

Content of Two Major Steroidal Glycoalkaloids in Tomato (Solanum lycopersicum cv. Micro-Tom) Mutant Lines at Different Ripening Stages
 , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
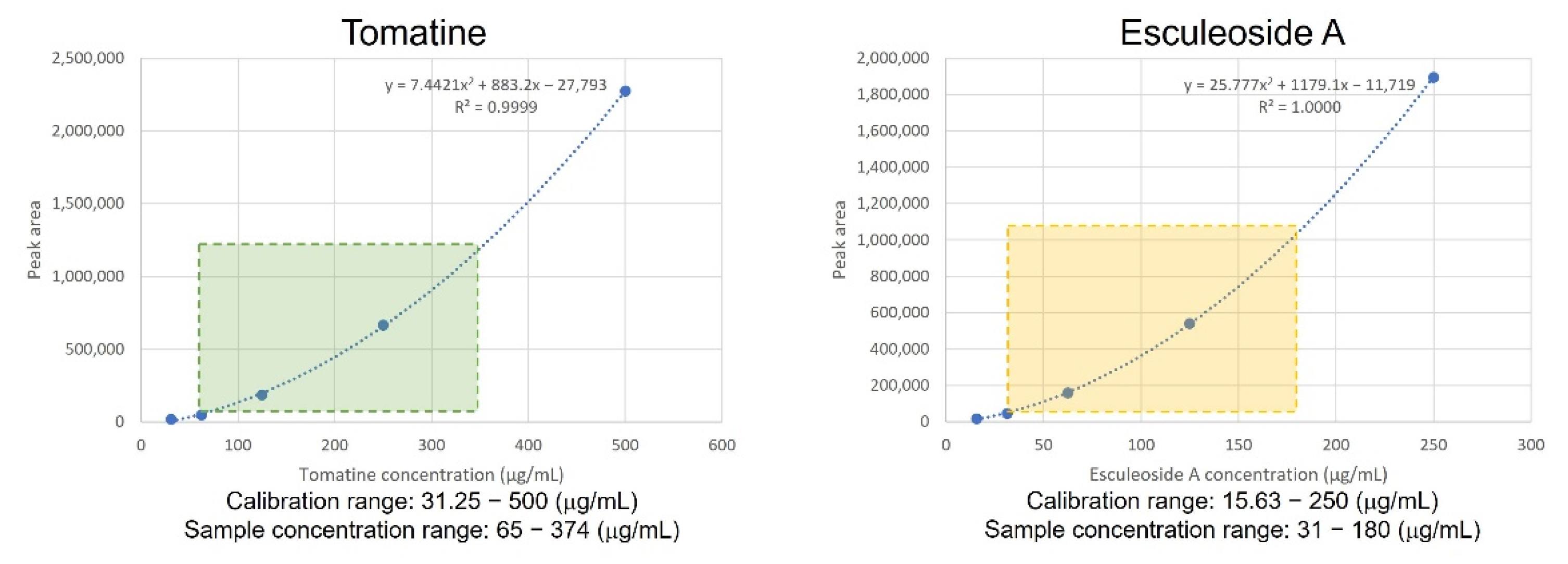
2.1. Quantitative Analysis of Two Steroidal Glycoalkaloids in Micro-Tom Mutant Lines at Two Different Ripening Stages
2.2. Cytotoxic Activities
3. Materials and Methods
3.1. General Procedures
3.2. Plant Materials
3.3. Isolation and Identification of Esculeoside A from the Ripe Red Micro-Tomato
3.4. Preparation of Standard Solutions and Sample Preparation
3.5. Quantitative Analysis
3.6. Cytotoxicity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#home (accessed on 18 September 2022).
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, C.; Ambrosino, P.; Esposito, F.; Fogliano, V. Antioxidant activity and carotenoid and tomatine contents in different typologies of fresh consumption tomatoes. J. Agric. Food Chem. 2000, 48, 4723–4727. [Google Scholar] [CrossRef]
- Yahara, S.; Uda, N.; Nohara, T. Lycoperosides A-C, three stereoisomeric 23-acetoxyspirosolan-3β-ol β-lycotetraosides from Lycopersicon esculentum. Phytochemistry 1996, 42, 169–172. [Google Scholar] [CrossRef]
- Faria-Silva, C.; de Sousa, M.; Carvalheiro, M.C.; Simões, P.; Simões, S. Alpha-tomatine and the two sides of the same coin: An anti-nutritional glycoalkaloid with potential in human health. Food Chem. 2022, 391, 133261. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, Y.; Takai, A.; Uehara, Y.; Ikeda, T.; Okawa, M.; Yamauchi, K.; Ono, M.; Yoshimitsu, H.; Nohara, T. Tomato steroidal alkaloid glycosides, esculeosides A and B, from ripe fruits. Tetrahedron 2004, 60, 4915–4920. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Yahara, S.; Ikeda, T.; Ono, M.; Nohara, T. Cytotoxic major saponin from tomato fruits. Chem. Pharm. Bull. 2003, 51, 234–235. [Google Scholar] [CrossRef] [Green Version]
- Itkin, M.; Rogachev, I.; Alkan, N.; Rosenberg, T.; Malitsky, S.; Masini, L.; Meir, S.; Iijima, Y.; Aoki, K.; de Vos, R.; et al. GLYCOALKALOID METABOLISM1 is required for steroidal alkaloid glycosylation and prevention of phytotoxicity in tomato. Plant Cell 2011, 23, 4507–4525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas, P.D.; Sonawane, P.D.; Heinig, U.; Bocobza, S.E.; Burdman, S.; Aharoni, A. The bitter side of the nightshades: Genomics drives discovery in Solanaceae steroidal alkaloid metabolism. Phytochemistry 2015, 113, 24–32. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; van Kan, J.A.L. Bitter and sweet make tomato hard to (b)eat. New Phytol. 2021, 230, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Nakayasu, M.; Akiyama, R.; Kobayashi, M.; Lee, H.J.; Kawasaki, T.; Watanabe, B.; Urakawa, S.; Kato, J.; Sugimoto, Y.; Iijima, Y.; et al. Identification of α-tomatine 23-hydroxylase involved in the detoxification of a bitter glycoalkaloid. Plant Cell Physiol. 2020, 61, 21–28. [Google Scholar] [CrossRef]
- Akiyama, R.; Nakayasu, M.; Umemoto, N.; Kato, J.; Kobayashi, M.; Lee, H.J.; Sugimoto, Y.; Iijima, Y.; Saito, K.; Muranaka, T.; et al. Tomato E8 encodes a C-27 hydroxylase in metabolic detoxification of α-tomatine during fruit ripening. Plant Cell Physiol. 2021, 62, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Nakamura, Y.; Ogata, Y.; Tanaka, K.; Sakurai, N.; Suda, K.; Suzuki, T.; Suzuki, H.; Okazaki, K.; Kitayama, M.; et al. Metabolite annotations based on the integration of mass spectral information. Plant J. 2008, 54, 949–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iijima, Y.; Fujiwara, Y.; Tokita, T.; Ikeda, T.; Nohara, T.; Aoki, K.; Shibata, D. Involvement of ethylene in the accumulation of esculeoside A during fruit ripening of tomato (Solanum lycopersicum). J. Agric. Food Chem. 2009, 57, 3247–3252. [Google Scholar] [CrossRef]
- Yamanaka, T.; Vincken, J.P.; Zuilhof, H.; Legger, A.; Takada, N.; Gruppen, H. C22 isomerization in alpha-tomatine-to-esculeoside A conversion during tomato ripening is driven by C27 hydroxylation of triterpenoidal skeleton. J. Agric. Food Chem. 2009, 57, 3786–3791. [Google Scholar] [CrossRef]
- Jayakumar, K.; Murugan, K. Solanum alkaloids and their pharmaceutical roles: A review. J. Anal. Pharm. Res. 2016, 3, 00075. [Google Scholar]
- Morris, S.C.; Lee, T.H. The toxicity and teratogenicity of Solanaceae glycoalkaloids, particularly those of the potato (Solanum tuberosum): A review. Food Tech Aust. 1984, 36, 118–124. [Google Scholar]
- Wu, H.; Li, W.; Wang, T.; Rong, Y.; He, Z.; Huang, S.; Zhang, L.; Wu, Z.; Liu, C. α-Tomatine, a novel early-stage autophagy inhibitor, inhibits autophagy to enhance apoptosis via Beclin-1 in Skov3 cells. Fitoterapia 2021, 152, 104911. [Google Scholar] [CrossRef]
- Yelken, B.Ö.; Balcı, T.; Süslüer, S.Y.; Kayabaşı, Ç.; Avcı, Ç.B.; Kırmızıbayrak, P.B.; Gündüz, C. The effect of tomatine on metastasis related matrix metalloproteinase (MMP) activities in breast cancer cell model. Gene 2017, 627, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Morrow, W.J.W.; Yang, Y.-W.; Sheikh, N.A. Immunobiology of the tomatine adjuvant. Vaccine 2004, 22, 2380–2384. [Google Scholar] [CrossRef] [PubMed]
- Chiu, F.L.; Lin, J.K. Tomatidine inhibits iNOS and COX-2 through suppression of NF-κB and JNK pathways in LPS-stimulated mouse macrophages. FEBS Lett. 2008, 582, 2407–2412. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Tomato glycoalkaloids: Role in the plant and in the diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef] [PubMed]
- Nohara, T.; Ono, M.; Ikeda, T.; Fujiwara, Y.; El-Aasr, M. The tomato saponin, esculeoside A. J. Nat. Prod. 2010, 73, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-R.; Kanda, Y.; Tanaka, A.; Manabe, H.; Nohara, T.; Yokomizo, K. Anti-hyaluronidase activity in vitro and amelioration of mouse experimental dermatitis by tomato saponin, esculeoside A. J. Agric. Food. Chem. 2016, 64, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-R.; Kimura, S.; Nohara, T.; Yokomizo, K. Competitive inhibition of mammalian hyaluronidase by tomato saponin, esculeoside A. Nat. Prod. Comm. 2018, 13, 1461–1463. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, Y.; Katayanagi, Y.; Kamiya, M.; Yamamoto, Y.; Fukutomi, R.; Imai, S.; Miyoshi, N.; Ohashi, N. Tomato saponin supplementation ameliorates the development of experimental arthritis by regulating inflammatory responses. J. Func. Foods 2018, 49, 458–464. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, L.; Liu, J.; Lu, F.; Wang, L.; Chen, Y.; Li, D. Hypoglycemic effects of esculeoside A are mediated via activation of AMPK and upregulation of IRS-1. BMC Complement. Altern. Med. 2019, 19, 136. [Google Scholar] [CrossRef] [Green Version]
- Rick, C.M.; Uhlig, J.W.; Jones, A.D. High α-tomatine content in ripe fruit of Andean Lycopersicon esculentum var. cerasiforme: Developmental and genetic aspects. Proc. Natl. Acad. Sci. USA 1994, 91, 12877–12881. [Google Scholar] [CrossRef] [Green Version]
- Steinert, K.; Hövelmann, Y.; Hübner, F.; Humpf, H.-U. Identification of novel iso-esculeoside B from tomato fruits and LC−MS/MS-based food screening for major dietary steroidal alkaloids focused on esculeosides. J. Agric. Food Chem. 2020, 68, 14492–14501. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Anticarcinogenic, cardioprotective, and other health benefits of tomato compounds lycopene, α-tomatine, and tomatidine in pure form and in fresh and processed tomatoes. J. Agric. Food Chem. 2013, 61, 9534–9550. [Google Scholar] [CrossRef]
- Friedman, M.; Levin, C.E.; Lee, S.U.; Kim, H.J.; Lee, I.S.; Byun, J.O.; Kozukue, N. Tomatine-containing green tomato extracts inhibit growth of human breast, colon, liver, and stomach cancer cells. J. Agric. Food Chem. 2009, 57, 5727–5733. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Prim. 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal violet assay for determining viability of cultured cells. Cold Spring Harb. Protoc. 2016, 4, pdb-prot087379. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mutant Lines | Ripening Stage | Tomatine | Esculeoside A |
|---|---|---|---|
| 14-3 | immature green | 961.4 ± 5.9 | nd |
| mature red | nd | 296.4 ± 3.2 | |
| 14-5 | immature green | 502.9 ± 1.7 | nd |
| mature red | nd | 356.1 ± 3.7 | |
| 15-4 | immature green | 660.0 ± 7.8 | nd |
| mature red | nd | 356.4 ± 2.0 | |
| 18-7 | immature green | 508.9 ± 3.3 | nd |
| mature red | nd | 348.9 ± 3.9 | |
| 19-2 | immature green | 782.0 ± 9.0 | nd |
| mature red | nd | 298.8 ± 3.3 | |
| 20-5 | immature green | 452.8 ± 2.7 | nd |
| mature red | nd 2 | 414.5 ± 7.9 | |
| 22-6 | immature green | 428.8 ± 6.2 | nd |
| mature red | nd | 495.7 ± 0.3 | |
| 26-2 | immature green | 346.1 ± 2.6 | nd |
| mature red | nd | 204.0 ± 1.8 | |
| 41-1 | immature green | 575.5 ± 8.1 | nd |
| mature red | nd 2 | 231.7 ± 2.0 | |
| 44-5 | immature green | 299.1 ± 3.7 1 | nd 2 |
| mature red | nd | 444.0 ± 3.2 | |
| Original cultivar | immature green | 1782.9 ± 7.2 | nd |
| mature red | nd | 399.8 ± 1.3 |
| Mutant Lines | Ripening Stage | Tomatine | Esculeoside A |
|---|---|---|---|
| 13-8 | immature green | 1494.6 ± 17.6 | nd |
| mature red | nd | 698.9 ± 4.6 | |
| 15-8 | immature green | 1430.8 ± 10.1 | nd |
| mature red | nd | 597.5 ± 4.6 | |
| 20-7 | immature green | 1537.3 ± 15.9 | nd |
| mature red | nd | 366.7 ± 1.4 | |
| 26-8 | immature green | 1299.2 ± 11.0 | nd |
| mature red | nd | 377.6 ± 2.7 | |
| 27-1 | immature green | 1198.0 ± 4.0 | nd |
| mature red | nd | 281.0 ± 4.0 | |
| 27-7 | immature green | 1266.6 ± 13.3 | nd |
| mature red | nd | 546.3 ± 4.6 | |
| 37-4 | immature green | 1700.8 ± 14.3 | nd |
| mature red | 510.2 ± 5.5 | 428.6 ± 2.3 | |
| 42-3 | immature green | 1894.8 ± 7.9 1 | nd 2 |
| mature red | nd | 281.7 ± 2.5 | |
| 42-5 | immature green | 1392.4 ± 8.5 | nd |
| mature red | nd | 560.4 ± 8.9 | |
| 43-2 | immature green | 1702.8 ± 4.0 1 | nd |
| mature red | nd | 283.6 ± 4.0 | |
| Original cultivar | immature green | 1782.9 ± 7.2 | nd |
| mature red | nd | 399.8 ± 1.3 |
| Mutant Lines | Ripening Stage | Tomatine | Esculeoside A |
|---|---|---|---|
| 13-6 | immature green | 1429.4 ± 7.4 | nd |
| mature red | nd | 786.0 ± 4.2 | |
| 13-8 | immature green | 1494.6 ± 17.6 1 | nd 2 |
| mature red | nd | 698.9 ± 4.6 | |
| 16-7 | immature green | 1044.5 ± 10.8 | nd |
| mature red | nd | 555.6 ± 3.2 | |
| 17-3 | immature green | 1153.2 ± 14.8 | nd |
| mature red | nd | 447.5 ± 4.8 | |
| 17-8 | immature green | 1156.6 ± 8.6 | nd |
| mature red | nd | 676.3 ± 1.8 | |
| 18-2 | immature green | 1178.6 ± 13.1 | nd |
| mature red | n | 479.7 ± 3.1 | |
| 22-1 | immature green | 900.1 ± 15.8 | nd |
| mature red | nd | 556.1 ± 2.9 | |
| 22-6 | immature green | 428.8 ± 6.2 | nd |
| mature red | nd | 495.7 ± 0.3 | |
| 23-1 | immature green | 947.7 ± 5.4 | nd |
| mature red | nd | 697.6 ± 3.7 | |
| 25-2 | immature green | 993.2 ± 11.0 | nd |
| mature red | nd | 620.4 ± 1.3 | |
| Original cultivar | immature green | 1782.9 ± 7.2 | nd |
| mature red | nd | 399.8 ± 1.3 |
| Mutant Lines | Ripening Stage | Tomatine | Esculeoside A |
|---|---|---|---|
| 13-2 | immature green | 1103.5 ± 8.2 1 | nd 2 |
| mature red | 413.8 ± 3.6 | 371.4 ± 4.0 | |
| 13-5 | immature green | 975.7 ± 6.7 | nd |
| mature red | 510.1 ± 6.2 | 550.1 ± 1.9 | |
| 17-7 | immature green | 946.5 ± 9.7 | nd |
| mature red | 335.3 ± 2.8 | 433.2 ± 1.6 | |
| 22-4 | immature green | 817.7 ± 20.6 | nd |
| mature red | 236.9 ± 7.9 | 413.9 ± 6.7 | |
| 23-3 | immature green | 855.8 ± 6.9 | nd |
| mature red | 555.5 ± 8.2 | 317.5 ± 7.0 | |
| 24-5 | immature green | 983.2 ± 17.6 | nd |
| mature red | 344.2 ± 2.5 | 317.7 ± 4.1 | |
| 24-7 | immature green | 1093.2 ± 6.4 | nd |
| mature red | 419.4 ± 4.2 | 549.7 ± 2.5 | |
| 26-1 | immature green | 971.5 ± 10.0 | nd |
| mature red | 259.8 ± 4.1 | 392.1 ± 3.9 | |
| 26-5 | immature green | 1025.6 ± 15.4 | nd |
| mature red | 207.7 ± 1.3 | 499.6 ± 4.1 | |
| 27-4 | immature green | 1072.1 ± 24.6 | nd |
| mature red | 278.0 ± 4.2 | 512.0 ± 4.0 | |
| 37-4 | immature green | 1700.8 ± 14.3 | nd |
| mature red | 510.2 ± 5.5 | 428.6 ± 2.3 | |
| 39-5 | immature green | 1173.3 ± 14.6 | nd |
| mature red | 322.3 ± 0.2 | 360.5 ± 3.1 | |
| 40-8 | immature green | 909.5 ± 20.6 | nd |
| mature red | 332.9 ± 1.2 | 417.2 ± 3.3 | |
| 41-2 | immature green | 1274.6 ± 22.5 | nd |
| mature red | 622.8 ± 4.3 | 409.7 ± 1.0 | |
| 45-2 | immature green | 781.5 ± 11.1 | nd |
| mature red | 258.0 ± 1.0 | 300.3 ± 2.6 | |
| Original cultivar | immature green | 1782.9 ± 7.2 | nd |
| mature red | nd | 399.8 ± 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngo, T.H.; Park, J.; Jo, Y.D.; Jin, C.H.; Jung, C.-H.; Nam, B.; Han, A.-R.; Nam, J.-W. Content of Two Major Steroidal Glycoalkaloids in Tomato (Solanum lycopersicum cv. Micro-Tom) Mutant Lines at Different Ripening Stages. Plants 2022, 11, 2895. https://doi.org/10.3390/plants11212895
Ngo TH, Park J, Jo YD, Jin CH, Jung C-H, Nam B, Han A-R, Nam J-W. Content of Two Major Steroidal Glycoalkaloids in Tomato (Solanum lycopersicum cv. Micro-Tom) Mutant Lines at Different Ripening Stages. Plants. 2022; 11(21):2895. https://doi.org/10.3390/plants11212895
Chicago/Turabian StyleNgo, Trung Huy, Jisu Park, Yeong Deuk Jo, Chang Hyun Jin, Chan-Hun Jung, Bomi Nam, Ah-Reum Han, and Joo-Won Nam. 2022. "Content of Two Major Steroidal Glycoalkaloids in Tomato (Solanum lycopersicum cv. Micro-Tom) Mutant Lines at Different Ripening Stages" Plants 11, no. 21: 2895. https://doi.org/10.3390/plants11212895
APA StyleNgo, T. H., Park, J., Jo, Y. D., Jin, C. H., Jung, C. -H., Nam, B., Han, A. -R., & Nam, J. -W. (2022). Content of Two Major Steroidal Glycoalkaloids in Tomato (Solanum lycopersicum cv. Micro-Tom) Mutant Lines at Different Ripening Stages. Plants, 11(21), 2895. https://doi.org/10.3390/plants11212895