

Using of Essential Oils and Plant Extracts against Pseudomonas savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens on Soybean


Abstract
:1. Introduction
2. Results
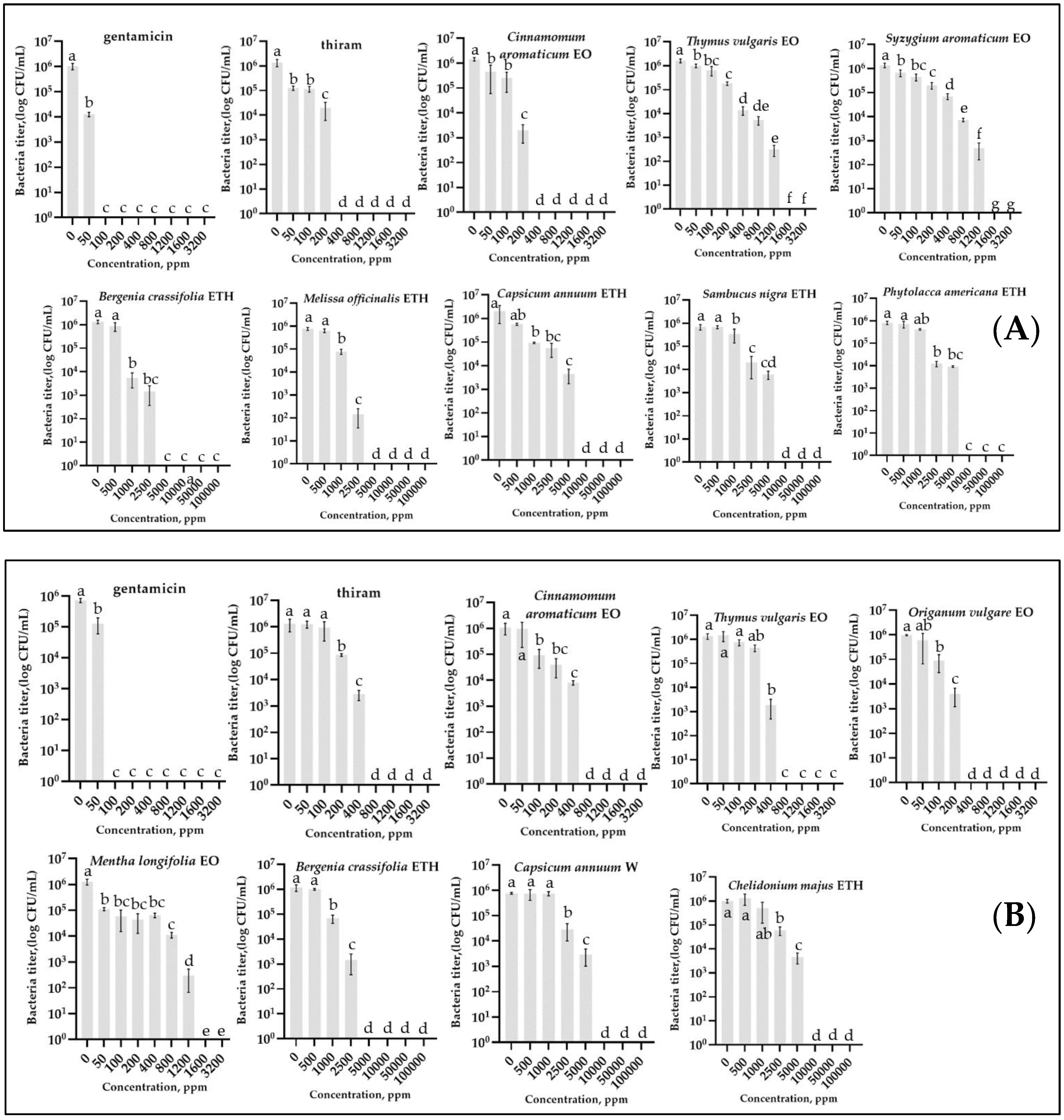
2.1. Antibacterial In Vitro Activity
2.1.1. Antibacterial In Vitro Activity by Disc Diffusion Method
2.1.2. Antibacterial In Vitro Activity by Determination of MIC and MBC Values
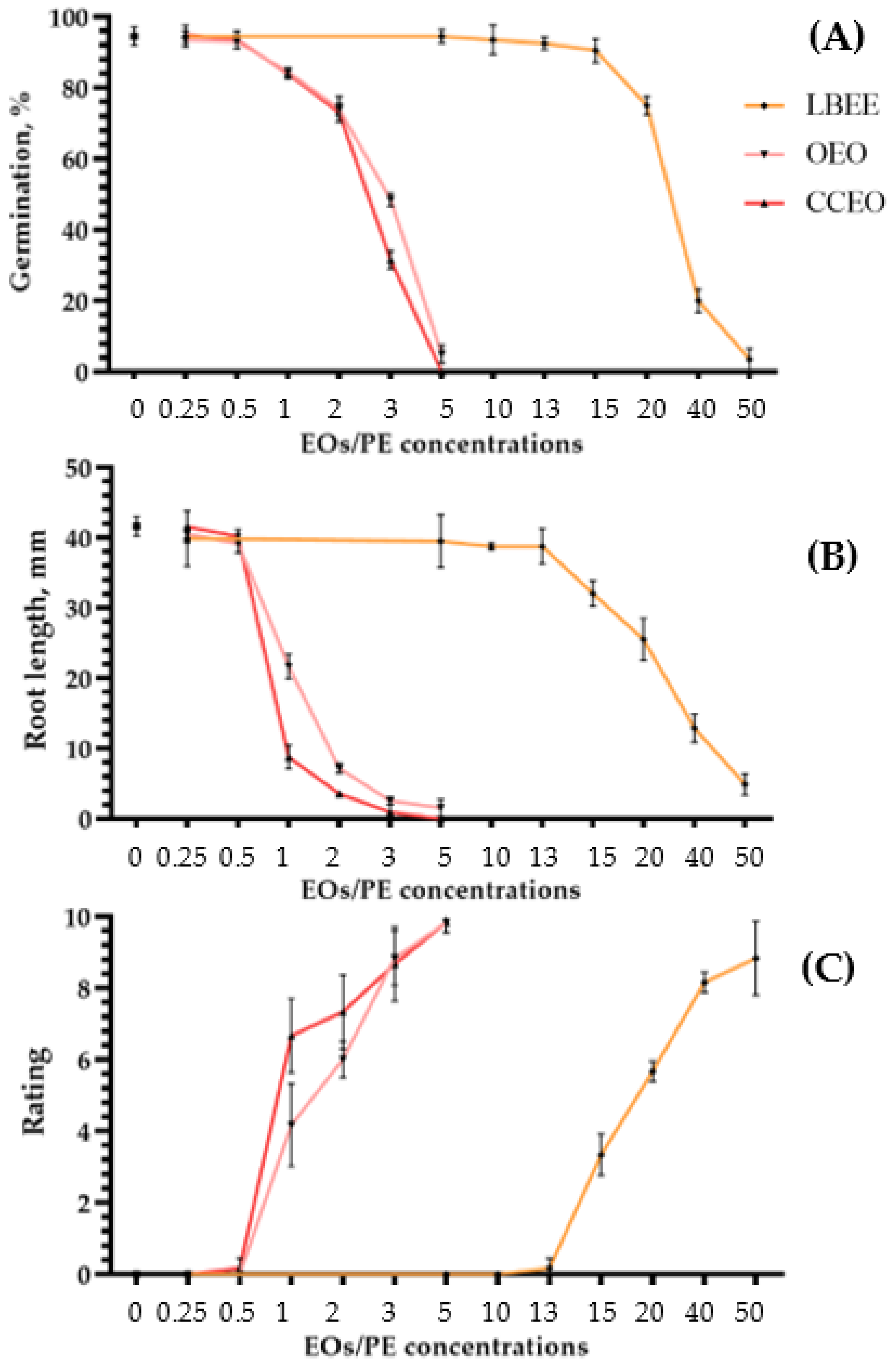
2.2. Phytotoxicity
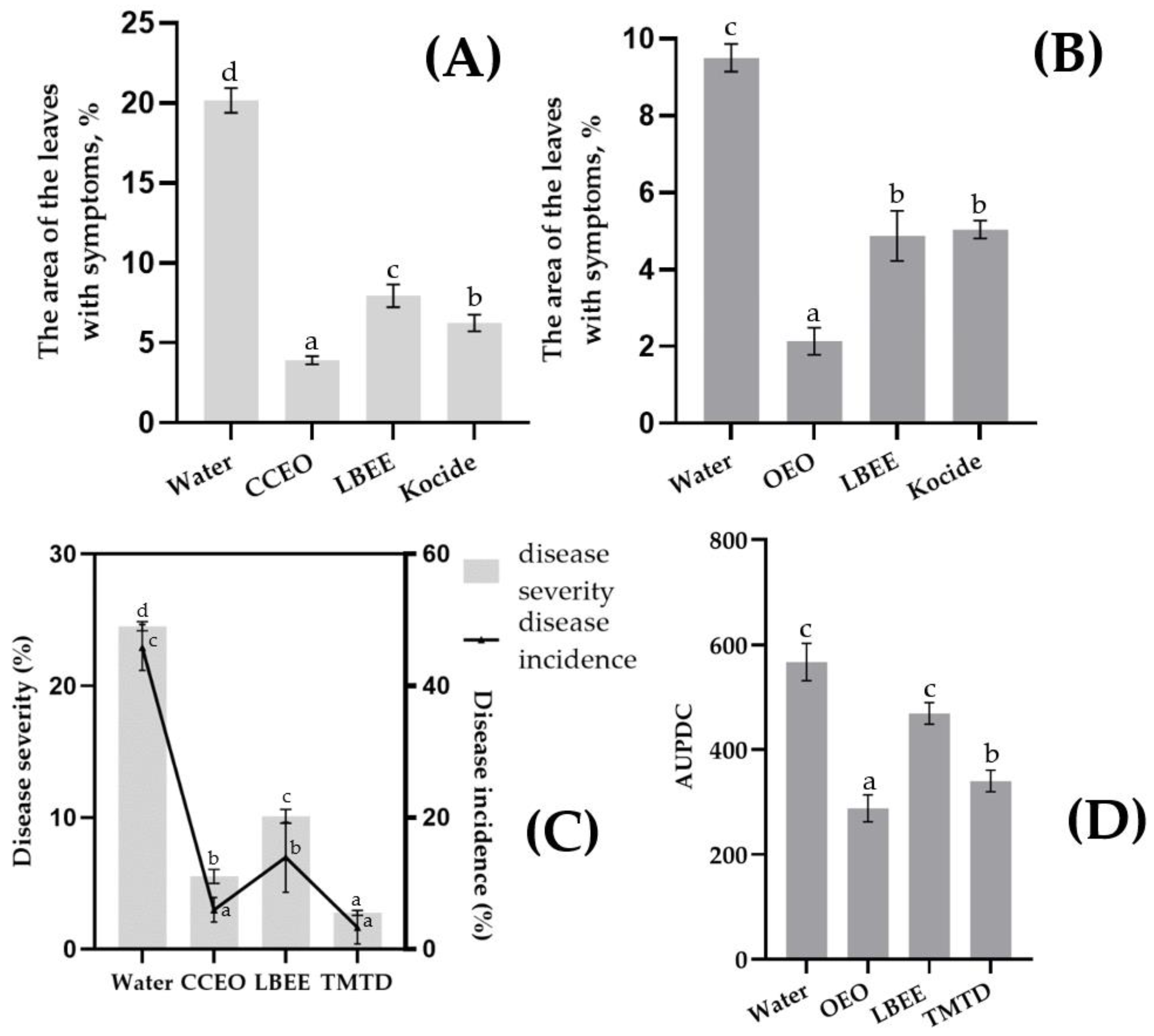
2.3. Control of Seed and Leaf Infections Psg and Cff with EOs and PE
2.4. Identification of Chemicals Comprising EOs u PEs
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Plant Material
4.2.1. Extraction of Essential Oils
4.2.2. Extraction of Plant Extracts
4.3. Determination of Antibacterial Activity
4.3.1. Determination of Antibacterial Activity by Disc Diffusion Method
4.3.2. Determination of MIC and MBC
4.4. Phytotoxicity on Soybean Seeds and Plants
4.5. Gas Chromatography with Flame Ionisation Detector and Mass Spectrometry of Selected EOs and PE
4.6. Control Psg and Cff Artificial Infection Using EOs and PE
4.6.1. Control Psg on Seeds
4.6.2. Control Psg on Leaves
4.6.3. Control Cff on Seeds
4.6.4. Control Cff on Leaves
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LBEE | Leather Bergenia Ethanolic Extract |
| CCEO | Chinese Cinnamon Essential Oil |
| OEO | Oregano Essential Oil |
| a.i. | active ingredient |
| WSC | Water Suspension Concentrate |
| WDG | Water-Dispersible Granules |
| PGPR | Plant Growth-Promoting Rhizobacteria |
| MSD | Mass Spectrometric Detector |
| FID | Flame Ionization Detector |
References
- Chen, K.I.; Erh, M.H.; Su, N.W.; Liu, W.H.; Chou, C.C.; Cheng, K.C. Soyfoods and soybean products: From traditional use to modern applications. Appl. Microbiol. Biotechnol. 2012, 96, 9–22. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture—Statistical Yearbook; FAO: Rome, Italy, 2021. [Google Scholar]
- Allen, T.W.; Bradley, C.A.; Sisson, A.J.; Byamukama, E.; Chilvers, M.I.; Coker, C.M.; Collins, A.A.; Damicone, J.P.; Dorrance, A.E.; Dufault, N.S.; et al. Soybean Yield Loss Estimates Due to Diseases in the United States and Ontario, Canada, from 2010 to 2014. Plant Health Prog. 2017, 18, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Abudulai, M.; Salifu, A.B.; Opare-Atakora, D.; Haruna, M.; Denwar, N.N.; Baba, I.I.Y. Yield Loss at the Different Growth Stages in Soybean Due to Insect Pests in Ghana. Arch. Phytopathol. Plant Prot. 2012, 45, 1796–1809. [Google Scholar] [CrossRef]
- Soltani, N.; Dille, J.; Burke, I.; Everman, W.; VanGessel, M.; Davis, V.; Sikkema, P. Perspectives on Potential Soybean Yield Losses from Weeds in North America. Weed Technol. 2017, 31, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Jagtap, G.P.; Dhopte, S.B.; Dey, U. Original Article Bio-Efficacy of Different Antibacterial Antibiotic, Plant Extracts and Bioagents against Bacterial Blight of Soybean Caused by Pseudomonas Syringae pv. Glycinea. Sci. J. Microbiol. 2012, 1, 1–9. [Google Scholar]
- Zhang, J.; Wang, X.; Lu, Y.; Bhusal, S.J.; Song, Q.; Cregan, P.B.; Yen, Y.; Brown, M.; Jiang, G.L. Genome-Wide Scan for Seed Composition Provides Insights into Soybean Quality Improvement and the Impacts of Domestication and Breeding. Mol. Plant 2018, 11, 460–472. [Google Scholar] [CrossRef] [Green Version]
- Ignjatov, M.; Milošević, M.; Nikolić, Z.; Vujaković, M.; Petrović, D. Characterization of Pseudomonas Savastanoi pv. Glycinea Isolates from Vojvodina. Phytopathol. Pol. 2007, 45, 43–54. [Google Scholar]
- Shepherd, L.M.; Block, C.C. Chapter 13: Detection of Pseudomonas savastanoi pv. glycinea in Soybean Seeds. In Detection of Plant-Pathogenic Bacteria in Seed and Other Planting Material, 2nd ed.; The American Phytopathological Society: St. Paul, MN, USA, 2017; ISBN 978-0-89054-539-3. [Google Scholar]
- Monteil, C.L.; Yahara, K.; Studholme, D.J.; Mageiros, L.; Méric, G.; Swingle, B.; Morris, C.E.; Vinatzer, B.A.; Sheppard, S.K. Population-Genomic Insights into Emergence, Crop Adaptation and Dissemination of Pseudomonas Syringae Pathogens. Microb. Genom. 2016, 2, e000089. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Erickson, R.S.; Balasubramanian, P.M.; Hsieh, T.F.; Conner, R.L. Resurgence of bacterial wilt of common bean in North America. Can. J. Plant Pathol. 2009, 31, 290–300. [Google Scholar] [CrossRef]
- Soares, R.M.; Fantinato, G.G.P.; Darben, L.M.; Marcelino-Guimarães, F.C.; Seixas, C.D.S.; Carneiro, G.E.D.S. First Report of Curtobacterium Flaccumfaciens pv. Flaccumfaciens on Soybean in Brazil. Trop. Plant Pathol. 2013, 38, 452–454. [Google Scholar] [CrossRef]
- Huang, H.C.; Mundel, H.H.; Erickson, R.S.; Chelle, C.D.; Balasubramanian, P.M.; Kiehn, F.; Conner, R. Resistance of common bean (Phaseolus vulgaris L.) cultivars and germplasm lines to the purple variant of bacterial wilt (Curtobacterium flaccumfaciens pv. flaccumfaciens). Plant Pathol. Bull. 2007, 16, 91–95. [Google Scholar]
- Camara, R.C.; Vigo, S.C.; Maringoni, A.C. Plant to seed transmission of Curtobacterium flaccumfaciens pv. flaccumfaciens in a dry bean cultivar. J. Plant Pathol. 2009, 91, 549–554. [Google Scholar]
- Hsieh, T.F.; Huang, H.C.; Erickson, R.S. Bacterial wilt of common bean: Effect of seed-borne inoculum on disease incidence and seedling vigor. Seed Sci. Technol. 2006, 34, 57–67. [Google Scholar] [CrossRef]
- Silva, T.A.F., Jr.; Itako, A.T.; Negrão, D.R.; Maringoni, A.C. Pathogenicity of Curtobacterium flaccumfaciens pv. flaccumfaciens to several plant species. J. Plant Pathol. 2012, 94, 427–430. [Google Scholar]
- Urrea, C.A.; Harveson, R.M. Identification of sources of bacterial wilt resistance in common beans (Phaseolus vulgaris L.). Plant Dis. 2014, 98, 973–976. [Google Scholar] [CrossRef]
- Estefani, R.C.C.; Miranda Filho, R.J.; Uesugi, C.H. Tratamentos térmico e químico de sementes de feijoeiro: Eficiência na erradicação de Curtobacterium flaccumfaciens pv. flaccumfaciens e efeitos na qualidade fisiológica das sementes. Fitopatol. Bras. 2007, 32, 434–438. [Google Scholar] [CrossRef]
- Cazorla, F.M.; Arrebola, E.; Sesma, A.; Pérez-García, A.; Codina, J.C.; Murillo, J.; de Vicente, A. Copper Resistance in Pseudomonas Syringae Strains Isolated from Mango Is Encoded Mainly by Plasmids. Phytopathology 2002, 92, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Völksch, B.; May, R. Biological Control of Pseudomonas Syringae pv. Glycinea by Epiphytic Bacteria under Field Conditions. Microb. Ecol. 2001, 41, 132–139. [Google Scholar] [CrossRef]
- Martins, S.J.; de Medeiros, F.H.V.; de Souza, R.M.; de Resende, M.L.V.; Ribeiro, P.M. Biological Control of Bacterial Wilt of Common Bean by Plant Growth-Promoting Rhizobacteria. Biol. Control 2013, 66, 65–71. [Google Scholar] [CrossRef]
- Tarakanov, R.I.; Lukianova, A.A.; Evseev, P.V.; Toshchakov, S.V.; Kulikov, E.E.; Ignatov, A.N.; Miroshnikov, K.A.; Dzhalilov, F.S.-U. Bacteriophage Control of Pseudomonas Savastanoi pv. Glycinea in Soybean. Plants 2022, 11, 938. [Google Scholar] [CrossRef]
- Tarakanov, R.I.; Lukianova, A.A.; Evseev, P.V.; Pilik, R.I.; Tokmakova, A.D.; Kulikov, E.E.; Toshchakov, S.V.; Ignatov, A.N.; Dzhalilov, F.S.-U.; Miroshnikov, K.A. Ayka, a Novel Curtobacterium Bacteriophage, Provides Protection against Soybean Bacterial Wilt and Tan Spot. Int. J. Mol. Sci. 2022, 23, 10913. [Google Scholar] [CrossRef]
- Orzali, L.; Valente, M.T.; Scala, V.; Loreti, S.; Pucci, N. Antibacterial Activity of Essential Oils and Trametes Versicolor Extract against Clavibacter michiganensis Subsp. Michiganensis and Ralstonia Solanacearum for Seed Treatment and Development of a Rapid In Vivo Assay. Antibiotics 2020, 9, 628. [Google Scholar]
- Steglińska, A.; Bekhter, A.; Wawrzyniak, P.; Kunicka-Styczyńska, A.; Jastrząbek, K.; Fidler, M.; Śmigielski, K.; Gutarowska, B. Antimicrobial Activities of Plant Extracts against Solanum Tuberosum L. Phytopathogens. Molecules 2022, 27, 1579. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, G.; Pucci, N.; Brasili, E.; Valletta, A.; Sammarco, I.; Carnevale, E.; Pasqua, G.; Loreti, S. In vitro antimicrobial activity of plant extracts against Pseudomonas syringae pv. actinidiae causal agent of bacterial canker in kiwifruit. Plant Biosyst. 2020, 154, 100–106. [Google Scholar] [CrossRef]
- Diánez, F.; Santos, M.; Parra, C.; Navarro, M.; Blanco, R.; Gea, F. Screening of antifungal activity of 12 essential oils against eight pathogenic fungi of vegetables and mushroom. Lett. Appl. Microbiol. 2018, 67, 400–410. [Google Scholar] [CrossRef]
- Amini, J.; Farhang, V.; Javadi, T.; Nazemi, J. Antifungal Effect of Plant Essential Oils on Controlling Phytophthora Species. Plant Pathol. J. 2016, 32, 16–24. [Google Scholar] [CrossRef]
- Satish, S.; Raveesha, K.A.; Janardhana, G.R. Antibacterial activity of plant extracts on phytopathogenic Xanthomonas campestris pathovars. Lett. Appl. Microbiol. 1999, 28, 145–147. [Google Scholar] [CrossRef]
- Sikkema, J.; de Bont, J.A.M.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, F.; Ji, B.P.; Pei, R.S.; Xu, N. The antibacterial mechanism of carvacrol and thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef]
- Šernaitė, L. Plant Extracts: Antimicrobial and Antifungal Activity and Appliance in Plant Protection. Sodinink. Daržinink. 2017, 36, 58–68. [Google Scholar]
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial Properties and Mechanism of Action of Some Plant Extracts Against Food Pathogens and Spoilage Microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar] [CrossRef]
- Khoshbakht, T.; Karami, A.; Tahmasebi, A.; Maggi, F. The Variability of Thymol and Carvacrol Contents Reveals the Level of Antibacterial Activity of the Essential Oils from Different Accessions of Oliveria Decumbens. Antibiotics 2020, 9, 409. [Google Scholar] [CrossRef]
- Altundag, S.; Aslim, B.; Ozturk, S. In Vitro Antimicrobial Activities of Essential Oils from Origanum Minutiflorum and Sideritis Erytrantha Subsp. Erytrantha on Phytopathogenic Bacteria. J. Essent. Oil Res. 2011, 23, 4–8. [Google Scholar] [CrossRef]
- Iacobellis, N.S.; Lo Cantore, P.; Capasso, F.; Senatore, F. Antibacterial Activity of Cuminum cyminum, L. and Carum carvi, L. Essential Oils. J. Agric. Food Chem. 2005, 53, 57–61. [Google Scholar] [CrossRef]
- Lo Cantore, P.; Shanmugaiah, V.; Iacobellis, N.S. Antibacterial Activity of Essential Oil Components and Their Potential Use in Seed Disinfection. J. Agric. Food Chem. 2009, 57, 9454–9461. [Google Scholar] [CrossRef]
- Sotelo, J.P.; Oddino, C.; Giordano, D.F.; Carezzano, M.E.; Oliva, M.D.L.M. Effect of Thymus Vulgaris Essential Oil on Soybeans Seeds Infected with Pseudomonas Syringae. Physiol. Mol. Plant Pathol. 2021, 116, 101735. [Google Scholar] [CrossRef]
- Pawlowski, M.L.; Hartman, G.L. First Report of Curtobacterium Flaccumfaciens pv. Flaccumfaciens Causing Bacterial Tan Spot on Soybean in Africa. Plant Dis. 2019, 103, 2665. [Google Scholar] [CrossRef]
- Sammer, U.; Wagner, K. Curtobacterium Flaccumfaciens pv. Flaccumfaciens on Soybean in Germany—A Threat for Farming. J. Phytopathol. 2012, 160, 314–316. [Google Scholar] [CrossRef]
- Aysan, Y.; Horuz, S. Plant Pathogenic Bacteria Control through Seed Application. CRC Press: Boca Raton, FL, USA, 2015; pp. 323–332. ISBN 978-1-4822-4053-5. [Google Scholar]
- Arshad, Z.; Hanif, M.A.; Qadri, R.W.K.; Khan, M. Role of essential oils in plant diseases protection: A review. Int. J. Chem. Biol. Sci. 2014, 6, 11–17. [Google Scholar]
- Barbosa, L.N.; Rall, V.L.M.; Fernandes, A.A.H.; Ushimaru, P.I.; da Silva Probst, I.; Fernandes, A. Essential Oils Against Foodborne Pathogens and Spoilage Bacteria in Minced Meat. Foodborne Pathog. Dis. 2009, 6, 725–728. [Google Scholar] [CrossRef]
- Isaac, G.S.; Abu-Tahon, M.A. In Vitro Antifugal Activity of Medicinal Plant Extract against Fusarium Oxysporum f. Sp. Lycopersici Race 3 the Causal Agent of Tomato Wilt. Acta Biol. Hung. 2014, 65, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbega, E.R.; Mortensen, C.N.; Mabagala, R.B.; Wulff, E.G. The effect of plant extracts as seed treatments to control bacterial leaf spot of tomato in Tanzania. J. Gen. Plant Pathol. 2012, 78, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Riccioni, L.; Orzali, L.; Romani, M.; Annicchiarico, P.; Pecetti, L. Organic seed treatments with essential oils to control Ascochyta blight in pea. Eur. J. Plant Pathol. 2019, 155, 831–840. [Google Scholar] [CrossRef]
- Camele, I.; Elshafie, H.S.; Caputo, L.; de Feo, V. Anti-Quorum Sensing and Antimicrobial Effect of Mediterranean Plant Essential Oils Against Phytopathogenic Bacteria. Front. Microbiol. 2019, 10, 2619. [Google Scholar] [CrossRef] [Green Version]
- Habeck, M. Mistletoe Compound Enters Clinical Trials. Drug Discov. Today 2003, 8, 52–53. [Google Scholar] [CrossRef]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Ngegba, P.M.; Cui, G.; Khalid, M.Z.; Zhong, G. Use of Botanical Pesticides in Agriculture as an Alternative to Synthetic Pesticides. Agriculture 2022, 12, 600. [Google Scholar] [CrossRef]
- Flores, J.B.; García, J.O.; Becheleni, F.R.C.; Espinoza, A.V.; Wong-Corral, F.J.; Rueda-Puente, E.O. Effect of essential oils in the control of the Clavibacter michiganensis subespecie michiganensis in tomato (Lycopersicum esculentum L.) plants. Biotecnia 2018, 20, 96–101. [Google Scholar] [CrossRef]
- Dow, M.; Slater, H. DISEASES|Bacterial Diseases. In Encyclopedia of Applied Plant Sciences; Thomas, B., Ed.; Elsevier: Oxford, UK, 2003; pp. 212–223. ISBN 978-0-12-227050-5. [Google Scholar]
- Nabavi, S.F.; Di Lorenzo, A.; Izadi, M.; Sobarzo-Sánchez, E.; Daglia, M.; Nabavi, S.M. Antibacterial Effects of Cinnamon: From Farm to Food, Cosmetic and Pharmaceutical Industries. Nutrients 2015, 7, 7729–7748. [Google Scholar] [CrossRef]
- Kokkini, S.; Karousou, R.; Dardioti, A.; Krigas, N.; Lanaras, T. Autumn essential oils of Greek oregano. Phytochemistry 1997, 44, 8836. [Google Scholar] [CrossRef]
- Fedoseyeva, L.M.; Kerasheva, S.I.; Karabasova, E.V. Antimicrobial activity of dry extract from leaves of Bergenia crassifolia (L.) Fritsch for pathogens of some suppurative inflammatory diseases. Rastit. Resur. 2000, 36, 153–157. [Google Scholar]
- Kokoska, L.; Polesny, Z.; Rada, V.; Nepovim, A.; Vanek, T. Screening of Some Siberian Medicinal Plants for Antimicrobial Activity. J. Ethnopharmacol. 2002, 82, 51–53. [Google Scholar] [CrossRef]
- Marchese, A.; Arciola, C.R.; Barbieri, R.; Silva, A.S.; Nabavi, S.F.; Tsetegho Sokeng, A.J.; Izadi, M.; Jafari, N.J.; Suntar, I.; Daglia, M.; et al. Update on Monoterpenesf as Antimicrobial Agents: A Particular Focus on p-Cymene. Materials 2017, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A.; Barbieri, R.; Coppo, E.; Orhan, I.E.; Daglia, M.; Nabavi, S.F.; Izadi, M.; Abdollahi, M.; Nabavi, S.M.; Ajami, M. Antimicrobial Activity of Eugenol and Essential Oils Containing Eugenol: A Mechanistic Viewpoint. Crit. Rev. Microbiol. 2017, 43, 668–689. [Google Scholar] [CrossRef]
- Ryssel, H.; Kloeters, O.; Germann, G.; Schäfer, T.; Wiedemann, G.; Oehlbauer, M. The Antimicrobial Effect of Acetic Acid--an Alternative to Common Local Antiseptics? Burns 2009, 35, 695–700. [Google Scholar] [CrossRef]
- Han, C.; Shao, H.; Zhou, S.; Mei, Y.; Cheng, Z.; Huang, L.; Lv, G. Chemical Composition and Phytotoxicity of Essential Oil from Invasive Plant, Ambrosia Artemisiifolia L. Ecotoxicol. Environ. Saf. 2021, 211, 111879. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Werrie, P.-Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.-L. Phytotoxicity of Essential Oils: Opportunities and Constraints for the Development of Biopesticides. A Review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef]
- Rys, M.; Saja-Garbarz, D.; Skoczowski, A. Phytotoxic Effects of Selected Herbal Extracts on the Germination, Growth, and Metabolism of Mustard and Oilseed Rape. Agronomy 2022, 12, 110. [Google Scholar] [CrossRef]
- Arminante, F.; de Falco, E.; de Feo, V.; de Martino, L.; Mancini, E.; Quaranta, E. Allelopathic activity of essential oils from Mediterranean Labiatae. In Proceedings of the I International Symposium on the Labiatae: Advances in Production, Biotechnology and Utilisation 723, Sanremo, Italy, 22–25 February 2006; pp. 347–356. [Google Scholar]
- Dudai, N.; Poljakoff-Mayber, A.; Mayer, A.; Putievsky, E.; Lerner, H. Essential oils as allelochemicals and their potential use as bioherbicides. J. Chem. Ecol. 1999, 25, 1079–1089. [Google Scholar] [CrossRef]
- Gougoulias, N.; Papachatzis, A.; Vagelas, I.; Chouliaras, N. Greek Oregano Essential Oils Production, Phytotoxicity, and Antifungal Activity. Biotechnol. Biotechnol. Equip. 2009, 23, 1150–1152. [Google Scholar]
- Werrie, P.-Y.; Juillard, A.; Heintz, C.; Brisset, M.-N.; Fauconnier, M.-L. Phytotoxicity and Plant Defence Induction by Cinnamomum Cassia Essential Oil Application on Malus Domestica Tree: A Molecular Approach. Agronomy 2022, 12, 512. [Google Scholar] [CrossRef]
- Massimo, E. Maffei Monoterpenoid Plant-Plant Interactions Upon Herbivory. Curr. Bioact. Compd. 2012, 8, 65–70. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Spencer, G.F. Volatile Monoterpenes as Potential Parent Structures for New Herbicides. Weed Sci. 1993, 41, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Garb, S. Differential Growth-Inhibitors Produced by Plants. Bot. Rev. 1961, 27, 422–443. [Google Scholar] [CrossRef]
- Ullah, N.; Haq, I.U.; Mirza, B. Phytotoxicity evaluation and phytochemical analysis of three medicinally important plants from Pakistan. Toxicol. Ind. Health. 2015, 31, 389–395. [Google Scholar] [CrossRef]
- Koul, B.; Kumar, A.; Yadav, D.; Jin, J.-O. Bergenia Genus: Traditional Uses, Phytochemistry and Pharmacology. Molecules 2020, 25, 5555. [Google Scholar] [CrossRef]
- Osdaghi, E.; Young, A.J.; Harveson, R.M. Bacterial Wilt of Dry Beans Caused by Curtobacterium Flaccumfaciens pv. Flaccumfaciens: A New Threat from an Old Enemy. Mol. Plant Pathol. 2020, 21, 605–621. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Jurado, F.; Navarro-Cruz, A.R.; Ochoa-Velasco, C.E.; Palou, E.; López-Malo, A.; Ávila-Sosa, R. Essential Oils in Vapor Phase as Alternative Antimicrobials: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1641–1650. [Google Scholar] [CrossRef]
- Isman, M.; Machial, C.M. Pesticides Based on Plant Essential Oils: From Traditional Practice to Commercialization. Nat. Occur. Bioact. Compd. 2006, 3, 29–44. [Google Scholar]
- Devrnja, N.; Milutinović, M.; Savić, J. When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides. Sustainability 2022, 14, 6847. [Google Scholar] [CrossRef]
- Lammari, N.; Louaer, O.; Meniai, A.H.; Elaissari, A. Encapsulation of Essential Oils via Nanoprecipitation Process: Overview, Progress, Challenges, and Prospects. Pharmaceutics 2020, 12, 431. [Google Scholar] [CrossRef] [PubMed]
- Menossi, M.; Ollier, R.; Casalongué, C.; Alvarez, V. Essential Oils-Loaded Bio-Nanomaterials for Sustainable Agricultural Applications. J. Chem. Technol. Biotechnol. 2021, 96, 2109–2122. [Google Scholar] [CrossRef]
- Tarakanov, R.; Lukianova, A.; Pilik, R.; Evseev, P.; Miroshnikov, K.; Dzhalilov, F.; Ignatov, A. First report of Curtobacterium flaccumfaciens pv. flaccumfaciens causing a bacterial tan spot of soybean in Russia. Plant Dis. 2022, 103, 2665. [Google Scholar]
- Sarkar, S.F.; Guttman, D.S. Evolution of the Core Genome of Pseudomonas Syringae, a Highly Clonal, Endemic Plant Pathogen. Appl. Environ. Microbiol. 2004, 70, 1999–2012. [Google Scholar] [CrossRef] [Green Version]
- Evseev, P.; Lukianova, A.; Tarakanov, R.; Tokmakova, A.; Shneider, M.; Ignatov, A.; Miroshnikov, K. Curtobacterium Spp. and Curtobacterium Flaccumfaciens: Phylogeny, Genomics-Based Taxonomy, Pathogenicity, and Diagnostics. Curr. Issues Mol. Biol. 2022, 44, 889–927. [Google Scholar] [CrossRef]
- Tegli, S.; Sereni, A.; Surico, G. PCR-Based Assay for the Detection of Curtobacterium Flaccumfaciens pv. Flaccumfaciens in Bean Seeds. Lett Appl Microbiol 2002, 35, 331–337. [Google Scholar] [CrossRef]
- Heinrich, M. The Identification of Medicinal Plants. A Handbook of the Morphology of Botanicals in Commerce; Applequist, W., Ed.; American Botanical Council: Austin, TX, USA; Univ. Missouri Botanical Garden Press: St. Louis, MO, USA, 2007; p. 209. [Google Scholar]
- Radušienė, J.; Judžentienė, A.; Pečiulytė, D.; Janulis, V. Essential oil composition and antimicrobial assay of Acorus calamus leaves from different wild populations. Plant Genet. Resour. 2007, 5, 37–44. [Google Scholar] [CrossRef]
- Kumar, S.S.; Akram, A.; Ahmed, T.S.F.; Jaabir, M.S.M. Phytochemical Analysis and Antimicrobial Activity of the Ethanolic Extract of Acorus Calamus Rhizome. Orient. J. Chem. 2010, 26, 223–227. [Google Scholar]
- Reichling, J.; Schnitzler, P.; Suschke, U.; Saller, R. Essential Oils of Aromatic Plants with Antibacterial, Antifungal, Antiviral, and Cytotoxic Properties—An Overview. CMR 2009, 16, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanjari, A.; Bahonar, A.; Noori, N.; Siahkalmahaleh, M.R.; Rezaeigolestani, M.; Asgarian, Z.; Khanjari, J. In Vitro Antibacterial Activity of Pimpinella Anisum Essential Oil and Its Influence on Microbial, Chemical, and Sensorial Properties of Minced Beef during Refrigerated Storage. J. Food Saf. 2019, 39, e12626. [Google Scholar] [CrossRef]
- Benkeblia, N. Antimicrobial Activity of Essential Oil Extracts of Various Onions (Allium Cepa) and Garlic (Allium Sativum). LWT—Food Sci. Technol. 2004, 37, 263–268. [Google Scholar] [CrossRef]
- Gull, I.; Saeed, M.; Shaukat, H.; Aslam, S.M.; Samra, Z.Q.; Athar, A.M. Inhibitory Effect of Allium Sativum and Zingiber Officinale Extracts on Clinically Important Drug Resistant Pathogenic Bacteria. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential Oils: Sources of Antimicrobials and Food Preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef] [Green Version]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils—Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- França Orlanda, J.F.; Nascimento, A.R. Chemical Composition and Antibacterial Activity of Ruta Graveolens L. (Rutaceae) Volatile Oils, from São Luís, Maranhão, Brazil. S. Afr. J. Bot. 2015, 99, 103–106. [Google Scholar] [CrossRef]
- Moslemi, H.R.; Hoseinzadeh, H.; Badouei, M.A.; Kafshdouzan, K.; Fard, R.M.N. Antimicrobial Activity of Artemisia Absinthium Against Surgical Wounds Infected by Staphylococcus Aureus in a Rat Model. Indian J. Microbiol. 2012, 52, 601–604. [Google Scholar] [CrossRef] [Green Version]
- Chassagne, F.; Samarakoon, T.; Porras, G.; Lyles, J.T.; Dettweiler, M.; Marquez, L.; Salam, A.M.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. A Systematic Review of Plants With Antibacterial Activities: A Taxonomic and Phylogenetic Perspective. Front. Pharmacol. 2021, 11, 586548. [Google Scholar] [CrossRef]
- Pundarikakshudu, K.; Jayvadan, P.; Momin, M.; Deans, S.G. Anti-Bacterial Activity of Galega Officinalis L. (Goat’s Rue). J. Ethnopharmacol. 2001, 77, 111–112. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on Plant Antimicrobials: A Mechanistic Viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Naime, W.A.; Fahim, J.R.; Fouad, M.A.; Kamel, M.S. Antibacterial, Antifungal, and GC–MS Studies of Melissa Officinalis. S. Afr. J. Bot. 2019, 124, 228–234. [Google Scholar] [CrossRef]
- Budiman, A.; Aulifa, D.; Kusuma, A.; Sulastri, A. Antibacterial and Antioxidant Activity of Black Mulberry (Morus Nigra L.) Extract for Acne Treatment. Pharmacogn. J. 2017, 9, 611–614. [Google Scholar] [CrossRef]
- Patra, J.K.; Kim, E.S.; Oh, K.; Kim, H.-J.; Kim, Y.; Baek, K.-H. Antibacterial Effect of Crude Extract and Metabolites of Phytolacca Americana on Pathogens Responsible for Periodontal Inflammatory Diseases and Dental Caries. BMC Complement. Altern. Med. 2014, 14, 343. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, H.; Kawai, Y.; Sawano, R.; Kurihara, H.; Yamazaki, K.; Inoue, N. Antimicrobial Substances from Rhizomes of the Giant Knotweed Polygonum Sachalinense against the Fish Pathogen Photobacterium Damselae Subsp. Piscicida. Z. Nat. C 2005, 60, 39–44. [Google Scholar] [CrossRef]
- Wang, W.; Zu, Y.; Fu, Y.; Reichling, J.; Suschke, U.; Nokemper, S.; Zhang, Y. In Vitro Antioxidant, Antimicrobial and Anti-Herpes Simplex Virus Type 1 Activity of Phellodendron Amurense Rupr. from China. Am. J. Chin. Med. 2012, 37, 195–203. [Google Scholar] [CrossRef]
- Abdelbaky, A.S.; Mohamed, A.M.H.A.; Alharthi, S.S. Antioxidant and Antimicrobial Evaluation and Chemical Investigation of Rosa Gallica Var. Aegyptiaca Leaf Extracts. Molecules 2021, 26, 6498. [Google Scholar] [CrossRef]
- Kiss, T.; Cank, K.B.; Orbán-Gyapai, O.; Liktor-Busa, E.; Zomborszki, Z.P.; Rutkovska, S.; Pučka, I.; Németh, A.; Csupor, D. Phytochemical and Pharmacological Investigation of Spiraea Chamaedryfolia: A Contribution to the Chemotaxonomy of Spiraea Genus. BMC Res. Notes 2017, 10, 762. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, Y.; Song, Q.; Wang, F.; Sun, L.; Liu, L.; Yang, X.; Yi, J.; Bao, Y.; Ma, H.; et al. Anti-Biofilm Activities from Bergenia Crassifolia Leaves against Streptococcus Mutans. Front. Microbiol. 2017, 8, 1738. [Google Scholar] [CrossRef] [Green Version]
- Koffi-Nevry, R.; Kouassi, K.C.; Nanga, Z.Y.; Koussémon, M.; Loukou, G.Y. Antibacterial Activity of Two Bell Pepper Extracts: Capsicum Annuum L. and Capsicum Frutescens. Int. J. Food Prop. 2012, 15, 961–971. [Google Scholar] [CrossRef]
- Żurek, N.; Pawłowska, A.; Pycia, K.; Grabek-Lejko, D.; Kapusta, I.T. Phenolic Profile and Antioxidant, Antibacterial, and Antiproliferative Activity of Juglans Regia L. Male Flowers. Molecules 2022, 27, 2762. [Google Scholar] [CrossRef] [PubMed]
- Okla, M.K.; Alamri, S.A.; Salem, M.Z.M.; Ali, H.M.; Behiry, S.I.; Nasser, R.A.; Alaraidh, I.A.; Al-Ghtani, S.M.; Soufan, W. Yield, phytochemical constituents, and antibacterial activity of essential oils from the leaves/twigs, branches, branch wood, and branch bark of Sour Orange (Citrus aurantium L.). Processes 2019, 7, 363. [Google Scholar] [CrossRef] [Green Version]
- Abeysinghe, P.D. Antibacterial Activity of Some Medicinal Mangroves against Antibiotic Resistant Pathogenic Bacteria. Indian J. Pharm. Sci. 2010, 72, 167. [Google Scholar] [CrossRef] [PubMed]
- Clarice, P.; Mudzengi, A.M.; Musa, T.; Chrispen, M.; Burumu, J.V.; Tinyiko, H. Antibacterial activity of aqueous and methanol extracts of selected species used in livestock health management. Pharm. Biol. 2017, 55, 1054–1060. [Google Scholar]
- Burt, S.A. Essential oils: Their antibacterial properties and potential applications in foods: A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- CLSI Document M07-A10; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Approved Standard. 10th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015.
- ISTA. International Rules for Seed Testing. Seed Sci. Technol. 1999, 27, 178. [Google Scholar]
- Nalini, S.; Parthasarathi, R. Optimization of Rhamnolipid Biosurfactant Production from Serratia Rubidaea SNAU02 under Solid-State Fermentation and Its Biocontrol Efficacy against Fusarium Wilt of Eggplant. Ann. Agrar. Sci. 2018, 16, 108–115. [Google Scholar] [CrossRef]
- Baharum, S.N.; Bunawan, H.; Ghani, M.A.; Mustapha, W.A.W.; Noor, N.M. Analysis of the Chemical Composition of the Essential Oil of Polygonum Minus Huds. Using Two-Dimensional Gas Chromatography-Time-of-Flight Mass Spectrometry (GC-TOF MS). Molecules 2010, 15, 7006–7015. [Google Scholar] [CrossRef] [Green Version]
- Sibiya, M.; Sumbwanyambe, M. An Algorithm for Severity Estimation of Plant Leaf Diseases by the Use of Colour Threshold Image Segmentation and Fuzzy Logic Inference: A Proposed Algorithm to Update a “Leaf Doctor” Application. AgriEngineering 2019, 1, 15. [Google Scholar] [CrossRef]
- Hsieh, T.F.; Huang, H.C.; Mündel, H.-H.; Erickson, R.S. A rapid indoor technique for screening common bean (Phaseolus vulgaris L.) for resistance to bacterial wilt [Curtobacterium flaccumfaciens pv. flaccumfaciens (Hedges) Collins and Jones]. Rev. Mex. Fitopatol. 2003, 21, 370–374. [Google Scholar]
- Madden, L.V.; Hughes, G.; van den Bosch, F. The Study of Plant Disease Epidemics. American Phytopathological Society (APS Press): St. Paul, MN, USA, 2007; ISBN 978-0-89054-505-8. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| EO/PE/Antibiotic/Reference Pesticide | Bacteria (Strain) | |||
|---|---|---|---|---|
| PSG (G2) | CFF (F-125-1) | |||
| MIC | MBC | MIC | MBC | |
| gentamicin | 50 | 80 | 50 | 100 |
| thiram | 200 | 360 | 400 | 720 |
| Essential oils | ||||
| Cinnamomum aromaticum | 200 | 280 | 400 | 560 |
| Thymus vulgaris | 1200 | 1440 | 400 | 720 |
| Origanum vulgare | 1600 | 3200 | 200 | 280 |
| Mentha longifolia | 1600 | 2560 | 1200 | 1520 |
| Mentha piperita | 1600 | 3200 | 1600 | 3200 |
| Syzygium aromaticum | 1200 | 1600 | ND | ND |
| Lavandula angustifolia | 1600 | 3200 | 1600 | 3200 |
| Achillea millefolium | 1600 | 2880 | 1600 | 3200 |
| Allium sativum | ND | ND | >3200 | >3200 |
| Oleum calami | >3200 | >3200 | ND | ND |
| Citrus aurantiifolia | >3200 | >3200 | ND | ND |
| Elettaria cardamomum | >3200 | >3200 | ND | ND |
| Citrus reticulata | >3200 | >3200 | ND | ND |
| Pimpinella anisum | >3200 | >3200 | ND | ND |
| Foeniculum vulgare | >3200 | >3200 | ND | ND |
| Sālvia officinālis | ND | ND | >3200 | >3200 |
| Extracts | ||||
| Bergenia crassifolia ETH | 1000 | 4000 | 2500 | 5000 |
| Melissa officinalis ETH | 2500 | 5000 | ND | ND |
| Capsicum annuum ETH | 5000 | 9000 | ND | ND |
| Sambucus nigra ETH | 5000 | 10,000 | ND | ND |
| Phytolacca americana ETH | 5000 | 10,000 | ND | ND |
| Capsicum annuum W | 10,000 | 50,000 | 5000 | 9000 |
| Galega officinalis W | 10,000 | 50,000 | 10,000 | 50,000 |
| Artemisia absinthium ETH | 10,000 | 50,000 | ND | ND |
| Phellodendron amurense ETH | 50,000 | 100,000 | 50,000 | >100,000 |
| Rosa pendulina ETH | 100,000 | >100,000 | ND | ND |
| Chelidonium majus ETH | ND | ND | 5000 | 10,000 |
| Morus nigra ETH | ND | ND | 50,000 | >100,000 |
| Spiraea salicifolia W | ND | ND | 100,000 | >100,000 |
| Oleum calami ETH | ND | ND | 100,000 | >100,000 |
| Retention Time (min) | Compound | Area ± SD, % | |||
|---|---|---|---|---|---|
| a MSD | b FID | LBEE | CCEO | OEO | |
| 9.5 ± 0.03 | - | Acetic acid | 27.91 ± 0.12 | - | - |
| 11.54 ± 0.0 | - | Propanoic acid | 0.93 ± 0.09 | - | - |
| 14.21 ± 0.01 | - | 2,3-Butanediol | 0.63 ± 0.02 | - | - |
| 15.93 ± 0.0 | - | Isovaleric acid | 1.04 ± 0.06 | - | - |
| 16.26 ± 0.0 | - | 2-Methylbutyric acid | 0.13 ± 0.01 | - | - |
| 19.39 ± 0.0 | 18.48 ± 0.02 | 3-Thujene | - | - | 0.33 ± 0.018 |
| 19.67 ± 0.0 | 18.75 ± 0.0 | α-Pinene | - | - | 1.65 ± 0.015 |
| 19.68 ± 0.01 | 18.76 ± 0.0 | Benzaldehyde | - | 0.08 ± 0.002 | - |
| 20.05 ± 0.0 | 19.15 ± 0.01 | α-Fenchene | - | - | 0.06 ± 0.002 |
| 20.12 ± 0.0 | 19.22 ± 0.0 | Camphene | - | 0.08 ± 0.01 | 0.45 ± 0.013 |
| 20.33 ± 0.02 | - | Hexanoic acid | 6.91 ± 0.14 | - | - |
| 20.97 ± 0.01 | 20.12 ± 0.0 | β-Pinene | - | - | 0.46 ± 0.004 |
| 21.16 ± 0.03 | 20.36 ± 0.0 | β-Myrcene | - | - | 0.84 ± 0.009 |
| 21.52 ± 0.02 | 20.77 ± 0.0 | 2,6-Dimethyl 2,6-octadiene | - | - | 0.03 ± 0.0 |
| 21.67 ± 0.03 | 20.91 ± 0.01 | Pseudolimonen | - | - | 0.07 ± 0.004 |
| 22.07 ± 0.01 | 21.33 ± 0.0 | α-Terpinene | - | - | 0.53 ± 0.01 |
| 22.16 ± 0.0 | 21.41 ± 0.0 | Cymene | - | - | 19.85 ± 0.36 |
| 22.35 ± 0.0 | 21.61 ± 0.0 | p-1-Menthene | - | - | 0.035 ± 0.014 |
| 22.46 ± 0.0 | 21.73 ± 0.0 | Eucalyptol, Limonene | - | - | 0.28 ± 0.003 |
| 22.50 ± 0.03 | - | 5-Methyl-3-methylenedihydro-2(3H)-furanone | 20.32 ± 0.71 | - | - |
| 22.53 ± 0.0 | 21.80 ± 0.0 | Limonene | - | 0.03 ± 0.001 | - |
| 23.27 ± 0.0 | 22.61 ± 0.02 | γ-Terpinene | - | - | 4.85 ± 0.002 |
| 23.92 ± 0.0 | 23.30 ± 0.0 | D-Fenchone | - | - | 0.04 ± 0.0 |
| 24.17 ± 0.01 | 23.61 ± 0.0 | Linalool | - | - | 2.53 ± 0.045 |
| 24.32 ± 0.03 | 23.86 ± 0.0 | Phenylethyl Alcohol | 0.29 ± 0.001 | 0.15 ± 0.007 | - |
| 25.36 ± 0.01 | 24.87 ± 0.0 | d-Camphor | - | - | 0.072 ± 0.009 |
| 25.43 ± 0.0 | 24.96 ± 0.0 | 3-Phenylpropanal | - | 0.10 ± 0.026 | - |
| 25.63 ± 0.0 | - | l-Menthone | 1.69 ± 0.08 | - | - |
| 25.86 ± 0.0 | - | Isomenthone | 1.25 ± 0.01 | - | - |
| 25.87 ± 0.0 | 25.42 ± 0.01 | 4-Phenylbutanal | - | 0.09 ± 0.01 | - |
| 26.04 ± 0.02 | 25.61 ± 0.0 | endo-Borneol | - | 0.1 ± 0.032 | 0.10 ± 0.03 |
| 26.06 ± 0.0 | - | Isoborneol | 0.46 ± 0.003 | - | - |
| 26.29 ± 0.01 | 25.9 ± 0.01 | Terpinen-4-ol | - | - | 0.04 ± 0.001 |
| 26.31 ± 0.01 | - | trans-Sabinene hydrate | 0.53 ± 0.006 | - | - |
| 26.47 ± 0.0 | - | Methyl salicylate | 5.18 ± 0.08 | - | - |
| 26.97 ± 0.01 | - | 2-Hydroxycineole | 1.16 ± 0.08 | - | - |
| 27.27 ± 0.0 | 26.91 ± 0.01 | 2-Anisaldehyde | - | 0.31 ± 0.037 | - |
| 27.49 ± 0.01 | - | Pulegone | 4.46 ± 0.0 | - | - |
| 27.58 ± 0.01 | 27.32 ± 0.01 | Thymol methyl ether | - | - | 0.15 ± 0.08 |
| 27.76 ± 0.02 | - | Piperitone | 0.66 ± 0.07 | - | - |
| 27.88 ± 0.0 | 27.53 ± 0.0 | (E)-Cinnamaldehyde | - | 84.25 ± 3.452 | - |
| 28.34 ± 0.0 | 28.13 ± 0.0 | Thymol | 3.33 ± 0.02 | - | 3.51 ± 0.055 |
| 28.57 ± 0.0 | 28.37 ± 0.0 | Carvacrol | 0.45 ± 0.01 | - | 62.37 ± 0.01 |
| 29.60 ± 0.0 | - | Eugenol | 10.89 ± 0.61 | - | - |
| 30.35 ± 0.0 | 30.24 ± 0.0 | Cinnamaldehyde dimethyl acetal | - | 3.36 ± 0.011 | - |
| 30.54 ± 0.0 | 30.43 ± 0.0 | Copaene | - | 0.38 ± 0.007 | - |
| 30.76 ± 0.01 | 30.71 ± 0.0 | Coumarin | - | 0.38 ± 0.001 | - |
| 30.90 ± 0.0 | 30.83 ± 0.0 | Cinnamyl acetate | - | 2.34 ± 0.052 | - |
| 31.22 ± 0.0 | 31.18 ± 0.01 | Caryophyllene | - | - | 0.6 ± 0.002 |
| 31.84 ± 0.01 | 31.82 ± 0.0 | Curcumene | - | 0.15 ± 0.04 | - |
| 31.94 ± 0.02 | 31.93 ± 0.01 | γ-Amorphene | - | 0.17 ± 0.0 | - |
| 32.02 ± 0.0 | - | 3,5-Di-tert-butylphenol | 1.6 ± 0.03 | - | - |
| 32.09 ± 0.0 | 32.08 ± 0.01 | o-Methoxycinnamaldehyde | - | 6.93 ± 0.024 | - |
| 32.26 ± 0.0 | 32.25 ± 0.01 | α-Muurolene | - | 0.34 ± 0.01 | - |
| 33.28 ± 0.01 | - | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | 0.35 ± 0.01 | - | - |
| 33.32 ± 0.0 | 33.39 ± 0.01 | α-Farnesene | - | - | 0.07 ± 0.002 |
| 33.35 ± 0.01 | 33.42 ± 0.0 | Methoxycinnamaldehyde dimethyl acetal | - | 0.4 ± 0.026 | - |
| 33.38 ± 0.01 | 33.46 ± 0.0 | Caryophylene oxide | - | - | 0.63 ± 0.03 |
| - | - | Total identified | 90.17 | 99.64 | 99.55 |
| Common Name | Latin Name | Family | Part of the Plant | EO or PE | References |
|---|---|---|---|---|---|
| Sweet-flag | Oleum calami L. | Acoraceae | rhizomes | EO, PE | [85,86] |
| Common yarrow | Achillea millefolium L. | Asteraceae | inflorescences | EO | [46] |
| Fennel | Foeniculum vulgare Mill. | Apiaceae | fruit | EO | [87] |
| Anise | Pimpinella anisum L. | Apiaceae | fruit | EO | [88] |
| Tansy | Tanacetum vulgare L. | Asteraceae | leaves and inflorescences | EO | [87] |
| Garlic | Allium sativum L. | Amaryllidaceae | bulbs | EO, PE | [89,90] |
| Thyme | Thymus vulgaris L. | Lamiaceae | leaves and inflorescences | EO | [37] |
| Oregano | Origanum vulgare L. | Lamiaceae | leaves and stems | EO | [52] |
| Peppermint | Mentha piperita L. | Lamiaceae | flowers, leaves, stems | EO | [87] |
| Horse mint | Mentha longifolia (L.) Huds. | Lamiaceae | flowers, leaves, stems | EO | [91] |
| English lavender | Lavandula angustifolia Mill. | Lamiaceae | leaves and inflorescences | EO | [92] |
| Rosemary | Rosmarinus officinalis L. | Lamiaceae | leaves and inflorescences | EO | [87] |
| Sage | Salvia officinālis L. | Lamiaceae | leaves and flowers | EO | [87] |
| Chinese cinnamon | Cinnamomum aromaticum (L.) Presl | Lauraceae | leaves | EO | [54] |
| Clove | Syzygium aromaticum (L.) Merr. & Perry | Myrtaceae | flowers, leaves, stems | EO | [85] |
| Key lime | Citrus aurantiifolia (Christm.) Swingle | Rutaceae | fruit peel | EO | [85] |
| Mandarin orange | Citrus reticulata Blanco | Rutaceae | fruit peel | EO | [87] |
| Common rue | Ruta graveolens L. | Rutaceae | leaves and stems | EO | [93] |
| Green cardamom | Elettaria cardamomum (L.) Maton | Zingiberaceae | fruit | EO | [87] |
| Caraway | Carum carvi L. | Apiaceae | fruit | PE | [36] |
| Wormwood | Artemisia absinthium L. | Asteraceae | leaves and stems | PE | [94] |
| Elderberry | Sambucus nigra L. | Caprifoliaceae | leaves | PE | [95] |
| Galega | Galega officinalis L. | Fabaceae | leaves and stems | PE | [96] |
| Common oak | Quercus robur L. | Fagaceae | leaves | PE | [97] |
| Lemon balm | Melissa officinalis L. | Lamiaceae | leaves and stems | PE | [98] |
| Black mulberry | Morus nigra L. | Moraceae | leaves | PE | [99] |
| Greater celandine | Chelidonium majus L. | Papaveraceae | leaves and stems | PE | [100] |
| Plume poppy | Macleaya cordata (Willd.) R. Br. | Papaveraceae | leaves and stems | PE | [97] |
| Pokeweed | Phytolacca americana L. | Phytolaccaceae | leaves and stems | PE | [100] |
| Giant knotweed | Reynoutria sachalinensis (F.Schmidt) Nakai, | Polygonaceae | leaves and stems | PE | [101] |
| Amur cork tree | Phellodendron amurense Rupr. (1857) | Rutaceae | leaves | PE | [102] |
| Alpine rose | Rosa pendulina L. | Rosaceae | leaves | PE | [103] |
| Bridewort | Spiraea salicifolia L. | Rosaceae | leaves and stems | PE | [104] |
| Leather bergenia | Bergenia crassifolia (L.) Fritsch | Saxifragaceae | rhizomes | PE | [105] |
| Cayenne pepper | Capsicum annuum L. | Solanaceae | fruit | PE | [106] |
| Manchurian walnut | Juglans mandshurica Maxim. | Juglandaceae | leaves | PE | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarakanov, R.I.; Dzhalilov, F.S.-U. Using of Essential Oils and Plant Extracts against Pseudomonas savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens on Soybean. Plants 2022, 11, 2989. https://doi.org/10.3390/plants11212989
Tarakanov RI, Dzhalilov FS-U. Using of Essential Oils and Plant Extracts against Pseudomonas savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens on Soybean. Plants. 2022; 11(21):2989. https://doi.org/10.3390/plants11212989
Chicago/Turabian StyleTarakanov, Rashit I., and Fevzi S.-U. Dzhalilov. 2022. "Using of Essential Oils and Plant Extracts against Pseudomonas savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens on Soybean" Plants 11, no. 21: 2989. https://doi.org/10.3390/plants11212989
APA StyleTarakanov, R. I., & Dzhalilov, F. S. -U. (2022). Using of Essential Oils and Plant Extracts against Pseudomonas savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens on Soybean. Plants, 11(21), 2989. https://doi.org/10.3390/plants11212989