.F. Increasing water use and water use efficiency in dryland wheat. Agron. J. 2001, 93, 290–298. [Google Scholar]
- Passioura, J. Increasing crop productivity when water is scarce—From breeding to field management. Agric. Water Manag. 2006, 80, 176–196. [Google Scholar] [CrossRef] [Green Version]
- Grzebisz, W.; Baer, A.; Barłóg, P.; Szczepaniak, W.; Potarzycki, J. Effect of nitrogen and potassium fertilizing systems on maize grain yield. Fertil. Fertil. 2010, 40, 45–56. [Google Scholar]
- Grzebisz, W.; Barłóg, P.; Szczepaniak, W.; Potarzycki, J. Effect of potassium fertilizing system on dynamics of dry matter accumulation by maize. Fertil. Fertil. 2010, 40, 57–69. [Google Scholar]
- Dobermann, A. Nitrogen use efficiency—State of the art. In Proceedings of the IFA International Workshop on Enhanced Efficiency Fertilizers, Frankfurt, Germany, 28–30 June 2005; pp. 1–16. [Google Scholar]
- Barłóg, P.; Grzebisz, W.; Łukowiak, R. Fertilizers and fertilization strategies mitigating soil factors constraining efficiency of nitrogen in plant production. Plants 2022, 11, 1855. [Google Scholar] [CrossRef]
- Grzebisz, W.; Niewiadomska, A.; Przygocka-Cyna, K. Nitrogen hotspots on the farm—A practice-oriented approach. Agronomy 2022, 12, 1305. [Google Scholar] [CrossRef]
- Rütting, T.; Aronsson, H.; Delin, S. Efficient use of nitrogen in agriculture. Nutr. Cycl. Agroecosys. 2018, 110, 105. [Google Scholar] [CrossRef]
- Hennessy, D.A. On monoculture and the structure of crop rotations. Agric. Econ. 2006, 88, 900–914. [Google Scholar] [CrossRef] [Green Version]
- Pasley, H.; Nichols, V.; Castellano, M.; Baum, M.; Kladivko, E.; Helmers, M.; Archontoulis, S. Rating maize reduces the risk and rate of nitrate leaching. Environ. Res. Lett. 2021, 16, 064063. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; Cadisch, G. Access and excess problems in soil nutrition. Plant Soil 2002, 247, 25–40. [Google Scholar] [CrossRef]
- Grzebisz, W.; Łukowiak, R.; Sassenrath, G. Virtual nitrogen as a tool for assessment of nitrogen at the field scale. Field Crops Res. 2018, 218, 182–184. [Google Scholar] [CrossRef]
- Kautz, T.; Amelung, W.; Ewert, F.; Gaiser, T.; Horn, R.; Jahn, R.; Javaux, M.; Kemna, A.; Kuzyakova, Y.; Munch, J.-C.; et al. Nutrient acquisition from arable subsoils in temperate climates: A review. Soil Biol. Biochem. 2013, 57, 1003–1022. [Google Scholar] [CrossRef]
- Smucker, A.J.M.; Aiken, R.M. Dynamic root responses to water deficits. Soil Sci. 1992, 154, 281–289. [Google Scholar] [CrossRef]
- Bodner, G.; Nakhforoosh, A.; Kaul, H.P. Management of crop water under drought: A review. Agron. Sustain. Dev. 2015, 35, 401–442. [Google Scholar] [CrossRef]
- Grzebisz, W.; Łukowiak, R.; Kotnis, K. Evaluation of nitrogen fertilization systems based on the in-season variability of the nitrogenous growth factors and soil fertility factors—A case of winter oilseed rape (Brassica napus L.). Agronomy 2020, 10, 1701. [Google Scholar] [CrossRef]
- Barłóg, P.; Łukowiak, R.; Grzebisz, W. Predicting the content of soil mineral nitrogen based on the content of calcium chloride-extractable nutrients. J. Plant Nutr. Soil Sci. 2017, 180, 624–635. [Google Scholar] [CrossRef]
- Łukowiak, R.; Barłóg, P.; Grzebisz, W. Soil mineral nitrogen and the rating of CaCl2 extractable nutrients. Plant Soil Environ. 2017, 63, 177–183. [Google Scholar]
- Babulicova, M. The influence of fertilization and crop rotation on the winter wheat production. Plant Soil Environ. 2014, 60, 297–302. [Google Scholar] [CrossRef]
- Torma, S.; Vilček, J.; Lošák, T.; Kužel, S.; Martensson, A. residual plant nutrients in crop residues—An important resource. Acta Agric. Scand. Sec. B Soil Plant Sci. 2017, 68, 358–366. [Google Scholar] [CrossRef]
- Kumar, T.K.; Rana, D.S.; Nain, L. Legume residue and N management for improving productivity and N economy and soil fertility in wheat (Triticum aestivum L.)—Based cropping system. Natl. Acad. Sci. Lett. 2019, 42, 297–307. [Google Scholar] [CrossRef]
- Zhang, W.-P.; Liu, G.-C.; Sun, J.H.; Fornara, D.; Zhang, L.-Z.; Zhang, F.-F.; Li, L. Temporal dynamics of nutrient uptake by neighbouring plant species: Evidence from intercropping. Func. Ecol. 2017, 31, 469–479. [Google Scholar] [CrossRef]
- Macholdt, J.; Piepho, H.P.; Honermeier, B.; Perryman, S.; MacDonald, A.; Poulton, P. The effects of cropping sequence, fertilization and straw management on the yield stability of winter wheat (1986–2017) in the Broadbalk Wheat Experiment, Rothamsted, UK. J. Agric. Sci. 2010, 158, 65–79. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 15th ed.; Pearson Education: Boston, MA, USA, 2017; 1071p. [Google Scholar]
- Usherwood, N.R.; Segars, W.I. Nitrogen interactions with phosphorus and potassium for optimum yield, nitrogen use effectiveness, and environmental stewardship. Sci. World 2001, 1, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, J.; Torres, G.; Driver, E.; Figueiredo, B.; Raun, W.R. World phosphorus use efficiency in cereal crops. Agron. J. 2017, 109, 1670–1677. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Liu, J.; Li, H.; Zou, J.; Xu, H.; Liu, X.; Li, X. Potassium-fertilizer management in winter oilseed-rape production in China. J. Plant Sci. Soil Sci. 2013, 176, 429–440. [Google Scholar] [CrossRef]
- Khan, S.A.; Mulvaney, R.L.; Ellsworth, T.R. The potassium paradox: Implications for soil fertility, crop production and human health. Renew. Agric. Food Syst. 2013, 29, 3–27. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, G.K.; Bennett, E.M.; Potter, P.A.; Ramankutty, N. Agronomic phosphorus imbalances across the world’s croplands. Proc. Natl. Acad. Sci. USA 2011, 108, 3086–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlen, D.L.; Varvel, G.E.; Bullock, D.G.; Cruse, R.M. Crop Rotations for the 21st Century. Adv. Agron. 1994, 53, 1–45. [Google Scholar]
- European Commission. Environmental Impacts of Different Crop Rotation in the European Union; Final Report; European Commission, Bio Intelligence Service S.A.S.: Paris, France, 2010; p. 141. Available online: https://ec.europa.eu/environment/agriculture/pdf/BIO_crop_rotations (accessed on 25 August 2022).
- Tanveer, A.; Ikram, R.M.; Ali, H.H. Crop rotation: Principles and practices. Agron. Crops 2019, 1–12. [Google Scholar] [CrossRef]
- Bruns, H.A. Concepts in crop rotations. In Agricultural Science; Alakpui, G., Ed.; InTech: Rijeka, Croatia, 2012; 26p. [Google Scholar]
- Costa, M.P.; Chadwick, D.; Saget, S.; Rees, R.M.; Williams, M.; Stylres, D. Representing crop rotations in life cycle assessment: A review of legume LCA studies. Inter. J. Life Cycle Assess. 2020, 14, 1942–1956. [Google Scholar] [CrossRef]
- Marini, L.; Stomartin, A.; Vici, G.; Baldoni, G.; Berti, A.; Blecharczyk, A.; Małecka-Jankowiak, I.; Morari, F.; Sawińska, Z.; Bommmaeco, R. Crop rotations sustain cereal yields under a changing climate. Environ. Res. Lett. 2010, 15, 124011. [Google Scholar] [CrossRef]
- Tan, Z.X.; Lal, R.; Wiebe, K.D. Global soil nutrient depletion and yield reduction. J. Sustain. Dev. 2015, 26, 123–146. [Google Scholar] [CrossRef]
- Siebers, N.; Wang, L.; Funk, T.; von Tucher, S.; Merbach, I.; Schweitzer, K.; Kruse, J. Subsoils—A sink for excess fertilizer P but a minor contribution to P plant nutrition: Evidence from long-term fertilization trials. Environ. Sci. Eur. 2021, 33, 60. [Google Scholar] [CrossRef]
- Grzebisz, W. Crop Plant Fertilization. Part II. Fertilizers and Fertilization Systems; PWRiL: Poznań, Poland, 2015; 376p. (In Polish) [Google Scholar]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Monitoring soil for sustainable development and land degradation neutrality. Environ. Monit. Assess. 2018, 190, 57. [Google Scholar] [CrossRef] [Green Version]
- Clarkson, D.T. Nutrient interception and transport by root system. In Physiological Processes Limiting Plant Productivity; Johnson, C.B., Ed.; Butterworths: London, UK, 2013; pp. 307–330. [Google Scholar]
- Johnston, A. Soil Testing Philosophy or “How Do We Make Fertilizer Recommendations”. In Proceedings of the Direct Seeding Conference: Managing Risk for the Future. 2006. Available online: http://www.ssca.ca/conference/conference2006/Johnston.pdf (accessed on 26 August 2022).
- Raynaud, X.; Leadley, P. Soil characteristics play a key role in modeling nutrient competition in plant communities. Ecology 2004, 85, 2200–2214. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop Rotation | Nutrients | |||
|---|---|---|---|---|
| Phosphorus, P2O5 | Potassium K2O | |||
| Demand | Full Recycling | Demand | Full Recycling | |
| Winter oilseed rape, 3.5 t ha−1 | ||||
| -seeds | 70 | - | 35 | - |
| -straw | 40 | 20 ** | 200 | 180 ** |
| Winter wheat, 7.0 t ha−1 | ||||
| -grain | 55 | - | 35 | - |
| -straw | 25 | 12 | 120 | 110 |
| Maize, 8.0 t ha−1 | ||||
| -grain | 60 | - | 40 | - |
| -straw | 30 | 15 | 160 | 145 |
| Spring barley, 5.0 t ha−1 | ||||
| -grain | 45 | - | 25 | - |
| -straw | 10 | 5 | 110 | 100 |
| Total sum | 335 | 53 | 725 | 535 |
| Balance | −282 | −190 | ||
| Total demand | 282 | 190 | ||
| Partial demand, kg ha−1 year−1 | 70.5 | 47.5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzebisz, W.; Diatta, J.; Barłóg, P.; Biber, M.; Potarzycki, J.; Łukowiak, R.; Przygocka-Cyna, K.; Szczepaniak, W. Soil Fertility Clock—Crop Rotation as a Paradigm in Nitrogen Fertilizer Productivity Control. Plants 2022, 11, 2841. https://doi.org/10.3390/plants11212841
Grzebisz W, Diatta J, Barłóg P, Biber M, Potarzycki J, Łukowiak R, Przygocka-Cyna K, Szczepaniak W. Soil Fertility Clock—Crop Rotation as a Paradigm in Nitrogen Fertilizer Productivity Control. Plants. 2022; 11(21):2841. https://doi.org/10.3390/plants11212841
Chicago/Turabian StyleGrzebisz, Witold, Jean Diatta, Przemysław Barłóg, Maria Biber, Jarosław Potarzycki, Remigiusz Łukowiak, Katarzyna Przygocka-Cyna, and Witold Szczepaniak. 2022. "Soil Fertility Clock—Crop Rotation as a Paradigm in Nitrogen Fertilizer Productivity Control" Plants 11, no. 21: 2841. https://doi.org/10.3390/plants11212841
APA StyleGrzebisz, W., Diatta, J., Barłóg, P., Biber, M., Potarzycki, J., Łukowiak, R., Przygocka-Cyna, K., & Szczepaniak, W. (2022). Soil Fertility Clock—Crop Rotation as a Paradigm in Nitrogen Fertilizer Productivity Control. Plants, 11(21), 2841. https://doi.org/10.3390/plants11212841