

Antifungal Potential of Canarian Plant Extracts against High-Risk Phytopathogens
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Selection of Plants under Study
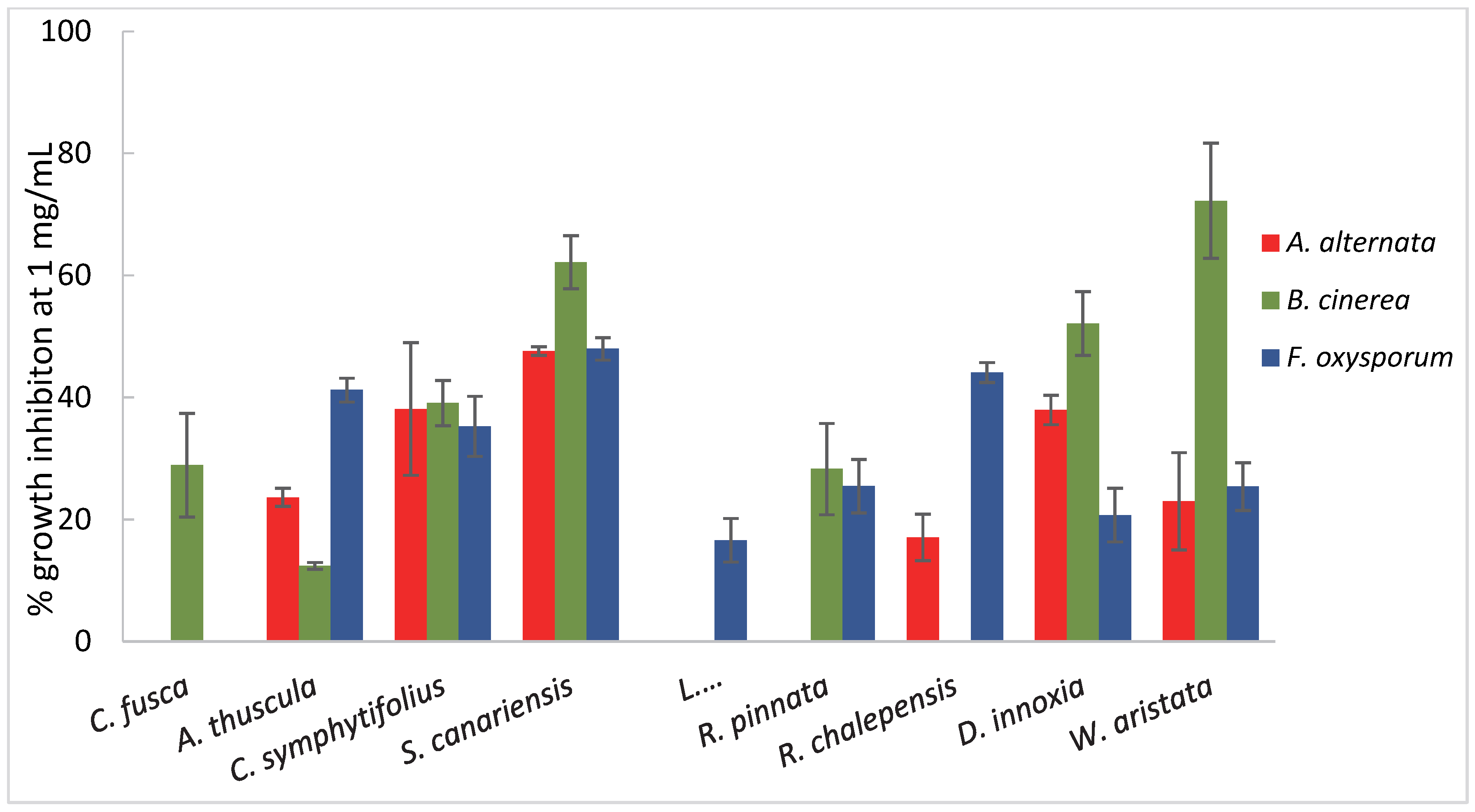
2.2. Screening for Antifungal Bioactivity of Plants’ Ethanolic Extracts
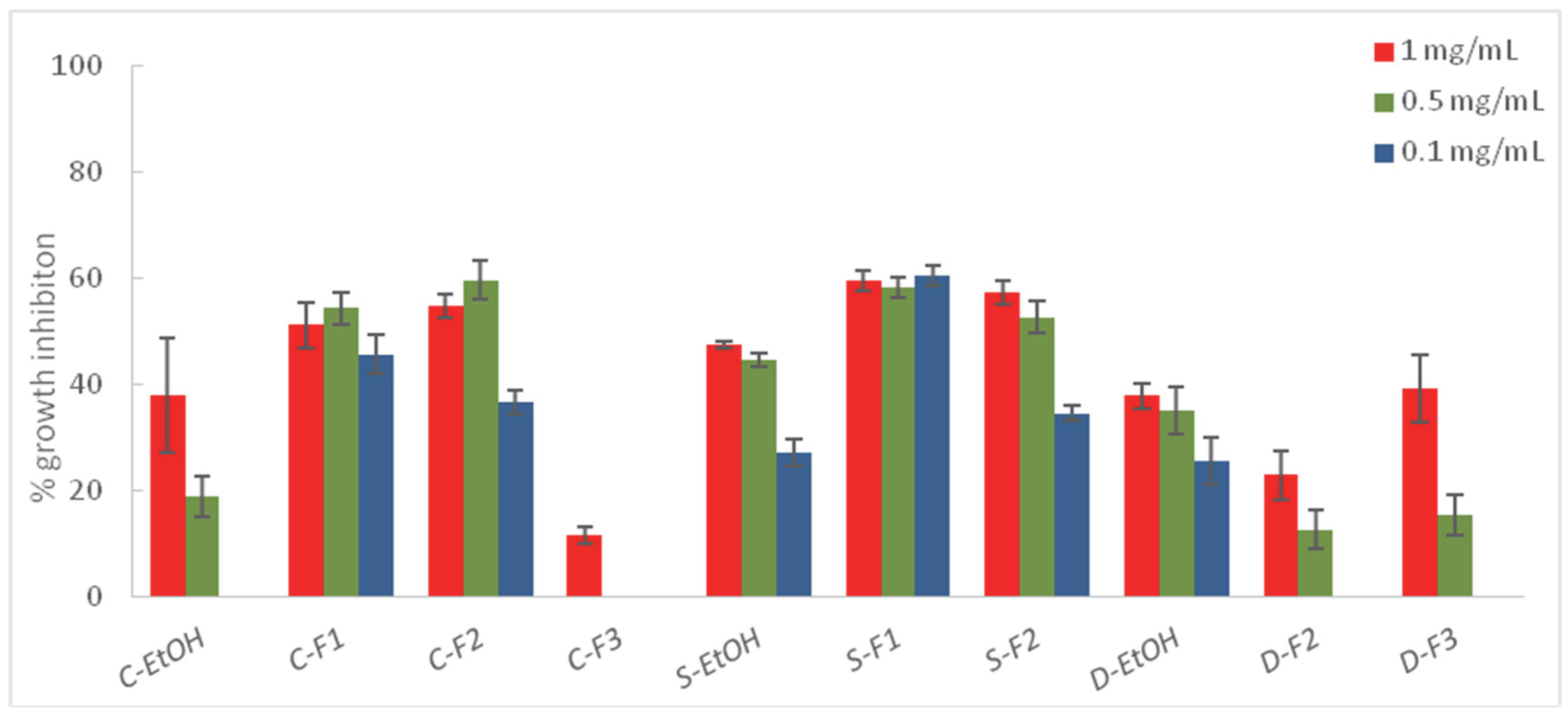
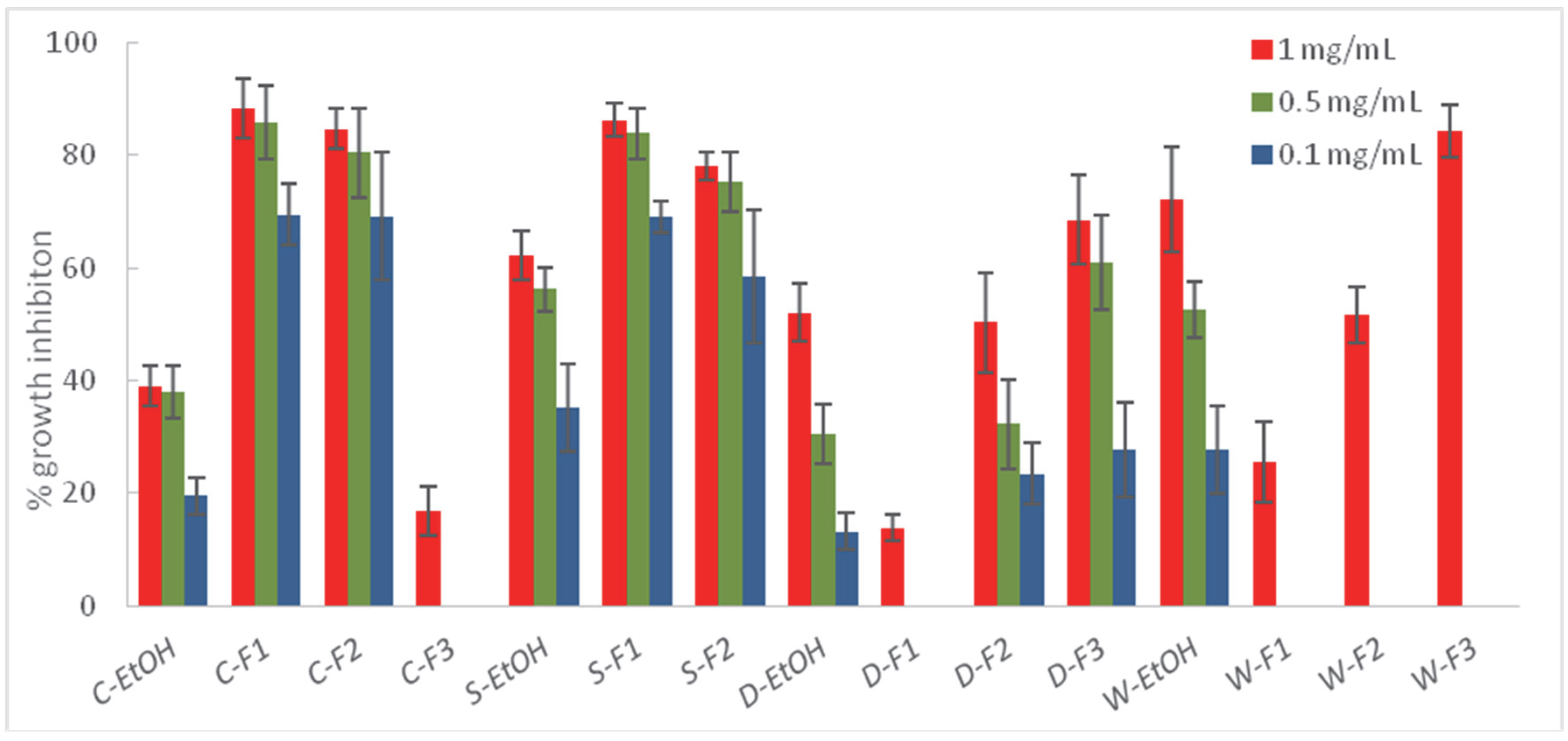
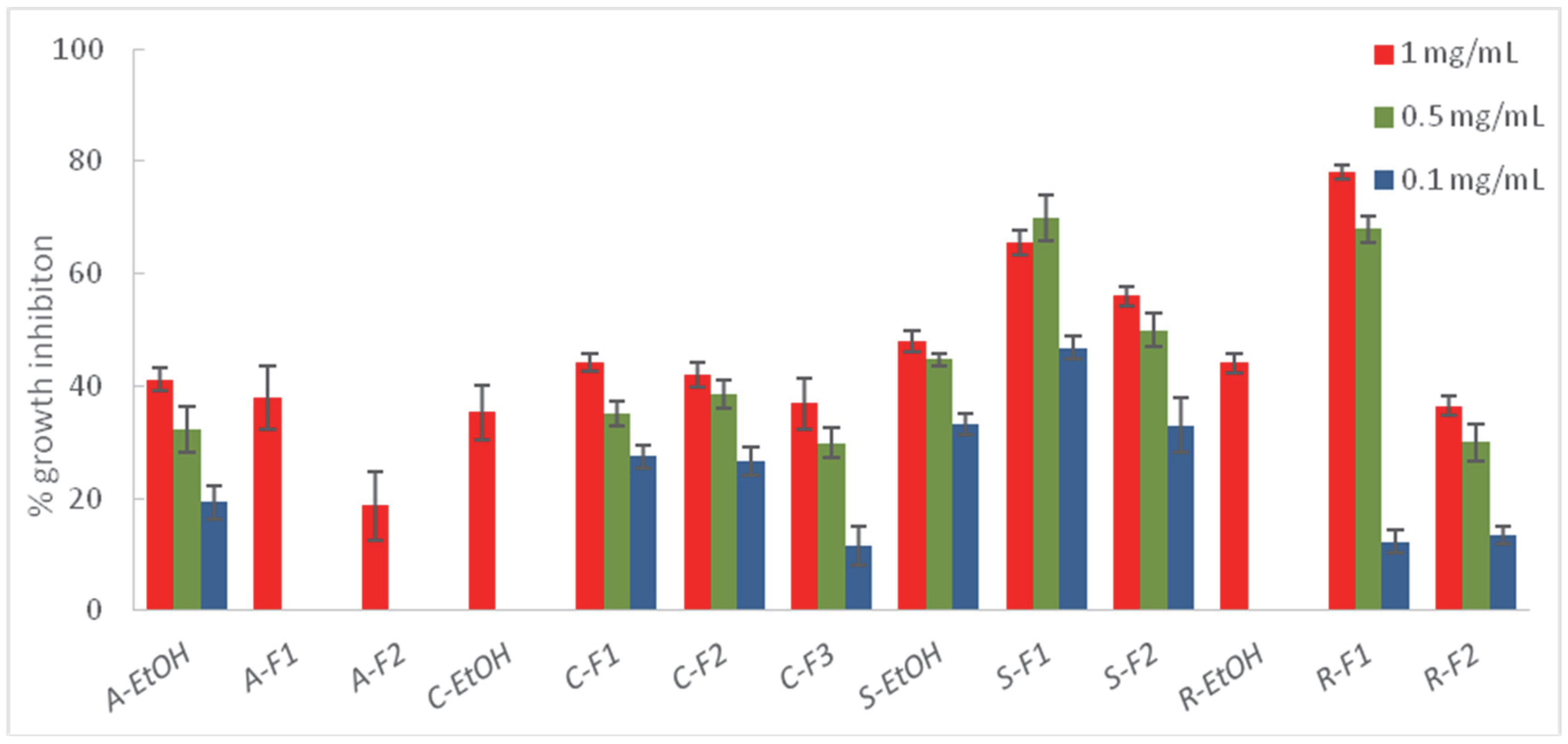
2.3. Liquid–Liquid Partition of the Most Effective Extracts
3. Materials and Methods
3.1. Chemical and Reagents
3.2. Plant Material
3.3. Fungal Culture
3.4. Plant Extracts Preparation
3.5. Liquid–Liquid Partition Procedure of Selected Plant Extracts
3.6. In Vitro Test-Assay on Mycelium
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Bruhl, C.A.; Imfeld, G.; Knabel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmuller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- Pandian, S.; Ramesh, M. Development of pesticide resistance in pests: A key challenge to the crop protection and environmental safety. In Pesticides in Crop Production: Physiological and Biochemical Action, 1st ed.; Srivastava, P.K., Singh, V.P., Singh, A., Tripathi, D.K., Singh, S., Prasad, S.M., Chauhan, D.K., Eds.; Willey: New Jersey, NJ, USA, 2020; Chapter 1; pp. 1–13. [Google Scholar]
- Xu, Z.; Tang, T.; Lin, Q.; Yu, J.; Zhang, C.; Zhao, X.; Kah, M.; Li, L. Environmental risks and the potential benefits of nanopesticides: A review. Environ. Chem. Lett. 2022, 20, 2097–2108. [Google Scholar] [CrossRef]
- Marrone, P.G. Pesticidal natural products–status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef]
- Souto, A.L.; Sylvestre, M.; Tölke, E.D.; Tavares, J.F.; Barbosa-Filho, J.M.; Cebrián-Torrejón, G. Plant-derived pesticides as an alternative to pest management and sustainable agricultural production: Prospects, applications and challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef] [PubMed]
- Gikas, G.D.; Parlakidis, P.; Mavropoulos, T.; Vryzas, Z. Particularities of fungicides and factors affecting their fate and removal efficacy: A review. Sustainability 2022, 14, 4056. [Google Scholar] [CrossRef]
- EU. European Commission Amending Annexes I and V to Regulation (EU) No 649/2012 of the European Parliament and of the Council Concerning the Export and Import of Hazardous Chemicals; EU: Brussels, Belgium, 2018; Volume 2016, pp. 48–119. [Google Scholar]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The Evolution of fungicide resistance. Adv. Appl. Microbiol. 2015, 90, 29–92. [Google Scholar] [CrossRef] [PubMed]
- Marc, R.A. Implications of Mycotoxins in Food Safety. In Mycotoxin and Food Safety-Recent Advances; Marc, R.A., Ed.; IntechOpen: London, UK, 2022; pp. 1–32. ISBN 978-1-83962-904-4. [Google Scholar] [CrossRef]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)—Prospects and challenges. Biocontrol. Sci. Technol. 2019, 29, 241–262. [Google Scholar] [CrossRef]
- Dalmais, B.; Schumacher, J.; Moraga, J.; Le Pêcheur, P.; Tudzynski, B.; Collado, I.G.; Viaud, M. The Botrytis cinerea phytotoxin botcinic acid requires two polyketide synthases for production and has a redundant role in virulence with botrydial. Mol. Plant Pathol. 2011, 12, 564–579. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Munkvold, G.P. Fusarium Species and Their Associated Mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Springer Science-Business Media LLC: New York, NY, USA, 2017; pp. 51–106. ISBN 978-1-4939-6705-6. [Google Scholar]
- Troncoso-Rojas, R.; Tiznado-Hernández, M.E. Alternaria alternata (Black Rot, Black Spot). In Postharvest Decay Control Strategies, 1st ed.; Bautista-Baños, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Chapter 5; pp. 147–187. [Google Scholar] [CrossRef]
- Fernández Pinto, V.E.; Patriarca, A. Alternaria Species and Their Associated Mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols; Moretti, A., Susca, A., Eds.; Springer Science-Business Media LLC: New York, NY, USA, 2017; pp. 13–32. ISBN 978-1-4939-6705-6. [Google Scholar]
- Rodríguez, A.; Acosta, A.; Rodríguez, C. Fungicide resistance of Botrytis cinerea in tomato greenhouses in the Canary Islands and effectiveness of non-chemical treatments against gray mold. World J. Microbiol. Biotechnol. 2014, 30, 2397–2406. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Cabanás, C.G.L.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Hernández, J.; Espino, A.; Rodríguez-Rodríguez, J.M.; Pérez-Sierra, A.; León, M.; Abad-Campos, P.; Armengol, J. Survey of diseases caused by Fusarium spp. on palm trees in the Canary Islands. Phytopathol. Mediterr. 2010, 49, 84–88. [Google Scholar] [CrossRef]
- Yang, L.N.; He, M.H.; Ouyang, H.B.; Zhu, W.; Pan, Z.C.; Sui, Q.J.; Shang, L.P.; Zhan, J. Cross-resistance of the pathogenic fungus Alternaria alternata to fungicides with different modes of action. BMC Microbiol. 2019, 19, 205. [Google Scholar] [CrossRef] [Green Version]
- Schnarr, L.; Segatto, M.L.; Olsson, O.; Zuin, V.G.; Kümmerer, K. Flavonoids as biopesticides—Systematic assessment of sources, structures, activities and environmental fate. Sci. Total Environ. 2022, 824, 153781. [Google Scholar] [CrossRef]
- Coşoveanu, A.; Cabrera, R.; Giménez, C.; Iacomi, B.M.; González-Coloma, A. Antifungal activity of plant extracts against pre and postharvest pathogens. Sci. Pap. Ser. A Agron. 2013, LVI, 206–211. [Google Scholar]
- Satish, S.; Raghavendra, M.P.; Raveesha, K.A. Antifungal potentiality of some plant extracts against Fusarium sp. Arch. Phytopathol. Plant Prot. 2009, 42, 618–625. [Google Scholar] [CrossRef]
- Francisco-Ortega, J.; Santos-Guerra, A.; Kim, S.C.; Crawford, D.J. Plant genetic diversity in the Canary Islands: A conservation perspective. Am. J. Bot. 2000, 87, 909–919. [Google Scholar] [CrossRef] [Green Version]
- Nxumalo, K.A.; Aremu, A.O.; Fawole, O.A. Potentials of medicinal plant extracts as an alternative to synthetic chemicals in postharvest protection and preservation of horticultural crops: A review. Sustainability 2021, 13, 5897. [Google Scholar] [CrossRef]
- Herrera, R.M.; Pérez, M.; Martín-Herrera, D.A.; López-García, R.; Rabanal, R.M.; Arias, A. Antimicrobial activity of extracts from plants endemic to the Canary Islands. Phyther. Res. 1996, 10, 364–366. [Google Scholar] [CrossRef]
- Cruz Suarez, S.J. Más de 100 Plantas Medicinales, 1st ed.; Obra Social de la Caja de Canaria: Las Palmas de Gran Canarias, Spain, 2007. [Google Scholar]
- González, A.G.; Jiménez, I.A.; Ravelo, A.G.; Bazzocchi, I.L. Minor sesquiterpenes from Maytenus canariensis with insecticidal and antifeedant activity. Tetrahedron 1993, 49, 6637–6644. [Google Scholar] [CrossRef]
- González, A.G.; Jiménez, I.A.; Ravelo, A.G.; Coll, J.; González, J.A.; Lloria, J. Antifeedant activity of sesquiterpenes from Celastraceae. Biochem. Syst. Ecol. 1997, 25, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Karim, H.; Boubaker, H.; Askarne, L.; Cherifi, K.; Lakhtar, H.; Msanda, F.; Boudyach, E.H.; Aoumar, A.A.B. Use of Cistus aqueous extracts as botanical fungicides in the control of citrus sour rot. Microb. Pathog. 2017, 104, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Talibi, I.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; Ait Ben Aoumar, A. Antifungal activity of some Moroccan plants against Geotrichum candidum, the causal agent of postharvest citrus sour rot. Crop Prot. 2012, 35, 41–46. [Google Scholar] [CrossRef]
- Abreu, M.E.; Müller, M.; Alegre, L.; Munné-Bosch, S. Phenolic diterpene and α-tocopherol contents in leaf extracts of 60 Salvia species. J. Sci. Food Agric. 2008, 88, 2648–2653. [Google Scholar] [CrossRef]
- Breton-Funes, J.L.; De Atauri, I.J.R. Triterpenes from Lavandula canariensis. J. Nat. Prod. 1986, 49, 937–949. [Google Scholar] [CrossRef]
- Furtado, R.; Baptista, J.; Lima, E.; Paiva, L.; Barroso, J.G.; Rosa, J.S.; Oliveira, L. Chemical composition and biological activities of Laurus essential oils from different Macaronesian Islands. Biochem. Syst. Ecol. 2014, 55, 333–341. [Google Scholar] [CrossRef]
- Rodilla, J.M.; Tinoco, M.T.; Morais, J.C.; Giménez, C.; Cabrera, R.; Martín-Benito, D.; Castillo, L.; González-Coloma, A. Laurus novocanariensis essential oil: Seasonal variation and valorization. Biochem. Syst. Ecol. 2008, 36, 167–176. [Google Scholar] [CrossRef]
- Bouajaj, S.; Romane, A.; Benyamna, A.; Amri, I.; Hanana, M.; Hamrouni, L.; Romdhane, M. Essential oil composition, phytotoxic and antifungal activities of Ruta chalepensis L. leaves from High Atlas Mountains (Morocco). Nat. Prod. Res. 2014, 28, 1910–1914. [Google Scholar] [CrossRef]
- Bouabidi, W.; Hanana, M.; Gargouri, S.; Amri, I.; Fezzani, T.; Ksontini, M.; Jamoussi, B.; Hamrouni, L. Chemical composition, phytotoxic and antifungal properties of Ruta chalepensis L. essential oils. Nat. Prod. Res. 2015, 29, 864–868. [Google Scholar] [CrossRef]
- Haddouchi, F.; Chaouche, T.M.; Zaouali, Y.; Ksouri, R.; Attou, A.; Benmansour, A. Chemical composition and antimicrobial activity of the essential oils from four Ruta species growing in Algeria. Food Chem. 2013, 141, 253–258. [Google Scholar] [CrossRef]
- Sharma, M.; Dhaliwal, I.; Rana, K.; Delta, A.K.; Kaushik, P. Phytochemistry, pharmacology, and toxicology of Datura species—A review. Antioxidants 2021, 10, 1291. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Chandra, R.; Behera, L. Assessment of plant extracts and their in vitro efficacy against potato early blight incited by Alternaria solani. J. Pure Appl. Microbiol. 2021, 15, 1591–1601. [Google Scholar] [CrossRef]
- Matias, R.; Fernandes, V.; Corrêa, B.O.; Pereira, S.R.; de Oliveira, A.K.M. Phytochemistry and antifungal potential of Datura inoxia mill. on soil phytopathogen control. Biosci. J. 2020, 36, 691–701. [Google Scholar] [CrossRef]
- Mdee, L.K.; Masoko, P.; Eloff, J.N. The activity of extracts of seven common invasive plant species on fungal phytopathogens. S. Afr. J. Bot. 2009, 75, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Mareggiani, G.; Picollo, M.I.; Veleiro, A.S.; Tettamanzi, M.C.; Benedetti-Doctorovich, M.O.V.; Burton, G.; Zerba, E. Response of Tribolium castaneum (Coleoptera, Tenebrionidae) to Salpichroa origanifolia withanolides. J. Agric. Food Chem. 2002, 50, 104–107. [Google Scholar] [CrossRef]
- LLanos, G.G.; Araujo, L.M.; Jiménez, I.A.; Moujir, L.M.; Bazzocchi, I.L. Withaferin A-related steroids from Withania aristata exhibit potent antiproliferative activity by inducing apoptosis in human tumor cells. Eur. J. Med. Chem. 2012, 54, 499–511. [Google Scholar] [CrossRef]
- de Paz, P.L.P.; Padrón, C.E.H. Plantas Medicinales o Útiles en la Flora Canaria: Aplicaciones Populares; Francisco Lemus: La Laguna, Spain, 1999. [Google Scholar]
- Margarita de Monte. CanariWiki, Consejería de Educación, Universidades, Cultura y Deportes Gobierno de Canarias. Available online: https://www3.gobiernodecanarias.org/medusa/wiki/index.php?title=Margarita_de_monte (accessed on 30 September 2022).
- Bramwell, D. Plantas Medicinales de las Islas Canarias; Rueda: Madrid, Spain, 2004. [Google Scholar]
- Amagante. CanariWiki, Consejería de Educación, Universidades, Cultura y Deportes, Gobierno de Canarias. Available online: https://www3.gobiernodecanarias.org/medusa/wiki/index.php?title=Amagante (accessed on 30 September 2022).
- Llorent-Martínez, E.J.; Spínola, V.; Castilho, P.C. Phenolic profiles of Lauraceae plant species endemic to Laurisilva forest: A chemotaxonomic survey. Ind. Crop. Prod. 2017, 107, 1–12. [Google Scholar] [CrossRef]
- Jeon, J.H.; Kim, M.G.; Lee, H.S. Insecticidal activities of Ruta chalepensis leaves isolated constituent and structure-relationships of its analogues against Sitophilus oryzae. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 591–596. [Google Scholar] [CrossRef]
- Aouadhi, C.; Ghazghazi, H.; Hamrouni, S.; Hasnaoui, B.; Maaroufi, A. In vitro antifungal activity of the essential oils and the methanolic extract of Ruta chalepensis. Archs. Inst. Pasteur Tunis 2013, 90, 39–46. [Google Scholar]
- Miller, A.B.; Cates, R.G.; Lawrence, M.; Soria, J.A.F.; Espinoza, L.V.; Martínez, J.V.; Arbizú, D.A. The antibacterial and antifungal activity of essential oils extracted from Guatemalan medicinal plants. Pharm. Biol. 2015, 53, 548–554. [Google Scholar] [CrossRef]
- Swathi, S.; Murugananthan, G.; Ghosh, S.; Pradeep, A.S. Larvicidal and repellent activities of ethanolic extract of Datura stramonium leaves against mosquitoes. Int. J. Pharm. Phytochem. Res. 2012, 4, 25–27. [Google Scholar]
- Pascual-Villalobos, M.J.; Robledo, A. Screening for anti-insect activity in Mediterranean plants. Ind. Crop. Prod. 1998, 8, 183–194. [Google Scholar] [CrossRef]
- Ali, S.M.A.; Ammad, A.M.; Ahmad, S.K.; Bilal, I.M.; Abdullah, A.; Talal, I.; Zulfiqar, M. Effectiveness of different plant extracts along with new chemistry insecticide against Tribolium castaneum. GSC Biol. Pharm. Sci. 2020, 13, 95–100. [Google Scholar] [CrossRef]
- Antigales, Huevito de Gallo. Available online: http://antigales.blogspot.com/2016/08/huevito-de-gallo.html (accessed on 30 September 2022).
- Llanos, G.G.; Varela, R.M.; Jiménez, I.A.; Molinillo, J.M.; Macías, F.A.; Bazzocchi, I.L. Metabolites from Withania aristata with potential phytotoxic activity. Nat. Prod. Commun. 2010, 5, 1043–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haman, C.; Dauchy, X.; Rosin, C.; Munoz, J.-F. Occurrence, fate and behavior of parabens in aquatic environments: A review. Water Res. 2015, 68, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zalegh, I.; Akssira, M.; Bourhia, M.; Mellouki, F.; Rhallabi, N.; Salamatullah, A.M.; Alkaltham, M.S.; Alyahya, H.K.; Mhand, R.A. A review on Cistus sp.: Phytochemical and antimicrobial activities. Plants 2021, 10, 1214. [Google Scholar] [CrossRef]
- Papaefthimiou, D.; Papanikolaou, A.; Falara, V.; Givanoudi, S.; Kostas, S.; Kanellis, A.K. Genus Cistus: A model for exploring labdane-type diterpenes’ biosynthesis and a natural source of high value products with biological, aromatic, and pharmacological properties. Front. Chem. 2014, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Calabuig, M.T.; Cortés, M.; Francisco, C.G.; Hernández, R.; Suárez, E. Labdane diterpenes from Cistus symphytifolius. Phytochemistry 1981, 20, 2255–2258. [Google Scholar] [CrossRef]
- Molina, Á.V.; Guerra, A.S. Investigaciones Fitoquímicas en Plantas Canarias; Centro de Estudios Ramón Aceres: Madrid, Spain, 2002. [Google Scholar]
- Exarchou, V.; Kanetis, L.; Charalambous, Z.; Apers, S.; Pieters, L.; Gekas, V.; Goulas, V. HPLC-SPE-NMR characterization of major metabolites in Salvia fruticosa mill. extract with antifungal potential: Relevance of carnosic acid, carnosol, and hispidulin. J. Agric. Food Chem. 2015, 63, 457–463. [Google Scholar] [CrossRef]
- Shu, C.; Zhao, H.; Jiao, W.; Liu, B.; Cao, J.; Jiang, W. Antifungal efficacy of ursolic acid in control of Alternaria alternata causing black spot rot on apple fruit and possible mechanisms involved. Sci. Hortic. 2019, 256, 108636. [Google Scholar] [CrossRef]
- Abdel-Motaal, F.F.; Nassar, M.S.M.; El-Zayat, S.A.; El-Sayed, M.A.; Ito, S. Responses of fungi to tropane alkaloids produced by a medicinal plant Hyoscyamus muticus (Egyptian henbane). Folia Microbiol. 2009, 54, 207–212. [Google Scholar] [CrossRef] [PubMed]
- El Bazaoui, A.; Bellimam, M.A.; Soulaymani, A. Tropane alkaloids of Datura innoxia from Morocco. Z. Naturforsch. C J. Biosci. 2012, 67, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Seepe, H.A.; Ramakadi, T.G.; Lebepe, C.M.; Amoo, S.O.; Nxumalo, W. Antifungal activity of isolated compounds from the leaves of Combretum erythrophyllum (Burch.) Sond. and Withania somnifera (L.) Dunal against Fusarium pathogens. Molecules 2021, 26, 4732. [Google Scholar] [CrossRef] [PubMed]
- Kupchan, S.M. Recent advances in the chemistry of terpenoid tumor inhibitors. Pure Appl. Chem. 1970, 21, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Cosoveanu, A.; Hernandez, M.; Iacomi-Vasilescu, B.; Zhang, X.; Shu, S.; Wang, M.; Cabrera, R. Fungi as endophytes in Chinese Artemisia spp.: Juxtaposed elements of phylogeny, diversity and bioactivity. Mycosphere 2016, 7, 102–117. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Plant Species/Common Name/ Voucher Specimen 1 | Ecological State 2/ Plant Organ | Collection Place 3/Geographic Coordinates 4/Date |
|---|---|---|---|
| Apocynaceae | Ceropegia fusca Bolle/ cardoncillo, TFC 53.261 | Endemic/ Aerial part | Montaña Pelada, Granadilla de Abona/ 28°04′03″ N, 16°31′18″ W, 100 m.a.s.l./09-2018 |
| Asteraceae | Argyranthemum broussonetii Pers. & Humphries/ magarza de monte/TFC 53.398 | Endemic/ Aerial part | Ctra. El Bailadero, La Cumbrilla, Anaga/ 28°32′12″ N, 16°14′04″ W, 830 m.a.s.l./ 06-2020 |
| Artemisia thuscula Cav./ incienso/TFC 53.702 | Endemic/ Aerial part | Ctra. Playa Santo Domingo, La Guancha/ 28°23′32″ N, 16°40′34″ W, 88 m.a.s.l./02-2021 | |
| Celastraceae | Gymnosporia cassinoides (L’Hér.) Masf. (=Maytenus canariensis (Loes.) G. Kunkel & Sunding)/peralillo/TFC 53.244 | Endemic/ Leaves | Centro Ambiental La Tahonilla, La Laguna/ 28°28′34″ N, 16°19′16″ W, 574 m.a.s.l./ 09-2018 |
| Cistaceae | Cistus symphytifolius Lam./ jara/TFC 53.703 | Endemic/ Leaves | Pista Forestal El Lagar, La Guancha/ 28°20′42″ N, 16°39′15″ W, 1023 m.a.s.l./02-2019 |
| Lamiaceae | Lavandula canariensis Mill./ lavanda, mato risco/TFC 53.400 | Endemic/ Aerial part | Rambla de Castro, Los Realejos/ 28°23′40″ N, 16°35′32″ W, 125 m.a.s.l./05-2020 |
| Salvia canariensis L./ salvia, salvia morisca/TFC 53.328 | Endemic/ Leaves | Gáldar/ 28°02′10″ N, 15°37′30″ W, 1415 m.a.s.l./06-2019 | |
| Lauraceae | Apollonias barbujana (Cav.) A. Braun subsp. barbujana/ barbusano, ébano de Canarias/ TFC 53.397 | Endemic/ Leaves | La Laguna/ 28°28′48″ N, 16°19′11″ W, 560 m.a.s.l./ 01-2020 |
| Laurus novocanariensis Rivas- Mart. et al./loro, laurel/ TFC 53.399 | Endemic/ Leaves | Cruz del Carmen, Anaga/ 28°31′63″ N, 16°16′48″ W, 970 m.a.s.l./ 06-2020 | |
| Rutaceae | Ruta pinnata L. f./ ruda salvaje/TFC 53.243 | Endemic/ Leaves | La Laguna/ 28°28′44″ N, 16°19′21″ W, 575 m.a.s.l./09-2018 |
| Ruta chalepensis L./ ruda/TFC 53.756 | Native to Macaronesian/Leaves | Araya, Candelaria/ 28°21′24″ N, 16°23′27″ W, 320 m.a.s.l./09-2018 | |
| Solanaceae | Datura innoxia Mill./ burladora, métel/TFC 53.216 | Non-native in the Canary Islands/Leaves | La Orotava/ 28°23′40″ N, 16°32′46″ W, 198 m.a.s.l./ 03-2018 |
| Datura stramonium L./ estramonio, hierba del diablo, santas noches/TFC 53.217 | Non-native in the Canary Islands/Leaves | La Orotava/ 28°23′40″ N, 16°32′46″ W, 198 m.a.s.l./ 03-2018 | |
| Nicotiana glauca Graham/ bobo, tabaco moro/TFC 53.214 | Invasive in the Canary Islands/Leaves | La Orotava/ 28°23′40″ N, 16°32′46″ W, 198 m.a.s.l./03-2018 | |
| Salpichroa origanifolia (Lam.) Baill./ huevito de gallo/TFC 53.215 | Non-native in the Canary Islands/Leaves | La Orotava/ 28°23′40″ N, 16°32′46″ W, 198 m.a.s.l./03-2018 | |
| Withania aristata (Aiton) Pauquy/ orobal/TFC 53.219 | Native to the Canary Islands/Leaves | Ctra. Playa Santo Domingo, La Guancha/ 28°23′33″ N, 16°40′32″ W, 88 m.a.s.l./04-2018 |
| Plant Species | Traditional/Scientific Uses | References |
|---|---|---|
| C. fusca | Vulnerary, anti-hemorrhagic | [44] |
| A. broussonetii | Stomach remedy, anti-asthmatic, used to regulate menstrual disorders | [45] |
| A. thuscula | Insecticide, antiseptic, Protection of potato crops from insects | [46] [26] |
| G. cassinoides | Antiviral, antifungal Insecticidal, antifeedant against Spodoptera littoralis | [46] [27] |
| C. symphytifolius | Analgesic | [47] |
| L. canariensis | Vermifuge, antiparasitic, antiseptic Insecticide, anti-inflammatory, diuretic, sedative | [46] [26] |
| S. canariensis | Antiseptic, antiviral Antifungal, antibacterial | [44] [26] |
| A. barbujana | Diuretic, analgesic, antiulcerogenic, cardiotonic, expectorant, stomachic, sedative | [48] |
| L. novocanariensis | To treat bronchitis, pharyngitis, rheumatism Insecticidal | [26] [46] |
| R. pinnata | Antiparasitic, antiseptic Antibiotic, antifungal, anti-inflammatory, toxic | [46] [26] |
| R. chalepensis | Toxic Insecticide against Sitophilus oryzae Phytotoxic and antifungal activities Antibacterial and antifungal activity | [44] [49] [36,50] [51] |
| D. innoxia | Anti-asthmatic, anesthetic, toxic | [44] |
| D. stramonium | Antiasthmatic, antiparkinsonian, narcotic, antispasmodic, toxic Larvicidal and repellent against mosquitoes Anti-insect activity in Mediterranean plants Insecticide against Tribolium castaneum | [44,46] [52] [53] [54] |
| N. glauca | Dermatic, toxic Active against phytopathogenic fungi | [44] [41] |
| S. origanifolia | Anti-inflammatory, heal skin abrasions, diuretic, narcotic Isolated withanolides act as feeding inhibitors on Tribolium castaneum | [55] [42] |
| W. aristata | Diuretic, analgesic, anesthetic, anti-asthmatic, antirheumatic, antitumor, aphrodisiac Apocarotenoids and carotenoids showed phytotoxicity towards Lepidium sativum, Lactuca sativa, Lycopersicum esculentum, and Allium cepa | [26,46] [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes, C.P.; Sabina, S.R.; López-Cabeza, R.; Montelongo, C.G.; Giménez, C.; Jiménez, I.A.; Cabrera, R.; Bazzochi, I.L. Antifungal Potential of Canarian Plant Extracts against High-Risk Phytopathogens. Plants 2022, 11, 2988. https://doi.org/10.3390/plants11212988
Reyes CP, Sabina SR, López-Cabeza R, Montelongo CG, Giménez C, Jiménez IA, Cabrera R, Bazzochi IL. Antifungal Potential of Canarian Plant Extracts against High-Risk Phytopathogens. Plants. 2022; 11(21):2988. https://doi.org/10.3390/plants11212988
Chicago/Turabian StyleReyes, Carolina P., Samuel Rodríguez Sabina, Rocío López-Cabeza, Cristina G. Montelongo, Cristina Giménez, Ignacio A. Jiménez, Raimundo Cabrera, and Isabel L. Bazzochi. 2022. "Antifungal Potential of Canarian Plant Extracts against High-Risk Phytopathogens" Plants 11, no. 21: 2988. https://doi.org/10.3390/plants11212988
APA StyleReyes, C. P., Sabina, S. R., López-Cabeza, R., Montelongo, C. G., Giménez, C., Jiménez, I. A., Cabrera, R., & Bazzochi, I. L. (2022). Antifungal Potential of Canarian Plant Extracts against High-Risk Phytopathogens. Plants, 11(21), 2988. https://doi.org/10.3390/plants11212988