
Utilization of Novel Perilla SSR Markers to Assess the Genetic Diversity of Native Perilla Germplasm Accessions Collected from South Korea


Abstract
:1. Introduction
2. Results
2.1. Polymorphic Test for New Developed PSPSs
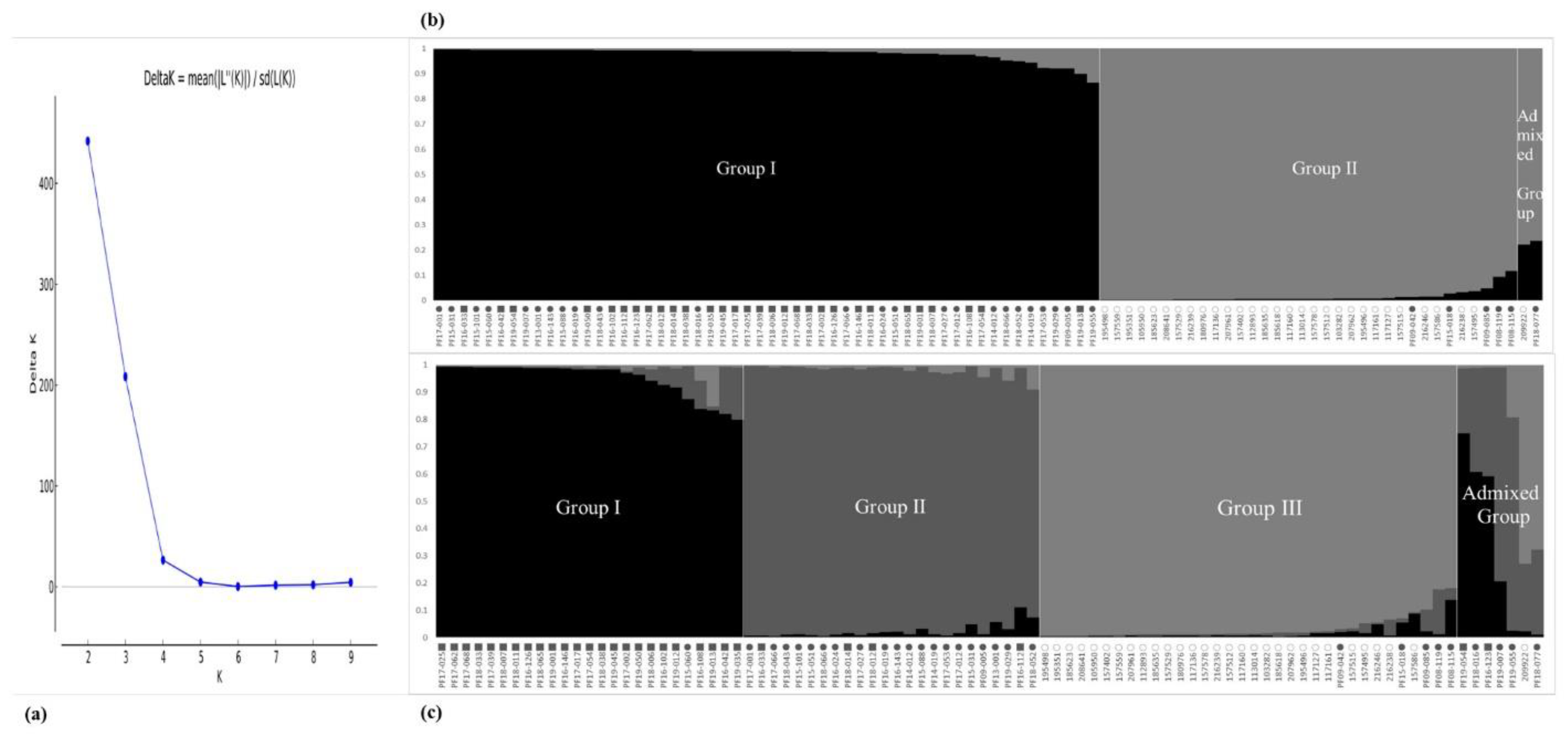
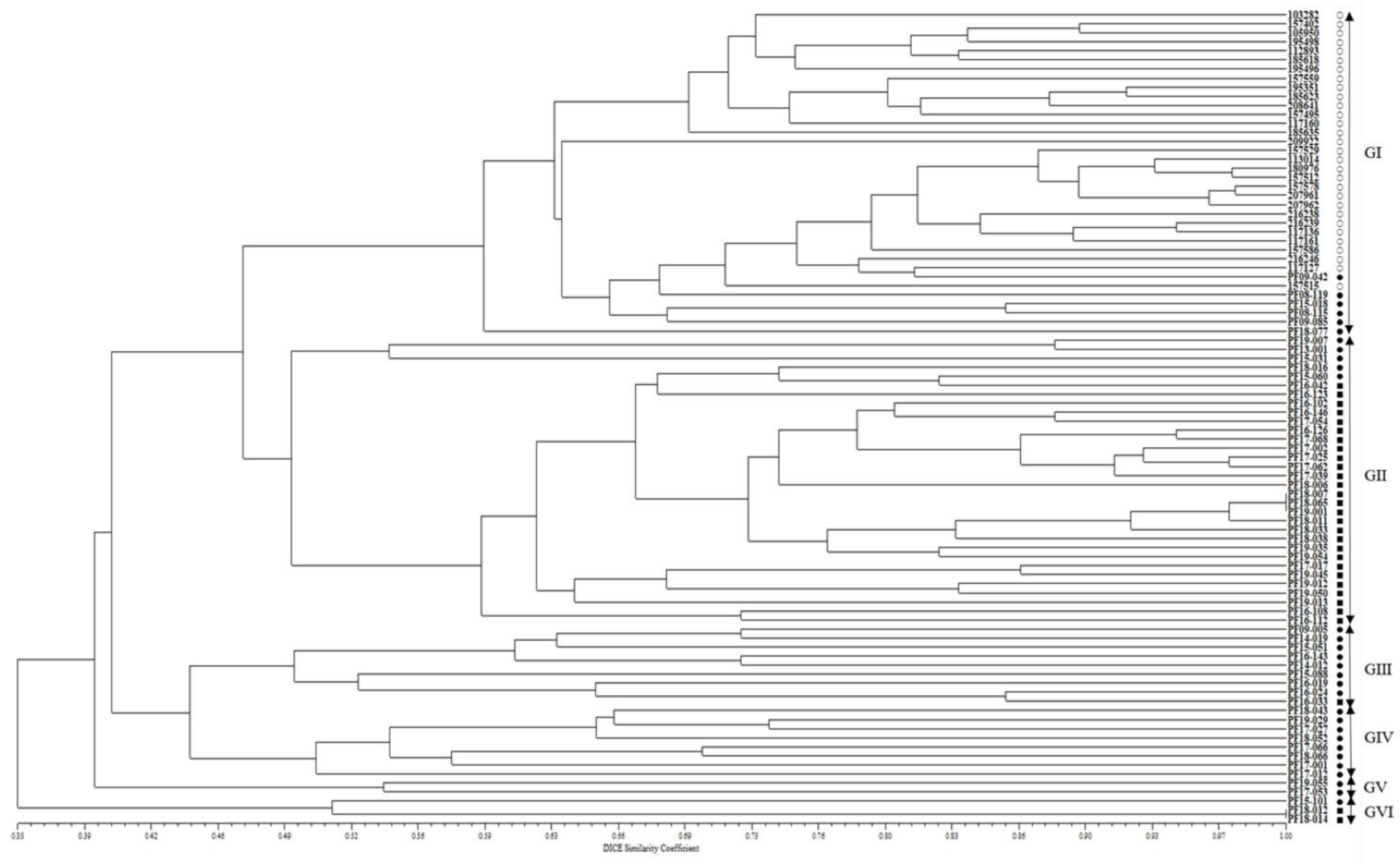
2.2. Population Structure and Phylogenetic Relationship among Accessions of the CWTPC Collected from South Korea
2.3. Seed Characteristics and Association Mapping Analysis of 90 Accessions of the CWTPC Using Novel PSPSs
3. Discussion
3.1. Development of Novel SSR Markers in Perilla Crop and Their Use for Genetic Variation Analysis
3.2. Analysis of Genetic Diversity, Phylogenetic Relationships and Association Mapping Analysis of the CWTPC Collected in South Korea
4. Materials and methods
4.1. Plant Materials and DNA Extraction
4.2. Development of PSPSs and SSR Amplification
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, J.K.; Ohnishi, O. Geographical differentiation of morphological characters among Perilla crops and their weedy types in East Asia. Breed. Sci. 2001, 51, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Ohnishi, O. Genetic relationships among cultivated types of Perilla frutescens and their weedy types in East Asia revealed by AFLP markers. Genet. Resour. Crop Evol. 2003, 50, 65–74. [Google Scholar] [CrossRef]
- Nitta, M.; Lee, J.K.; Ohnishi, O. Asian Perilla crop and their weedy forms: Their cultivation, utilization and genetic relationships. Econ. Bot. 2003, 57, 245–253. [Google Scholar] [CrossRef]
- Nitta, M.; Lee, J.K.; Kang, C.W.; Katsuta, M.; Yasumoto, S.; Liu, D.; Nagamine, T.; Ohnishi, O. The distribution of Perilla species. Genet. Resour. Crop Evol. 2005, 52, 797–804. [Google Scholar] [CrossRef]
- Sa, K.J.; Kim, D.M.; Oh, J.S.; Park, H.; Hyun, D.Y.; Lee, S.; Rhee, J.H.; Lee, J.K. Construction of a core collection of native Perilla germplasm collected from South Korea based on SSR markers and morphological characteristics. Sci. Rep. 2021, 11, 1–13. [Google Scholar]
- Lee, J.K.; Nitta, M.; Kim, N.S.; Park, C.H.; Yoon, K.M.; Shin, Y.B.; Ohnishi, O. Genetic diversity of Perilla and related weedy types in Korea determined by AFLP analyses. Crop Sci. 2002, 42, 2161–2166. [Google Scholar] [CrossRef]
- Sa, K.J.; Choi, S.H.; Ueno, M.; Park, K.C.; Park, Y.J.; Ma, K.H.; Lee, J.K. Identification of genetic variations of cultivated and weedy types of Perilla species in Korea and Japan using morphological and SSR markers. Genes Genom. 2013, 35, 649–659. [Google Scholar] [CrossRef]
- Li, H.L. The vegetables of ancient China. Econ. Bot. 1969, 23, 235–260. [Google Scholar] [CrossRef]
- Makino, T. Makino’s New Illustrated Flora of Japan; Hokuryukan Co.: Tokyo, Japan, 1961. (In Japanese) [Google Scholar]
- Nitta, M. Origin Perilla Crops and Their Weedy Type. Ph.D. Thesis, Kyoto University, Kyoto, Japan, 2001; p. 78. [Google Scholar]
- Ma, S.J.; Sa, K.J.; Hong, T.K.; Lee, J.K. Genetic diversity and population structure analysis in Perilla crop and their weedy types from northern and southern areas of China based on simple sequence repeat (SSRs). Genes Genom. 2019, 41, 267–281. [Google Scholar] [CrossRef]
- Nitta, M.; Ohnishi, O. Genetic relationships among two Perilla crop, shiso and egoma, and the weedy type revealed by RAPD markers. Genes Genet. Syst. 1999, 74, 43–48. [Google Scholar] [CrossRef]
- Yamane, Y. Cytogenetic studies in Perilla and Coleus. I. Chromosome numbers. Jpn. J. Genet. 1950, 25, 220. (In Japanese) [Google Scholar]
- Nagai, I. On “Shiso” and “Egoma”. Agric. Hortic. 1935, 10, 2265–2273. (In Japanese) [Google Scholar]
- Honda, G.; Koezuka, Y.; Tabata, M. Genetic studies of fruit color and hardness in Perilla frutescens. Jpn. J. Breed. 1990, 40, 469–474. [Google Scholar] [CrossRef]
- Honda, G.; Yuba, A.; Kojima, T.; Tabata, M. Chemotaxonomic and cytogenetic studies on Perilla frutescens var. citiodora (‘Lemon Egoma’). Nat. Med. 1994, 48, 185–190. [Google Scholar]
- Lim, S.E.; Sa, K.J.; Ha, Y.J.; Lee, J.K. Genetic analysis of F2 population derived from the cross between Perilla frutescens var. frutescens and var. crispa. Korean J. Breed. Sci. 2019, 51, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.E.; Sa, K.J.; Lee, J.K. Bulk segregant analysis identifies SSR markers associated with leaf-and seed related traits in Perilla crop (Perilla frutescens L.). Genes Genom. 2021, 43, 323–332. [Google Scholar] [CrossRef]
- Kim, J.Y.; Sa, K.J.; Ha, Y.J.; Lee, J.K. Genetic variation and association mapping in F2 population of Perilla crop (Perilla frutescens L.) using new developed Perilla SSR markers. Euphytica 2021, 217, 135. [Google Scholar] [CrossRef]
- Park, Y.J.; Lee, J.K.; Kim, N.S. Simple sequence repeat polymorphisms (SSRPs) for evaluation of molecular diversity and germplasm classification of minor crops. Molecules 2009, 14, 4546–4569. [Google Scholar] [CrossRef] [Green Version]
- Sa, K.J.; Choi, I.K.; Park, K.C.; Lee, J.K. Genetic diversity and population structure among accessions of Perilla frutescens (L.) Britton in East Asia using new developed microsatellite markers. Genes Genom. 2018, 40, 1319–1329. [Google Scholar] [CrossRef]
- Ha, Y.J.; Sa, K.J.; Lee, J.K. Identifying SSR markers associated with seed characteristics in Perilla (Perilla frutescens L.). Physiol. Mol. Biol. Plants 2021, 27, 93–105. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lee, J.K.; Kim, N.S.; Yu, J.W.; Dixit, A.; Cho, E.G.; Park, Y.J. Isolation and characterization of microsatellite markers in Perilla frutescens Britt. Mol. Ecol. Notes. 2005, 5, 455–457. [Google Scholar] [CrossRef]
- Park, Y.J.; Dixit, A.; Ma, K.H.; Lee, J.K.; Lee, M.H.; Chung, C.S.; Nitta, M.; Okuno, K.; Kim, T.S.; Cho, E.G.; et al. Evaluation of genetic diversity and relationships within an on-farm collection of Perilla frutescens (L.) Britt. using SSR markers. Genet. Resour. Crop Evol. 2008, 55, 523–535. [Google Scholar] [CrossRef]
- Sa, K.J.; Lim, S.E.; Choi, I.K.; Park, K.C.; Lee, J.K. Development and characterization of new microsatellite markers for Perilla frutescens (L.) Britton. Am. J. Plant. Sci. 2019, 10, 1623–1630. [Google Scholar] [CrossRef]
- Oh, J.K.; Sa, K.J.; Hyun, D.Y.; Cho, G.T.; Lee, J.K. Assessment of genetic diversity and population structure among a collection of Korean Perilla germplasms based on SSR markers. Genes Genom. 2020, 42, 1419–1430. [Google Scholar] [CrossRef]
- Oh, J.S.; Sa, K.J.; Park, H.; Hyun, D.Y.; Lee, S.; Rhee, J.H.; Lee, J.K. Genetic variation of native Perilla germplasms collected from south korea using simple sequence repeat (SSR) markers and morphological characteristics. Plants 2021, 10, 1764. [Google Scholar] [CrossRef]
- Park, D.H.; Sa, K.J.; Lim, S.E.; Ma, S.J.; Lee, J.K. Genetic diversity and population structure of Perilla frutescens collected from Korea and China based on simple sequence repeats (SSRs). Genes Genom. 2019, 41, 1329–1340. [Google Scholar] [CrossRef]
- Mutz, K.O.; Heilkenbrinker, A.; Lonne, M.; Walter, J.G.; Stahl, F. Transcriptome analysis using next-generation sequencing. Curr. Opin. Biotech. 2013, 24, 22–30. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Z.; Chen, J.; Liu, C.; Zhu, W.; Wang, L.; Meng, L. De Novo transcriptome assembly and development of novel microsatellite markers for the traditional Chinese medicinal herb, Veratrilla baillonii Franch (Gentianaceae). Evol. Bioinform. 2015, 11, 39–45. [Google Scholar]
- Fukushima, A.; Nakamura, M.; Suzuki, H.; Saito, K.; Yamazaki, M. High-throughput sequencing and de novo assembly of red and green forms of the Perilla frutescens var. crispa transcriptome. PLoS ONE 2015, 10, e0129154. [Google Scholar] [CrossRef] [Green Version]
- Tong, W.; Kwon, S.J.; Lee, J.; Choi, I.K.; Park, Y.J.; Choi, S.H.; Sa, K.J.; Kim, B.W.; Lee, J.K. Gene set by de novo assembly of Perilla species and expression profiling between P. frutescens (L.) var. frutescens and var. crispa. Gene 2015, 559, 155–163. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Sa, K.J.; Choi, S.H.; Ueno, M.; Lee, J.K. Genetic diversity and population structure in cultivated and weedy types of Perilla in East Asia and other countries as revealed by SSR markers. Hortic. Environ. Biote. 2015, 56, 524–534. [Google Scholar] [CrossRef]
- Ma, S.J.; Sa, K.J.; Hong, T.K.; Lee, J.K. Genetic diversity and population structure analysis in Perilla frutescens from Northern areas of China based on simple sequence repeats. Genet. Mol. Res. 2017, 16, gmr16039746. [Google Scholar] [CrossRef]
- Park, H.; Sa, K.J.; Hyun, D.Y.; Lee, S.; Lee, J.K. Identifying ssr markers related to seed fatty acid content in Perilla crop (Perilla frutescens L.). Plants 2021, 10, 1404. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, L.; Qian, E.; Long, F.; Cui, W.; Ji, A.; Zhang, F.; Zou, K.; Huang, J.; Li, C. Genetic polymorphisms and haplotypic structure analysis of the Guizhou Gelao ethnic group based on 35 Y-STR loci. Legal. Med. 2020, 43, 101666. [Google Scholar] [CrossRef]
- Zhu, B.F.; Zhang, Y.D.; Liu, W.J.; Meng, H.T.; Yuan, G.L.; Lv, Z.; Dong, N.; Li, Q.; Yang, C.H.; Zhang, Y.H.; et al. Genetic diversity and haplotype structure of 24 Y-chromosomal STR in Chinese Hui ethnic group and its genetic relationships with other populations. Electrophoresis 2014, 35, 1993–2000. [Google Scholar] [CrossRef]
- Hancock, J.F. Plant Evolution and the Origin of Crop Species; Prentice Hall: Englewood Cliffs, NJ, USA, 1992. [Google Scholar]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The molecular genetics of crop domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [Green Version]
- Gross, B.L.; Olsen, K.M. Genetic perspectives on crop domestication. Trends Plant Sci. 2010, 15, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohindji, F.S.; Sogbohossou, D.E.O.; Zohoungbogbo, H.P.F.; Houdegbe, C.A.; Achigan-Dako, E.G. Understanding molecular mechanisms of seed dormancy for improved germination in traditional leafy vegetables: An overview. Agronomy 2020, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, S. Origin and Dispersal of Common Millet and Foxtail Millet. JARQ 1987, 21, 84–89. [Google Scholar]
- Kim, K.S.; Kim, S.H.; Kim, J.; Tripathi, P.; Lee, J.D.; Chung, Y.S.; Kim, Y. A Large Root Phenome Dataset Wide-Opened the Potential for Underground Breeding in Soybean. Front. Plant Sci. 2021, 12, 704239. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Ladizinsky, G. Plant Evolution under Domestication; Chapman & Hall Published: Raleigh, NC, USA, 1998. [Google Scholar]
- Lee, J.K.; Kim, N.S. Genetic diversity and relationships of cultivated and weedy types of Perilla frutescens collected from East Asia revealed by microsatellite markers. Korean J. Breed. Sci. 2007, 39, 491–499. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small amounts of leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System version 2.0 User Guide; Exter Software: Setauket, NY, USA, 1998. [Google Scholar]
- Pritchard, J.K.; Wen, W.; Falush, D. Documentation for Structure Software: Version 2.3. 2003. Available online: http://pritc.h.bsd.uchicago.edu/structure.html (accessed on 1 December 2021).
- Bradbury, P.; Zhang, Z.; Kroon, D.; Casstevens, T.; Ramdoss, Y.; Buckler, E. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| SSR Loci | Forward Primer (5’–3’) | Reverse Primer (3’–5’) | Repeat Motif | Ta | Allele Size (bp) | Allele No | MAF | GD | PIC |
|---|---|---|---|---|---|---|---|---|---|
| KNUPF127 | CTGAGCAGAATGGGATAAAATC | CATGAATCCAAACCTGAGAAAT | (AT)8 | 61 | 210–240 | 5 | 0.778 | 0.378 | 0.356 |
| KNUPF128 | TTTTCTGGAAAGAAAAACCAAA | GTCATTTTCCAAACCGTAAAAA | (AT)8 | 60 | 160–180 | 9 | 0.544 | 0.597 | 0.532 |
| KNUPF129 | AATACATGAACACTGTCACACCA | AGATCATGTTAGCAGGCAATTT | (AT)8 | 64 | 155–180 | 14 | 0.344 | 0.831 | 0.817 |
| KNUPF130 | TGAGAAATCTAACCCCAAACTT | CCTGTTTTTGATCTCTTACTTGC | (CA)8 | 62 | 200–220 | 2 | 0.778 | 0.346 | 0.286 |
| KNUPF131 | TGGATCAAACATTGTAACAGGA | ACCAACACCAAAACTACTGACC | (CA)8 | 63 | 190–220 | 3 | 0.778 | 0.370 | 0.340 |
| KNUPF132 | TTTGAGATAGCTCGGTTCAAAT | CTTCAGGAGCCACATATTCTTC | (AG)16 | 62 | 190–230 | 9 | 0.478 | 0.709 | 0.678 |
| KNUPF133 | TTAAAAGATTGCATGTCTGCAC | CCTTTTCCTGTGTTTTCTCAAG | (AG)16 | 62 | 220–260 | 3 | 0.456 | 0.592 | 0.505 |
| KNUPF134 | TATAATACACGAAGACGCCACA | TTTTGTCCTGTCAACTTCCTCT | (AG)15 | 64 | 130–150 | 6 | 0.367 | 0.749 | 0.710 |
| KNUPF135 | AATAGGTCGACTATGTTCGTGG | ATCAAATCTGCCAATCTCATTT | (CT)12 | 62 | 135–175 | 6 | 0.811 | 0.332 | 0.318 |
| KNUPF136 | TCAAGCAGAGATTGATTCAGTG | CAAAGAATAATCACCACACCAA | (AG)12 | 62 | 140–170 | 6 | 0.589 | 0.589 | 0.544 |
| KNUPF137 | AATCAAGGTGTGCAATCATACA | GGTGTTCACTAGAGTCTCGGTC | (CT)11 | 64 | 300–315 | 3 | 0.911 | 0.164 | 0.153 |
| KNUPF138 | CTGCGTGTGCTGATAAAACTC | TTCTGCTGCTGTATTCTGAGTG | (AG)11 | 64 | 150–180 | 5 | 0.556 | 0.607 | 0.554 |
| KNUPF139 | CCCTAAATCAAACTTGAATCCC | GGGTCGCTAGTAAAGAAGGTTT | (CT)10 | 62 | 170–190 | 6 | 0.567 | 0.598 | 0.545 |
| KNUPF140 | GGGTTCTTTCTTTCTCCCTTTA | AGCTAAGCTGGCTTCTCTATTTT | (CT)10 | 63 | 200–220 | 5 | 0.689 | 0.448 | 0.374 |
| KNUPF141 | ATCTTTCGCAATATGTTTCCTG | AAGTTCACAAAGTTGAACGCTT | (CT)10 | 63 | 180–190 | 4 | 0.656 | 0.515 | 0.466 |
| KNUPF142 | ATCTCGCATTCTTTTAGCTACG | TTTCTCGGAAAATCACTCTGTT | (CT)10 | 62 | 200–230 | 6 | 0.411 | 0.713 | 0.665 |
| KNUPF143 | GGATCTTCTGGGATTTCTTACC | GCCGTATGTCGTCCTTGAT | (CT)10 | 63 | 165–190 | 8 | 0.267 | 0.800 | 0.770 |
| Average | 5.9 | 0.587 | 0.549 | 0.507 |
| SSR Loci | Cultivated var. frutescens (n = 30) | Weedy var. frutescens (n = 30) | Weedy var. crispa (n = 30) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Allele No | MAF | GD | PIC | Allele No | MAF | GD | PIC | Allele No | MAF | GD | PIC | |
| KNUPF127 | 2 | 0.933 | 0.124 | 0.117 | 4 | 0.833 | 0.296 | 0.282 | 4 | 0.567 | 0.562 | 0.486 |
| KNUPF128 | 4 | 0.633 | 0.536 | 0.484 | 6 | 0.533 | 0.618 | 0.561 | 5 | 0.467 | 0.591 | 0.507 |
| KNUPF129 | 8 | 0.300 | 0.796 | 0.766 | 10 | 0.267 | 0.851 | 0.835 | 4 | 0.733 | 0.429 | 0.393 |
| KNUPF130 | 1 | 1.000 | 0.000 | 0.000 | 2 | 0.567 | 0.491 | 0.371 | 2 | 0.767 | 0.358 | 0.294 |
| KNUPF131 | 1 | 1.000 | 0.000 | 0.000 | 3 | 0.733 | 0.418 | 0.370 | 3 | 0.600 | 0.540 | 0.466 |
| KNUPF132 | 6 | 0.433 | 0.664 | 0.605 | 9 | 0.567 | 0.644 | 0.622 | 4 | 0.833 | 0.293 | 0.276 |
| KNUPF133 | 2 | 0.967 | 0.064 | 0.062 | 3 | 0.400 | 0.640 | 0.563 | 2 | 0.900 | 0.180 | 0.164 |
| KNUPF134 | 4 | 0.767 | 0.393 | 0.371 | 6 | 0.533 | 0.618 | 0.561 | 3 | 0.567 | 0.540 | 0.450 |
| KNUPF135 | 1 | 1.000 | 0.000 | 0.000 | 5 | 0.567 | 0.609 | 0.561 | 3 | 0.867 | 0.238 | 0.221 |
| KNUPF136 | 4 | 0.533 | 0.620 | 0.561 | 4 | 0.433 | 0.638 | 0.568 | 4 | 0.833 | 0.293 | 0.276 |
| KNUPF137 | 2 | 0.933 | 0.124 | 0.117 | 3 | 0.800 | 0.331 | 0.294 | 1 | 1.000 | 0.000 | 0.000 |
| KNUPF138 | 1 | 1.000 | 0.000 | 0.000 | 5 | 0.567 | 0.627 | 0.591 | 4 | 0.633 | 0.551 | 0.511 |
| KNUPF139 | 3 | 0.900 | 0.184 | 0.175 | 5 | 0.600 | 0.589 | 0.551 | 3 | 0.633 | 0.531 | 0.475 |
| KNUPF140 | 4 | 0.867 | 0.242 | 0.232 | 3 | 0.567 | 0.518 | 0.414 | 2 | 0.800 | 0.320 | 0.269 |
| KNUPF141 | 3 | 0.600 | 0.540 | 0.466 | 3 | 0.533 | 0.598 | 0.526 | 2 | 0.833 | 0.278 | 0.239 |
| KNUPF142 | 4 | 0.500 | 0.620 | 0.551 | 6 | 0.433 | 0.720 | 0.680 | 2 | 0.900 | 0.180 | 0.164 |
| KNUPF143 | 6 | 0.367 | 0.724 | 0.679 | 6 | 0.333 | 0.791 | 0.762 | 5 | 0.333 | 0.733 | 0.686 |
| Mean | 3.3 | 0.749 | 0.331 | 0.305 | 4.9 | 0.545 | 0.588 | 0.536 | 3.1 | 0.722 | 0.389 | 0.346 |
| Trait | SSR Marker | GLM | SSR Marker | GLM |
|---|---|---|---|---|
| SS | KNUPF132 | ** | KNUPF141 | ** |
| KNUPF133 | ** | KNUPF142 | ** | |
| KNUPF135 | * | KNUPF143 | * | |
| KNUPF136 | ** | KNUPF145 | ** | |
| KNUPF137 | ** | KNUPF146 | ** | |
| KNUPF138 | ** | KNUPF147 | * | |
| KNUPF139 | * | KNUPF148 | ** | |
| KNUPF140 | ** | |||
| SH | KNUPF132 | ** | KNUPF141 | ** |
| KNUPF133 | ** | KNUPF142 | ** | |
| KNUPF135 | * | KNUPF143 | * | |
| KNUPF136 | ** | KNUPF145 | ** | |
| KNUPF137 | ** | KNUPF146 | ** | |
| KNUPF138 | ** | KNUPF147 | * | |
| KNUPF139 | * | KNUPF148 | ** | |
| KNUPF140 | ** | |||
| SCC | KNUPF134 | * | KNUPF145 | * |
| KNUPF135 | * | KNUPF146 | * |
| Source of Variation | Degree of Freedom (df) | Sum of Square (SS) | Variance Component | Percentage of Total Variance | p Value |
|---|---|---|---|---|---|
| Among Pop | 2 | 176.16 | 2.70 | 27 | 0.275 |
| Within Pop | 87 | 619.37 | 7.12 | 73 | 0.001 |
| Total | 89 | 795.52 | 9.82 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Z.Y.; Sa, K.J.; Park, H.; Jang, S.J.; Kim, Y.J.; Lee, J.K. Utilization of Novel Perilla SSR Markers to Assess the Genetic Diversity of Native Perilla Germplasm Accessions Collected from South Korea. Plants 2022, 11, 2974. https://doi.org/10.3390/plants11212974
Fu ZY, Sa KJ, Park H, Jang SJ, Kim YJ, Lee JK. Utilization of Novel Perilla SSR Markers to Assess the Genetic Diversity of Native Perilla Germplasm Accessions Collected from South Korea. Plants. 2022; 11(21):2974. https://doi.org/10.3390/plants11212974
Chicago/Turabian StyleFu, Zhen Yu, Kyu Jin Sa, Hyeon Park, So Jung Jang, Yeon Joon Kim, and Ju Kyong Lee. 2022. "Utilization of Novel Perilla SSR Markers to Assess the Genetic Diversity of Native Perilla Germplasm Accessions Collected from South Korea" Plants 11, no. 21: 2974. https://doi.org/10.3390/plants11212974
APA StyleFu, Z. Y., Sa, K. J., Park, H., Jang, S. J., Kim, Y. J., & Lee, J. K. (2022). Utilization of Novel Perilla SSR Markers to Assess the Genetic Diversity of Native Perilla Germplasm Accessions Collected from South Korea. Plants, 11(21), 2974. https://doi.org/10.3390/plants11212974