
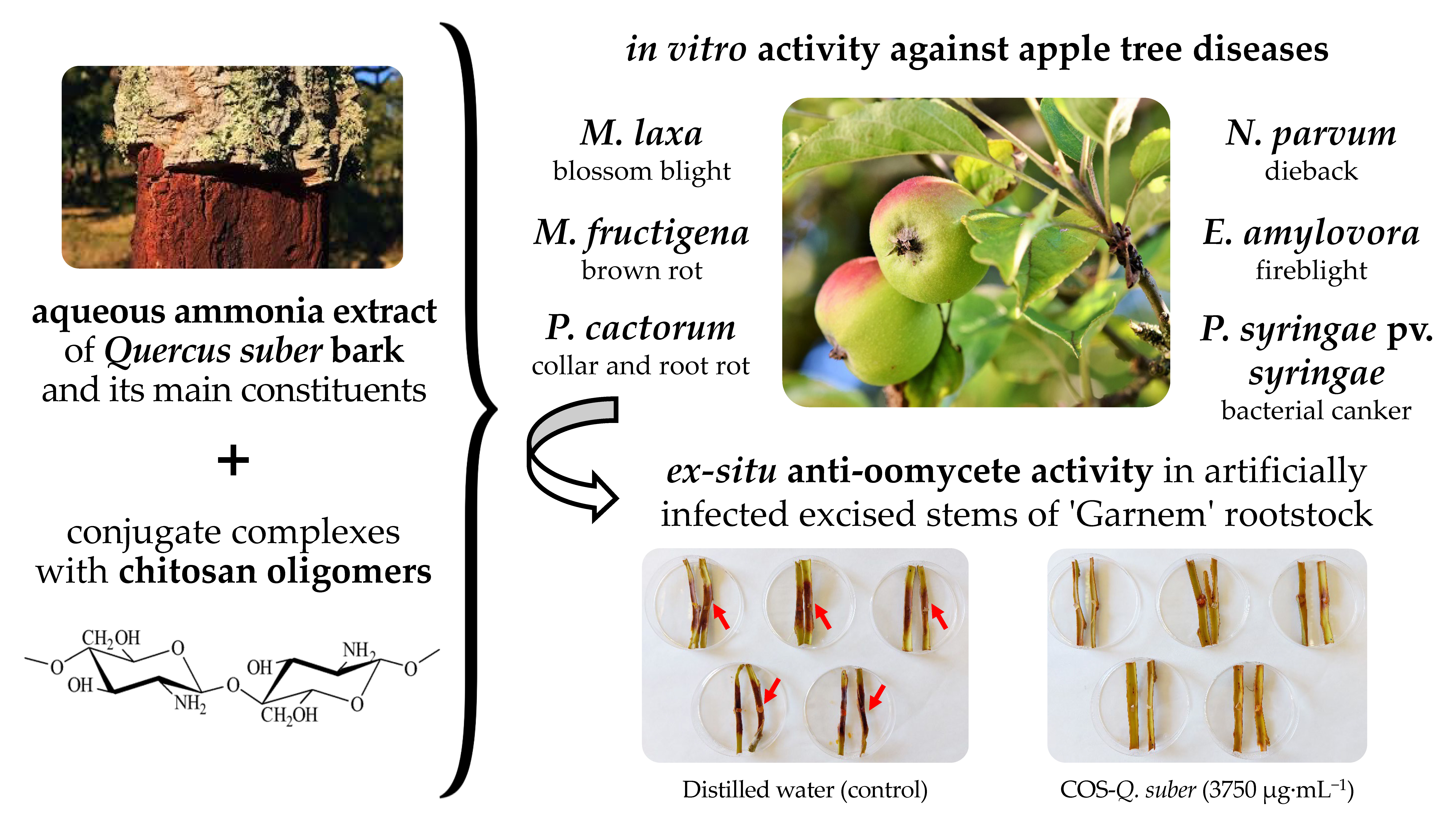
Valorization of Quercus suber L. Bark as a Source of Phytochemicals with Antimicrobial Activity against Apple Tree Diseases

 ,
, 
 ,
,  , and
, and 
Abstract
:
1. Introduction
2. Results
2.1. Identification of Phytochemicals by GC–MS
2.2. In Vitro Antimicrobial Activity Assessment
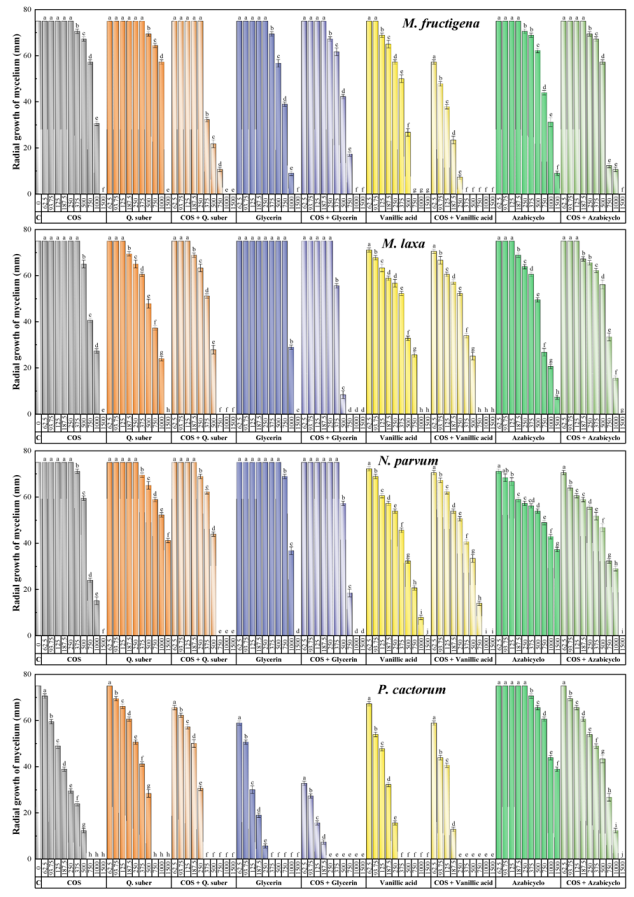
2.2.1. Antifungal and Anti-Oomycete Activity
2.2.2. In Vitro Antibacterial Assessment
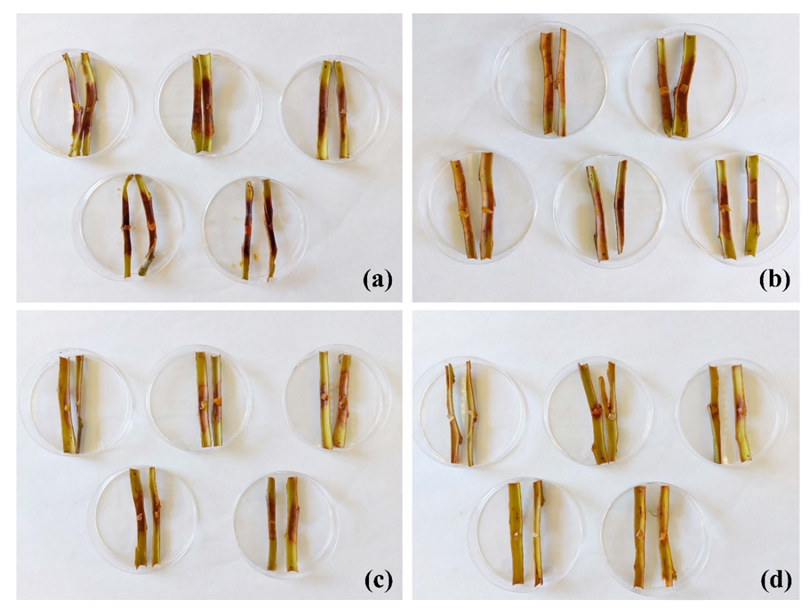
2.3. Protection of Excised Stems against P. cactorum
3. Discussion
3.1. The Phytochemical Profiles
3.2. Mode of Action
3.3. Antimicrobial Activity Comparison
3.3.1. Comparison with Antimicrobial Activities Reported for Other Q. suber Extracts
3.3.2. Comparison of Efficacy with Other Natural Compounds
3.3.3. Comparison of Efficacy with Conventional Fungicides
4. Materials and Methods
4.1. Vegetal Material
4.2. Reagents
4.3. Phytopathogen Isolates
4.4. Preparation of Bark Extracts, Chitosan Oligomers, and Conjugate Complexes
4.5. Extract and Conjugate Complexe Characterization
4.6. In Vitro Antimicrobial Activity Assessment
4.7. Protection Tests on Artificially Inoculated Excised Stems
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bugalho, M.N.; Caldeira, M.C.; Pereira, J.S.; Aronson, J.; Pausas, J.G. Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front. Ecol. Environ. 2011, 9, 278–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mota, S.; Pinto, C.; Cravo, S.; Rocha e Silva, J.; Afonso, C.; Sousa Lobo, J.M.; Tiritan, M.E.; Cidade, H.; Almeida, I.F. Quercus suber: A promising sustainable raw material for cosmetic application. Appl. Sci. 2022, 12, 4604. [Google Scholar] [CrossRef]
- Gonçalves, F.; Correia, P.; Silva, S.P.; Almeida-Aguiar, C.; Bowater, L. Evaluation of antimicrobial properties of cork. FEMS Microbiol. Lett. 2016, 363, fnv231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.P.; Sabino, M.A.; Fernandes, E.M.; Correlo, V.M.; Boesel, L.F.; Reis, R.L. Cork: Properties, capabilities and applications. Int. Mater. Rev. 2013, 50, 345–365. [Google Scholar] [CrossRef] [Green Version]
- Carriço, C.; Ribeiro, H.M.; Marto, J. Converting cork by-products to ecofriendly cork bioactive ingredients: Novel pharmaceutical and cosmetics applications. Ind. Crops Prod. 2018, 125, 72–84. [Google Scholar] [CrossRef]
- Touati, R.; Santos, S.A.O.; Rocha, S.M.; Belhamel, K.; Silvestre, A.J.D. The potential of cork from Quercus suber L. grown in Algeria as a source of bioactive lipophilic and phenolic compounds. Ind. Crops Prod. 2015, 76, 936–945. [Google Scholar] [CrossRef]
- Coquet, C.; Bauza, E.; Oberto, G.; Berghi, A.; Farnet, A.; Ferré, E.; Peyronel, D.; Dal Farra, C.; Domloge, N. Quercus suber cork extract displays a tensor and smoothing effect on human skin: An in vivo study. Drugs Under Exp. Clin. Res. 2005, 31, 89–99. [Google Scholar]
- Coquet, C.; Ferré, E.; Peyronel, D.; Dal Farra, C.; Farnet, A.M. Identification of new molecules extracted from Quercus suber L. cork. Comptes Rendus Biol. 2008, 331, 853–858. [Google Scholar] [CrossRef]
- Fernandes, A.; Sousa, A.; Mateus, N.; Cabral, M.; de Freitas, V. Analysis of phenolic compounds in cork from Quercus suber L. by HPLC–DAD/ESI–MS. Food Chem. 2011, 125, 1398–1405. [Google Scholar] [CrossRef]
- Pinto, J.; Oliveira, A.S.; Lopes, P.; Roseira, I.; Cabral, M.; Bastos, M.d.L.; Guedes de Pinho, P. Characterization of chemical compounds susceptible to be extracted from cork by the wine using GC-MS and 1H NMR metabolomic approaches. Food Chem. 2019, 271, 639–649. [Google Scholar] [CrossRef]
- Borrero, C.; Castillo, S.; Casanova, E.; Segarra, G.; Trillas, M.I.; Castaño, R.; Avilés, M. Capacity of composts made from agriculture industry residues to suppress different plant diseases. Acta Hortic. 2013, 1013, 459–463. [Google Scholar] [CrossRef]
- Garcia, H.; Ferreira, R.; Martins, C.; Sousa, A.F.; Freire, C.S.R.; Silvestre, A.J.D.; Kunz, W.; Rebelo, L.P.N.; Silva Pereira, C. Ex situ reconstitution of the plant biopolyester suberin as a film. Biomacromolecules 2014, 15, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Subhashini, S.; Begum, S.M.; Rajesh, G. Antimicrobial characterisation combining spectrophotometric analysis of different oak species. Int. J. Herb. Med. 2016, 4, 32–35. [Google Scholar]
- Rungjindamai, N.; Jeffries, P.; Xu, X.-M. Epidemiology and management of brown rot on stone fruit caused by Monilinia laxa. Eur. J. Plant Pathol. 2014, 140, 1–17. [Google Scholar] [CrossRef]
- Balsells-Llauradó, M.; Silva, C.J.; Usall, J.; Vall-llaura, N.; Serrano-Prieto, S.; Teixidó, N.; Mesquida-Pesci, S.D.; de Cal, A.; Blanco-Ulate, B.; Torres, R. Depicting the battle between nectarine and Monilinia laxa: The fruit developmental stage dictates the effectiveness of the host defenses and the pathogen’s infection strategies. Hortic. Res. 2020, 7, 167. [Google Scholar] [CrossRef]
- Delgado-Cerrone, L.; Mondino-Hintz, P.; Alaniz-Ferro, S. Botryosphariaceae species associated with stem canker, die-back and fruit rot on apple in Uruguay. Eur. J. Plant Pathol. 2016, 146, 637–655. [Google Scholar] [CrossRef]
- Di Francesco, A.; Rusin, C.; Di Foggia, M.; Marceddu, S.; Rombolà, A.; Botelho, R.V.; Baraldi, E. Characterization of apple cultivar susceptibility to Neofusicoccum parvum Brazilian strains. Eur. J. Plant Pathol. 2020, 156, 939–951. [Google Scholar] [CrossRef]
- Donat, V.; Biosca, E.G.; Peñalver, J.; López, M.M. Exploring diversity among Spanish strains of Erwinia amylovora and possible infection sources. J. Appl. Microbiol. 2007, 103, 1639–1649. [Google Scholar] [CrossRef]
- Peng, L.; Yang, S.; Zhang, Y.; Haseeb, H.; Song, S.; Xu, X.; Yang, M.; Zhang, J. Characterization and genetic diversity of Pseudomonas syringae pv. syringae isolates associated with rice bacterial leaf spot in heilongjiang, china. Biology 2022, 11, 720. [Google Scholar] [CrossRef]
- Usall, J.; Casals, C.; Sisquella, M.; Palou, L.; De Cal, A. Alternative technologies to control postharvest diseases of stone fruits. Stewart Postharvest Rev. 2015, 11, 1–6. [Google Scholar] [CrossRef]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; FHoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Lawal, O.; Bello, A.; Idem, R. The role of methyl diethanolamine (MDEA) in preventing the oxidative degradation of CO2 loaded and concentrated aqueous monoethanolamine (MEA)−MDEA blends during CO2 absorption from flue gases. Ind. Eng. Chem. Res. 2005, 44, 1874–1896. [Google Scholar] [CrossRef]
- Xie, H.-B.; Ma, F.; Wang, Y.; He, N.; Yu, Q.; Chen, J. Quantum chemical study on ·Cl-initiated atmospheric degradation of monoethanolamine. Environ. Sci. Technol. 2015, 49, 13246–13255. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Hoffmann, S.; Blomberg, L.G. Gas chromatographic—mass spectrometric analysis of compounds generated upon thermal degradation of some stationary phases in gas chromatography—Part II. J. High. Resolut. Chromatogr. 1985, 8, 734–740. [Google Scholar] [CrossRef]
- Michal, H.; Blanka, K.; Miloš, K. Chiral and nonchiral GC×GC/TOFMS analysis of natural compounds: The case of possible aggregation pheromones of chinese bark beetles Ips shangrila and Ips nitidus. In Gas Chromatography in Plant Science, Wine Technology, Toxicology and Some Specific Applications; Bekir, S., Çelikbıçak, O., Eds.; IntechOpen: Rijeka, Croatia, 2012; pp. 325–346. [Google Scholar] [CrossRef]
- Cunha, M.; Lourenço, A.; Barreiros, S.; Paiva, A.; Simões, P. Valorization of cork using subcritical water. Molecules 2020, 25, 4695. [Google Scholar] [CrossRef]
- Bouras, M.; Chadni, M.; Barba, F.J.; Grimi, N.; Bals, O.; Vorobiev, E. Optimization of microwave-assisted extraction of polyphenols from Quercus bark. Ind. Crops Prod. 2015, 77, 590–601. [Google Scholar] [CrossRef]
- Batista, M.; Rosete, M.; Ferreira, I.; Ferreira, J.; Duarte, C.; Matias, A.; Poejo, J.; Crespo, J.; Valério, R.; Fraga, M. Extracto Hidro-Glicólico de Cortiça, Processo para a Sua Preparação, Formulações Compreendendo o Referido Extracto e Sua Utilização. WO2015152746A1, 1 April 2014. [Google Scholar]
- Sánchez-Hernández, E.; Balduque-Gil, J.; Barriuso-Vargas, J.J.; Casanova-Gascón, J.; González-García, V.; Cuchí-Oterino, J.A.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Holm oak (Quercus ilex subsp. ballota (Desf.) Samp.) bark aqueous ammonia extract for the control of invasive forest pathogens. Int. J. Mol. Sci. 2022, 23, 11882. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Martín-Ramos, P.; Martín-Gil, J.; Santiago-Aliste, A.; Hernández-Navarro, S.; Oliveira, R.; González-García, V. Bark extract of Uncaria tomentosa L. for the control of strawberry phytopathogens. Horticulturae 2022, 8, 672. [Google Scholar] [CrossRef]
- Canonico, P.G.; Jahrling, P.B.; Pannier, W.L. Antiviral efficacy of pyrazofurin against selected rna viruses. Antivir. Res. 1982, 2, 331–337. [Google Scholar] [CrossRef]
- Kumar, V.; Bhatt, V.; Kumar, N. Amides from Plants: Structures and Biological Importance; Atta-ur-Rahman, F.R.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 56, pp. 287–333. [Google Scholar]
- Pan, Y.-j. A novel lactam from Delphinium caeruleum. J. Zhejiang Univ. Sci. A 2000, 1, 186–187. [Google Scholar] [CrossRef]
- Lesage-Meessen, L.; Delattre, M.; Haon, M.; Thibault, J.-F.; Ceccaldi, B.C.; Brunerie, P.; Asther, M. A two-step bioconversion process for vanillin production from ferulic acid combining Aspergillus niger and Pycnoporus cinnabarinus. J. Biotechnol. 1996, 50, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.W.; Tschaplinski, T.J.; Wang, L.; Glazebrook, J.; Greenberg, J.T. Priming in systemic plant immunity. Science 2009, 324, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Hojo, Y.; Saito, Y.; Tanimoto, T.; Hoefen, R.J.; Baines, C.P.; Yamamoto, K.; Haendeler, J.; Asmis, R.; Berk, B.C. Fluid shear stress attenuates hydrogen peroxide-induced c-Jun NH2-terminal kinase activation via a glutathione reductase-mediated mechanism. Circ. Res. 2002, 91, 712–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenther, A.; Zimmerman, P.; Wildermuth, M. Natural volatile organic compound emission rate estimates for U.S. woodland landscapes. Atmos. Environ. 1994, 28, 1197–1210. [Google Scholar] [CrossRef] [Green Version]
- Mudd, S.H. The mechanism of the enzymatic cleavage of S-adenosylmethionine to α-amino-γ-butyrolactone. J. Biol. Chem. 1959, 234, 1784–1786. [Google Scholar] [CrossRef]
- Stout, E.I.; McKessor, A. Glycerin-based hydrogel for infection control. Adv. Wound Care 2012, 1, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Mackie, D.P. The Euro Skin Bank: Development and application of glycerol-preserved allografts. J. Burn Care Rehabil. 1997, 18, s7–s9. [Google Scholar] [CrossRef]
- Linser, A. Glycerine as Fungicide or Bactericide Active. Substance. Patent WO/2002/069708, 12 September 2002. [Google Scholar]
- Jamilah, J.; Sharifa, A.; Sharifah, N. GC-MS analysis of various extracts from leaf of Plantago major used as traditional medicine. World Appl. Sci. J. 2012, 17, 67–70. [Google Scholar]
- Prabhadevi, V.; Sahaya, S.S.; Johnson, M.; Venkatramani, B.; Janakiraman, N. Phytochemical studies on Allamanda cathartica L. using GC–MS. Asian Pac. J. Trop. Biomed. 2012, 2, S550–S554. [Google Scholar] [CrossRef]
- Jatin, R.R.; Priya, R.S. Determination of bioactive components of Cynodon dactylon by GC-MS analysis & its in vitro antimicrobial activity. Int. J. Pharm. Life Sci. 2016, 7, 4880–4885. [Google Scholar]
- Hameed, R.H.; Mohammed, G.J.; Hameed, I.H. Characterization of antimicrobial metabolites produced by Salvadora persica and analysis of its chemical compounds using GC-MS and FTIR. Indian J. Public Health Res. Dev. 2018, 9, 241. [Google Scholar] [CrossRef]
- Maria, K.K.; Joanna, G.K.; Katerina, A.D.; Leland, G.K. Chemical composition and antibacterial activity against Escherichia coli of extracts of a common household plant. J. Med. Plants Res. 2021, 15, 56–63. [Google Scholar] [CrossRef]
- Kaur, J.; Gulati, M.; Singh, S.K.; Kuppusamy, G.; Kapoor, B.; Mishra, V.; Gupta, S.; Arshad, M.F.; Porwal, O.; Jha, N.K.; et al. Discovering multifaceted role of vanillic acid beyond flavours: Nutraceutical and therapeutic potential. Trends Food Sci. Technol. 2022, 122, 187–200. [Google Scholar] [CrossRef]
- Li, Y.; Cai, C.; Zeng, Q.; Zhang, J.; Liu, M.; Sun, Z.; Wang, T.; Yang, M.I.N.; Qian, W. Antibacterial mechanism of vanillic acid on physiological, morphological, and biofilm properties of carbapenem-resistant Enterobacter hormaechei. J. Food Prot. 2020, 83, 576–583. [Google Scholar] [CrossRef]
- Delaquis, P.; Stanich, K.; Toivonen, P. Effect of pH on the inhibition of Listeria spp. by vanillin and vanillic acid. J. Food Prot. 2005, 68, 1472–1476. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Kim, M.-C.; Kim, J.-S.; Balasundaram, C.; Jawahar, S.; Heo, M.-S. Identification and antimicrobial activity of combined extract from Azadirachta indica and Ocimum sanctum. Isr. J. Aquac.-Bamidgeh 2010, 62, 85–95. [Google Scholar] [CrossRef]
- Mudhafar, M.; Zainol, I.; Jaafar, C.; Alsailawi, H.; Majhool, A.A.; Alsaady, M. Phytochemical screening and characterization of Melia dubia leaves extract for antimicrobial activity against Escherichia coli and Staphylococcus aureus. Indian J. Ecol. 2020, 47, 493–496. [Google Scholar]
- Suryowati, T.; Sirait, R.H.; Siagian, F.E.; Nursyam, M. Bioactive compound impacting the metabolism and antibacterial activity of gadung tuber (Dioscorea hispida Dennst). J. Phys. Conf. Ser. 2020, 1665, 012030. [Google Scholar] [CrossRef]
- Akroum, S. Antifungal activity of acetone extracts from Punica granatum L., Quercus suber L. and Vicia faba L. J. Mycol. Médicale 2017, 27, 83–89. [Google Scholar] [CrossRef]
- Popović, B.M.; Štajner, D.; Ždero, R.; Orlović, S.; Galić, Z. Antioxidant characterization of oak extracts combining spectrophotometric assays and chemometrics. Sci. World J. 2013, 2013, 134656. [Google Scholar] [CrossRef] [Green Version]
- Lahlimi-Alami, Q.; Layachi, R.; Hassikou, R.; Benjelloun, J.; Amallah, L.; Guennoun, N.; Zaid, Y.; Bouzroud, S. Anticandidosic activity and acute toxicity of Quercus suber L. bark extracts. J. Med. Chem. Sci. 2022, 5, 769–778. [Google Scholar] [CrossRef]
- Hassikou, R.; Oulladi, H.; Arahou, M. Activité antimycosique des extraits du chêne-liège Quercus suber sur Trichophyton rubrum et Candida albicans. Phytothérapie 2014, 12, 206–212. [Google Scholar] [CrossRef]
- Akroum, S.; Rouibah, M. Utilisation d’extraits méthanoliques de plantes pour la protection des cultures de tomates-cerises (Solanum lycopersicum var. cerasiforme) contre l’infection fongique par Alternaria alternata. Biol. Aujourd’hui 2020, 214, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Morales, D. Oak trees (Quercus spp.) as a source of extracts with biological activities: A narrative review. Trends Food Sci. Technol. 2021, 109, 116–125. [Google Scholar] [CrossRef]
- Onaran, A.; Yanar, Y. In vivo and in vitro antifungal activities of five plant extracts against various plant pathogens. Egypt. J. Biol. Pest Control 2016, 26, 405–411. [Google Scholar]
- El Khetabi, A.; Lahlali, R.; Askarne, L.; Ezrari, S.; El Ghadaroui, L.; Tahiri, A.; Hrustić, J.; Amiri, S. Efficacy assessment of pomegranate peel aqueous extract for brown rot (Monilinia spp.) disease control. Physiol. Mol. Plant Pathol. 2020, 110, 101482. [Google Scholar] [CrossRef]
- Mamoci, E.; Cavoski, I.; Simeone, V.; Mondelli, D.; Al-Bitar, L.; Caboni, P. Chemical composition and in vitro activity of plant extracts from Ferula communis and Dittrichia viscosa against postharvest fungi. Molecules 2011, 16, 2609–2625. [Google Scholar] [CrossRef] [Green Version]
- El Khetabi, A.; Ezrari, S.; El Ghadraoui, L.; Tahiri, A.; Ait Haddou, L.; Belabess, Z.; Merah, O.; Lahlali, R. In vitro and in vivo antifungal activities of nine commercial essential oils against brown rot in apples. Horticulturae 2021, 7, 545. [Google Scholar] [CrossRef]
- Andreu, V.; Levert, A.; Amiot, A.; Cousin, A.; Aveline, N.; Bertrand, C. Chemical composition and antifungal activity of plant extracts traditionally used in organic and biodynamic farming. Environ. Sci. Pollut. Res. 2018, 25, 29971–29982. [Google Scholar] [CrossRef]
- Langa-Lomba, N.; Buzón-Durán, L.; Martín-Ramos, P.; Casanova-Gascón, J.; Martín-Gil, J.; Sánchez-Hernández, E.; González-García, V. Assessment of conjugate complexes of chitosan and Urtica dioica or Equisetum arvense extracts for the control of grapevine trunk pathogens. Agronomy 2021, 11, 976. [Google Scholar] [CrossRef]
- Langa-Lomba, N.; Buzón-Durán, L.; Sánchez-Hernández, E.; Martín-Ramos, P.; Casanova-Gascón, J.; Martín-Gil, J.; González-García, V. Antifungal activity against Botryosphaeriaceae fungi of the hydro-methanolic extract of Silybum marianum capitula conjugated with stevioside. Plants 2021, 10, 1363. [Google Scholar] [CrossRef] [PubMed]
- Langa-Lomba, N.; Sánchez-Hernández, E.; Buzón-Durán, L.; González-García, V.; Casanova-Gascón, J.; Martín-Gil, J.; Martín-Ramos, P. Activity of anthracenediones and flavoring phenols in hydromethanolic extracts of Rubia tinctorum against grapevine phytopathogenic fungi. Plants 2021, 10, 1527. [Google Scholar] [CrossRef] [PubMed]
- Oladejo, O.; Imani, J. Inhibitory effect of CUSTOS, a formulated Allium-based extract, on the growth of some selected plant pathogens. Int. J. Plant Biol. 2022, 13, 44–54. [Google Scholar] [CrossRef]
- Minova, S.; Sešķēna, R.; Voitkāne, S.; Metla, Z.; Daugavietis, M.; Jankevica, L. Impact of pine (Pinus sylvestris L.) and spruce (Picea abies (L.) Karst.) bark extracts on important strawberry pathogens. Proc. Latv. Acad. Sci. Sect. B. Nat. Exact Appl. Sci. 2015, 69, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-S.; Kim, J.; Shin, S.-C.; Lee, S.-G.; Park, I.-K. Antifungal activity of Myrtaceae essential oils and their components against three phytopathogenic fungi. Flavour Fragr. J. 2008, 23, 23–28. [Google Scholar] [CrossRef]
- Kim, J.; Lee, Y.S.; Lee, S.G.; Shin, S.C.; Park, I.K. Fumigant antifungal activity of plant essential oils and components from West Indian bay (Pimenta racemosa) and thyme (Thymus vulgaris) oils against two phytopathogenic fungi. Flavour Fragr. J. 2008, 23, 272–277. [Google Scholar] [CrossRef]
- Fontana, R.; Macchi, G.; Caproni, A.; Sicurella, M.; Buratto, M.; Salvatori, F.; Pappadà, M.; Manfredini, S.; Baldisserotto, A.; Marconi, P. Control of Erwinia amylovora growth by Moringa oleifera leaf extracts: In vitro and in planta effects. Plants 2022, 11, 957. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Buzón-Durán, L.; Cuchí-Oterino, J.A.; Martín-Gil, J.; Lorenzo-Vidal, B.; Martín-Ramos, P. Dwarf pomegranate (Punica granatum L. var. nana): Source of 5-HMF and bioactive compounds with applications in the protection of woody crops. Plants 2022, 11, 550. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Buzón-Durán, L.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Physicochemical characterization and antimicrobial activity against Erwinia amylovora, Erwinia vitivora, and Diplodia seriata of a light purple Hibiscus syriacus L. cultivar. Plants 2021, 10, 1876. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Buzón-Durán, L.; Langa-Lomba, N.; Casanova-Gascón, J.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Characterization and antimicrobial activity of a halophyte from the Asturian coast (Spain): Limonium binervosum (G.E.Sm.) C.E.Salmon. Plants 2021, 10, 1852. [Google Scholar] [CrossRef]
- Shabani, B.; Rezaei, R.; Charehgani, H.; Salehi, A. Study on antibacterial effect of essential oils of six plant species against Pseudomonas syringae pv. syringae Van Hall 1902 and Pseudomonas fluorescens Migula 1894. J. Plant Pathol. 2019, 101, 671–675. [Google Scholar] [CrossRef]
- Islam, M.S.; Sultana, R.; Hasan, M.A.; Alam, M.S.; Sikdar, B.; Kamaruzzaman, M.; Islam, M.A. Characterization and biocontrol measures of Pseudomonas syringae pv. syringae associated with citrus blast disease. Vegetos 2020, 33, 555–569. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Langa-Lomba, N.; González-García, V.; Casanova-Gascón, J.; Martín-Gil, J.; Santiago-Aliste, A.; Torres-Sánchez, S.; Martín-Ramos, P. Lignin–chitosan nanocarriers for the delivery of bioactive natural products against wood-decay phytopathogens. Agronomy 2022, 12, 461. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Fernandez-Arrojo, L.; Mengibar, M.; Belmonte-Reche, E.; Peñalver, P.; Acosta, F.N.; Ballesteros, A.O.; Morales, J.C.; Kidibule, P.; Fernandez-Lobato, M.; et al. Enzymatic production of fully deacetylated chitooligosaccharides and their neuroprotective and anti-inflammatory properties. Biocatal. Biotransform. 2017, 36, 57–67. [Google Scholar] [CrossRef]
- Sannan, T.; Kurita, K.; Iwakura, Y. Studies on chitin, 2. Effect of deacetylation on solubility. Die Makromol. Chem. 1976, 177, 3589–3600. [Google Scholar] [CrossRef]
- Yang, Y.; Shu, R.; Shao, J.; Xu, G.; Gu, X. Radical scavenging activity of chitooligosaccharide with different molecular weights. Eur. Food Res. Technol. 2005, 222, 36–40. [Google Scholar] [CrossRef]
- Maghami, G.G.; Roberts, G.A.F. Evaluation of the viscometric constants for chitosan. Die Makromol. Chem. 1988, 189, 195–200. [Google Scholar] [CrossRef]
- Tian, M.; Tan, H.; Li, H.; You, C. Molecular weight dependence of structure and properties of chitosan oligomers. RSC Adv. 2015, 5, 69445–69452. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corp.: Carol Stream, IL, USA, 2007; p. 804. [Google Scholar]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST). Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef] [Green Version]
- Levy, Y.; Benderly, M.; Cohen, Y.; Gisi, U.; Bassand, D. The joint action of fungicides in mixtures: Comparison of two methods for synergy calculation. EPPO Bull. 1986, 16, 651–657. [Google Scholar] [CrossRef]
- CLSI. CLSI standard M07—Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Matheron, M.E. Seasonal variation in susceptibility of Juglans hindsii and Paradox rootstocks of English walnut trees to Phytophthora citricola. Phytopathology 1985, 75, 970. [Google Scholar] [CrossRef]
- Álvarez Bernaola, L.A. Estudios de Etiología, Epidemiología y Control de un Nuevo Síndrome de Lesiones en Tronco y Ramas Principales de Cítricos Asociado a Phytophthora; Universitat Politècnica de València: Valencia, Spain, 2008. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Nehad, M.G.; Abdulrahaman, S.H. Antimicrobial efficacy of Casuarina equisetifolia extracts against some pathogenic microorganisms. J. Med. Plants Res. 2012, 6, 5819–5825. [Google Scholar]
- Naz, S.; Ahmad, S.; Ajaz Rasool, S.; Asad Sayeed, S.; Siddiqi, R. Antibacterial activity directed isolation of compounds from Onosma hispidum. Microbiol. Res. 2006, 161, 43–48. [Google Scholar] [CrossRef]
- Ouerghemmi, I.; Bettaieb Rebey, I.; Rahali, F.Z.; Bourgou, S.; Pistelli, L.; Ksouri, R.; Marzouk, B.; Saidani Tounsi, M. Antioxidant and antimicrobial phenolic compounds from extracts of cultivated and wild-grown Tunisian Ruta chalepensis. J. Food Drug Anal. 2017, 25, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ćurković-Perica, M.; Hrenović, J.; Kugler, N.; Goić-Barišić, I.; Tkalec, M. Antibacterial activity of Pinus pinaster bark extract and its components against multidrug-resistant clinical isolates of Acinetobacter baumannii. Croat. Chem. Acta 2015, 88, 133–137. [Google Scholar] [CrossRef]
- Salamone, A.; Zizzo, G.V.; Scarito, G. The antimicrobial activity of water extracts from Labiatae. Acta Hortic. 2006, 465–470. [Google Scholar] [CrossRef]
- Carezzano, M.E.; Sotelo, J.P.; Primo, E.; Reinoso, E.B.; Paletti Rovey, M.F.; Demo, M.S.; Giordano, W.F.; Oliva, M.D.L.M.; Flemetakis, E. Inhibitory effect of Thymus vulgaris and Origanum vulgare essential oils on virulence factors of phytopathogenic Pseudomonas syringae strains. Plant Biol. 2017, 19, 599–607. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Effective Concentration | M. fructigena | M. laxa | N. parvum | P. cactorum |
|---|---|---|---|---|---|
| COS | EC50 | 940.9 | 808.0 | 677.5 | 200.8 |
| EC90 | 1356.0 | 1342.7 | 1198.2 | 592.8 | |
| Q. suber bark | EC50 | 1073.6 | 684.4 | 1738.7 | 409.4 |
| EC90 | 1466.8 | 1360.4 | 3449.4 | 674.9 | |
| COS–Q. suber bark | EC50 | 314.2 | 446.4 | 508.0 | 224.4 |
| EC90 | 801.1 | 671.8 | 706.3 | 339.7 | |
| Glycerin | EC50 | 773.5 | 1012.0 | 964.3 | 115.5 |
| EC90 | 1216.1 | 1343.8 | 1470.8 | 267.1 | |
| COS–Glycerin | EC50 | 545.7 | 420.5 | 622.2 | 54.5 |
| EC90 | 880.6 | 623.2 | 890.7 | 185.1 | |
| Vanillic acid | EC50 | 440.2 | 498.8 | 448.0 | 159.3 |
| EC90 | 668.8 | 931.0 | 1108.7 | 313.1 | |
| COS–Vanillic acid | EC50 | 128.5 | 351.3 | 424.0 | 129.2 |
| EC90 | 256.1 | 669.9 | 861.6 | 214.5 | |
| Azabicyclo | EC50 | 862.0 | 636.0 | 1444.9 | 1677.1 |
| EC90 | 1556.3 | 1420.7 | 8779.8 | 6805.6 | |
| COS–Azabicyclo | EC50 | 606.5 | 696.8 | 667.4 | 567.7 |
| EC90 | 1117.1 | 1244.0 | 1399.9 | 1198.9 |
| Treatment | Effective Concentration | M. fructigena | M. laxa | N. parvum | P. cactorum |
|---|---|---|---|---|---|
| COS–Q. suber bark | EC50 | 3.19 | 1.66 | 1.92 | 1.20 |
| EC90 | 1.76 | 2.01 | 2.52 | 1.86 | |
| COS–Glycerin | EC50 | 1.56 | 2.14 | 1.28 | 2.69 |
| EC90 | 1.46 | 2.16 | 1.48 | 1.99 | |
| COS–Vanillic acid | EC50 | 6.08 | 1.76 | 1.27 | 1.38 |
| EC90 | 2.60 | 1.64 | 1.34 | 1.91 | |
| COS–Azabicyclo | EC50 | 1.48 | 1.02 | 1.38 | 0.63 |
| EC90 | 1.30 | 1.11 | 1.51 | 0.91 |
| Commercial Fungicide | Pathogen | Radial Growth of Mycelium (mm) | Inhibition (%) | ||||
|---|---|---|---|---|---|---|---|
| Rd/10 | Rd * | Rd × 10 | Rd/10 | Rd * | Rd × 10 | ||
| Azoxystrobin | M. laxa | 33.7 | 30.3 | 29.2 | 55 | 59.6 | 61.1 |
| M. fructigena | 60.3 | 56 | 33.2 | 19.6 | 25.3 | 55.8 | |
| N. parvum | 74.4 | 72.5 | 68.1 | 0.9 | 3.4 | 9.2 | |
| P. cactorum | 6 | 0 | 0 | 92 | 100 | 100 | |
| Mancozeb | M. laxa | 0 | 0 | 0 | 100 | 100 | 100 |
| M. fructigena | 0 | 0 | 0 | 100 | 100 | 100 | |
| N. parvum | 0 | 0 | 0 | 100 | 100 | 100 | |
| P. cactorum | 0 | 0 | 0 | 100 | 100 | 100 | |
| Fosetyl-Al | M. laxa | 72.1 | 13.3 | 0 | 3.9 | 82.2 | 100 |
| M. fructigena | 82.4 | 18.4 | 0 | 0 | 75.5 | 100 | |
| N. parvum | 59.1 | 8.7 | 0 | 21.2 | 88.4 | 100 | |
| P. cactorum | 64 | 0 | 0 | 14.7 | 100 | 100 | |
| Pathogen | Compound | Concentration (μg·mL−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 62.5 | 93.75 | 125 | 187.5 | 250 | 375 | 500 | 750 | 1000 | 1500 | ||
| E. amylovora | COS | + | + | + | + | + | + | + | + | + | − |
| Q. suber bark | + | + | + | + | + | + | + | + | − | − | |
| Glycerin | + | + | + | + | + | + | − | − | − | − | |
| Vanillic acid | + | + | + | + | + | + | + | − | − | − | |
| Azabicyclo | + | + | + | + | + | + | + | + | + | + | |
| COS−Q. suber bark | + | + | + | + | + | + | + | − | − | − | |
| COS−Glycerin | + | + | + | + | + | − | − | − | − | − | |
| COS−Vanillic acid | + | + | + | + | + | + | − | − | − | − | |
| COS−Azabicyclo | + | + | + | + | + | + | + | − | − | − | |
| P. syringae pv. syringae | COS | + | + | + | + | + | + | + | + | − | − |
| Q. suber bark | + | + | + | + | + | + | + | − | − | − | |
| Glycerin | + | + | + | + | + | + | + | − | − | − | |
| Vanillic acid | + | + | + | + | + | + | + | − | − | − | |
| Azabicyclo | + | + | + | + | + | + | + | + | + | + | |
| COS−Q. suber bark | + | + | + | + | + | − | − | − | − | − | |
| COS−Glycerin | + | + | + | + | + | − | − | − | − | − | |
| COS−Vanillic acid | + | + | + | + | + | + | − | − | − | − | |
| COS−Azabicyclo | + | + | + | + | + | + | + | − | − | − | |
| Sample | Mean of Ranks | Groups | ||
|---|---|---|---|---|
| MIC × 10 | 5.6 | A | ||
| MIC × 5 | 18.1 | B | ||
| MIC | 28.9 | C | ||
| Non-treated control | 29.4 | C | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Hernández, E.; González-García, V.; Casanova-Gascón, J.; Barriuso-Vargas, J.J.; Balduque-Gil, J.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Valorization of Quercus suber L. Bark as a Source of Phytochemicals with Antimicrobial Activity against Apple Tree Diseases. Plants 2022, 11, 3415. https://doi.org/10.3390/plants11243415
Sánchez-Hernández E, González-García V, Casanova-Gascón J, Barriuso-Vargas JJ, Balduque-Gil J, Lorenzo-Vidal B, Martín-Gil J, Martín-Ramos P. Valorization of Quercus suber L. Bark as a Source of Phytochemicals with Antimicrobial Activity against Apple Tree Diseases. Plants. 2022; 11(24):3415. https://doi.org/10.3390/plants11243415
Chicago/Turabian StyleSánchez-Hernández, Eva, Vicente González-García, José Casanova-Gascón, Juan J. Barriuso-Vargas, Joaquín Balduque-Gil, Belén Lorenzo-Vidal, Jesús Martín-Gil, and Pablo Martín-Ramos. 2022. "Valorization of Quercus suber L. Bark as a Source of Phytochemicals with Antimicrobial Activity against Apple Tree Diseases" Plants 11, no. 24: 3415. https://doi.org/10.3390/plants11243415
APA StyleSánchez-Hernández, E., González-García, V., Casanova-Gascón, J., Barriuso-Vargas, J. J., Balduque-Gil, J., Lorenzo-Vidal, B., Martín-Gil, J., & Martín-Ramos, P. (2022). Valorization of Quercus suber L. Bark as a Source of Phytochemicals with Antimicrobial Activity against Apple Tree Diseases. Plants, 11(24), 3415. https://doi.org/10.3390/plants11243415