

Influence of Different Intensities of Tillage on Physiological Characteristics and Productivity of Crop-Rotation Plants



Abstract
:1. Introduction
2. Results
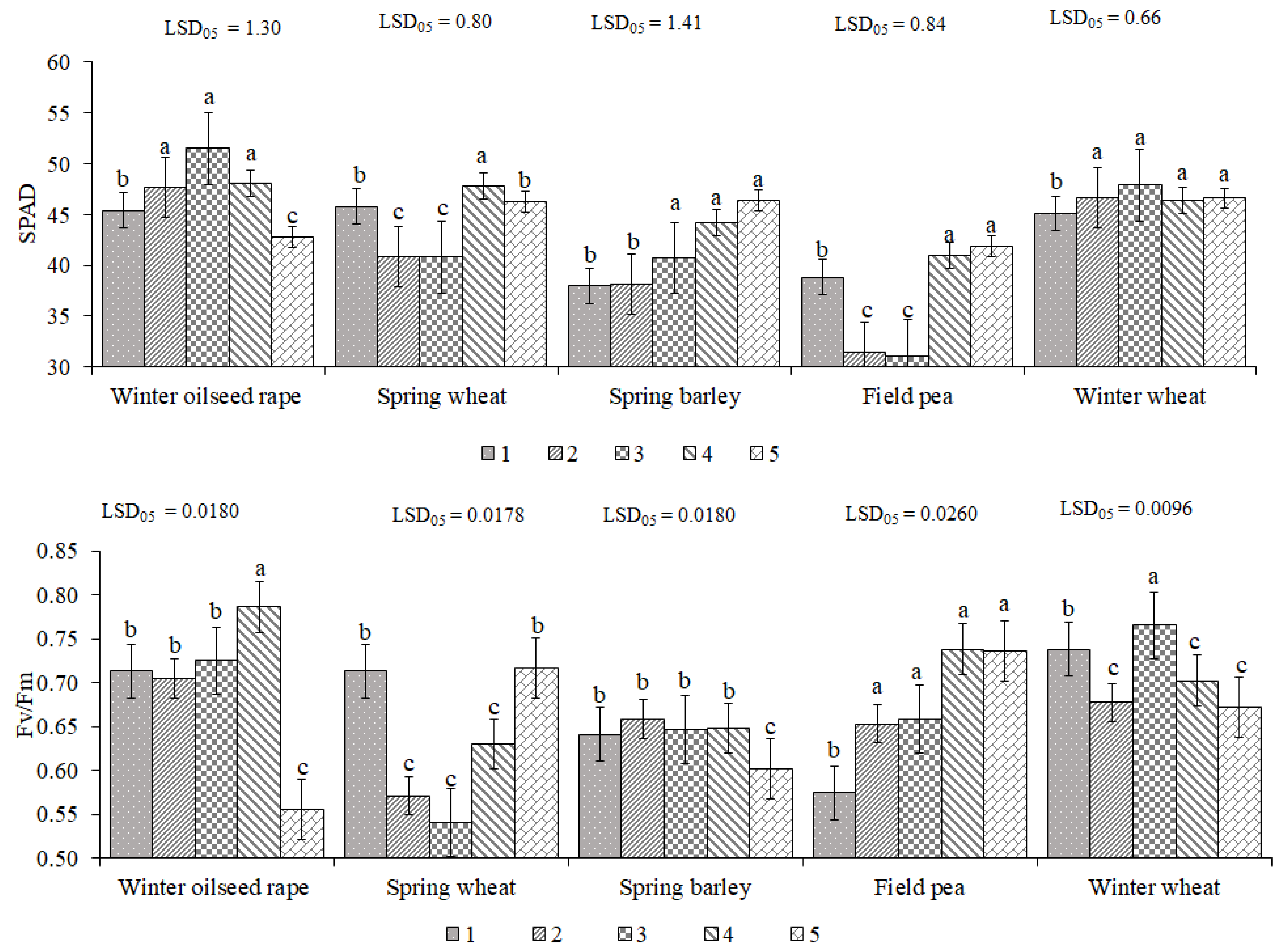
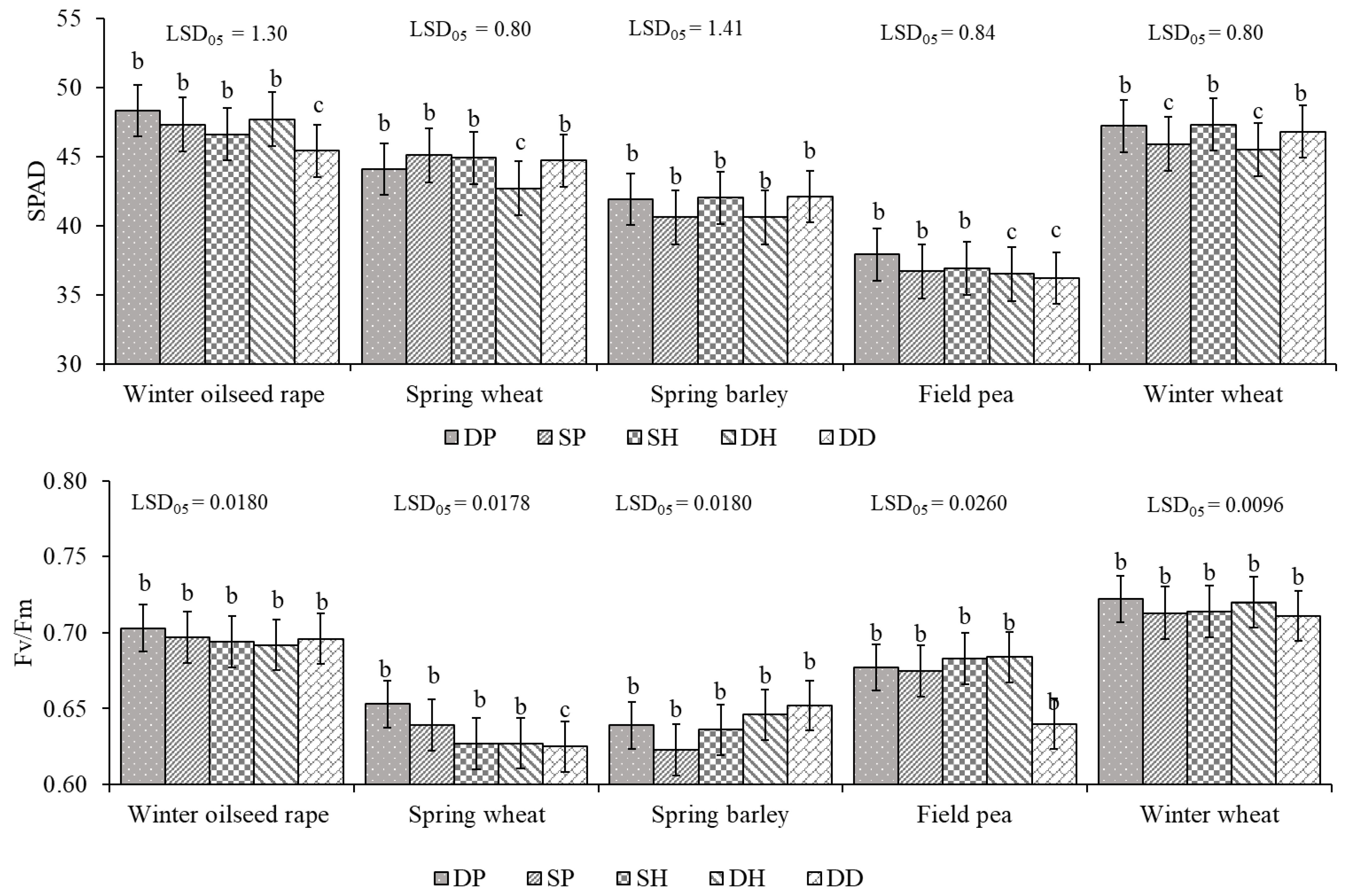
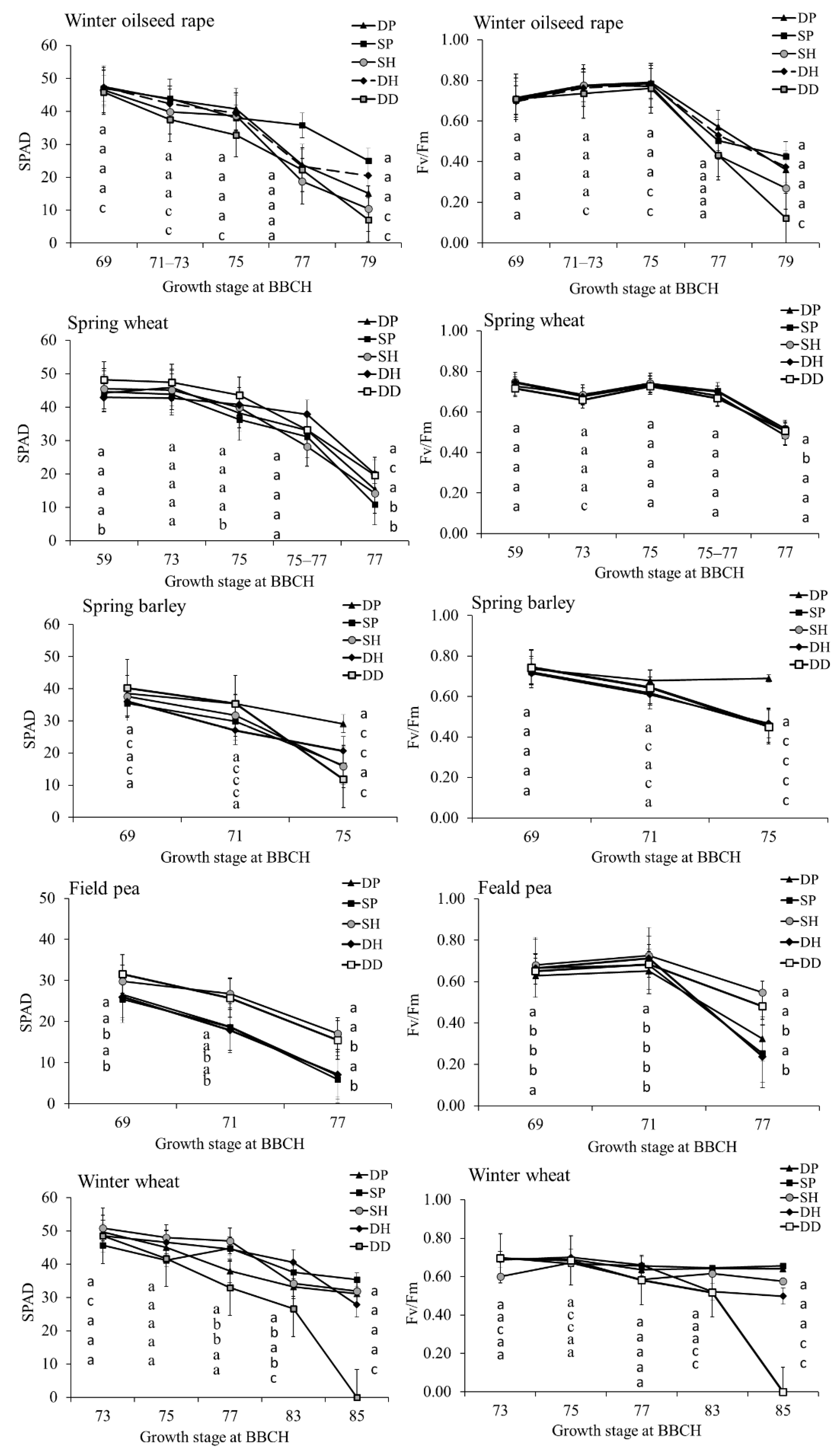
2.1. The Impact of Growth Stage and Tillage on Physiological Indices of Plants in Cropping System
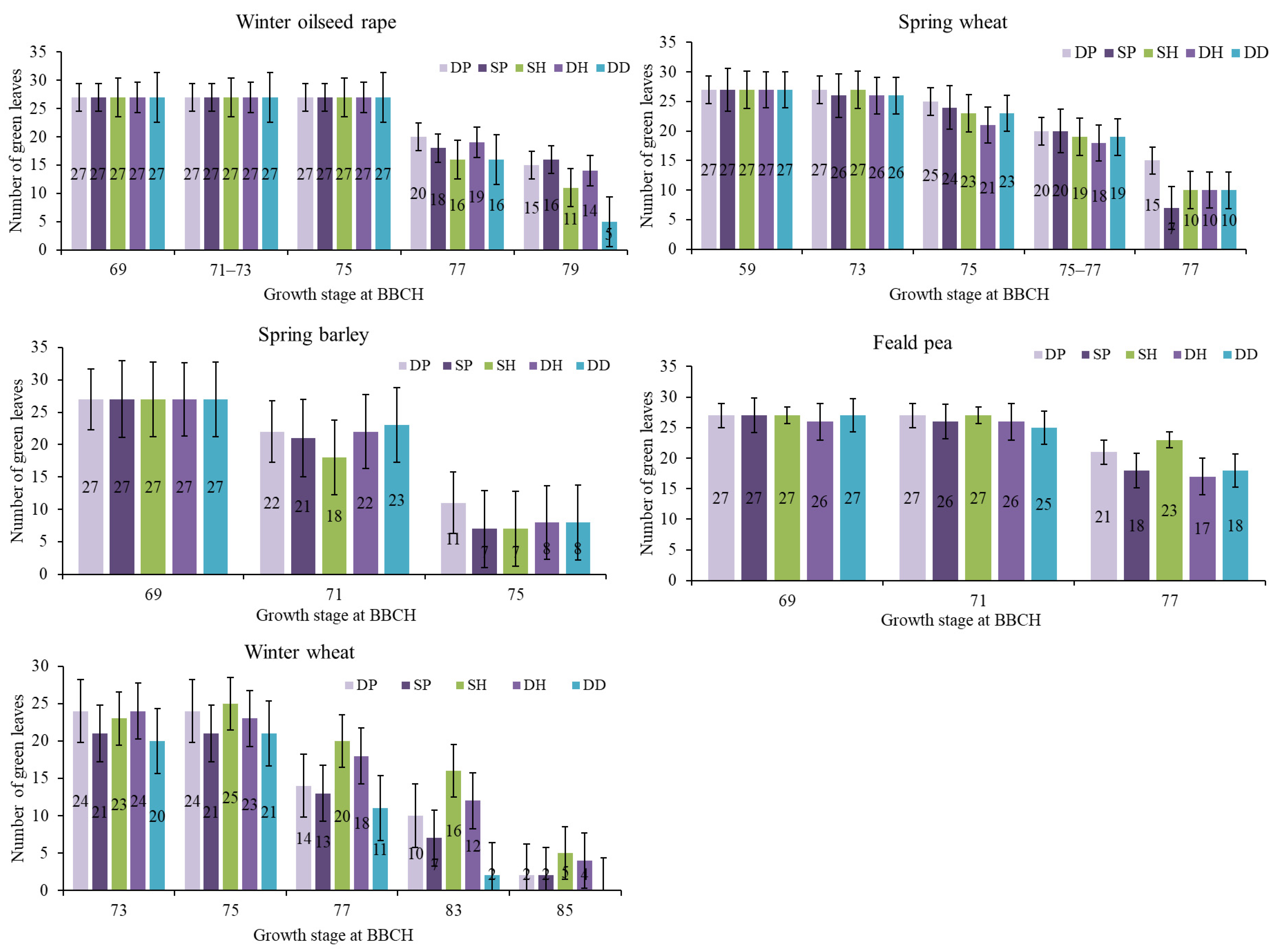
2.2. Tillage Intensity Effect on Leaf Senescence in the Final Growth Stages of Crops
3. Discussion
3.1. The Influence of Tillage on Crop Physiological Parameters
3.2. The Influence of Tillage on Crop Leaf Senescence
3.3. The Influence of Tillage on Productivity and Quality of Crop-Rotation Plants
3.4. Relationship between Grain Yield, Grain Quality, Soil Temperature, and Plant Physiological Traits under Different Tillage Intensities
4. Materials and Methods
4.1. Site and Soil Description
4.2. Experimental Details and Agronomic Management
4.3. Measurements of Physiological Parameters
4.4. Measurement of Crop Leaf Senescence
4.5. Measurement of Soil Temperature
4.6. Grain/Seed Yield (GY) and Grain/Seed Quality Analyses
4.7. Statistical Analysis
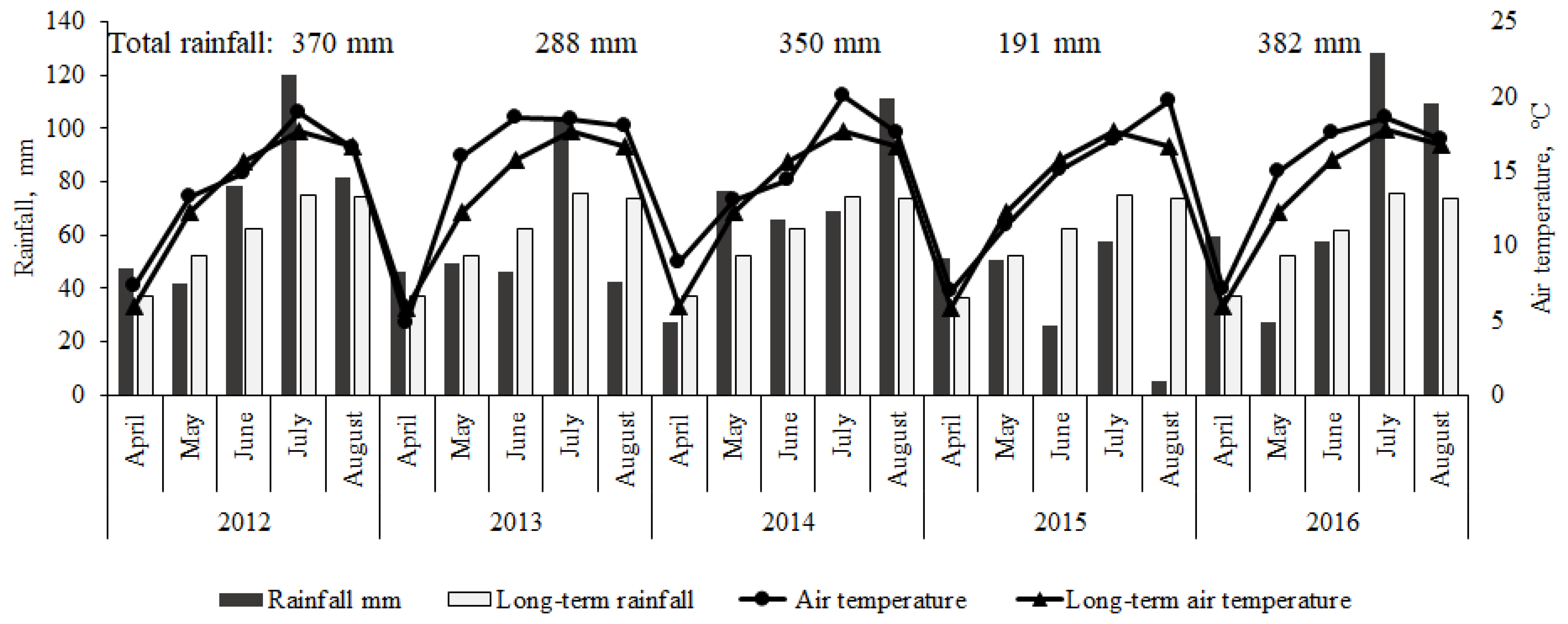
4.8. Meteorological Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brennan, J.; Hackett, R.; McCabe, T.; Grant, J.; Fortune, R.A.; Forristal, P.D. The effect of tillage system and residue management on grain yield and nitrogen use efficiency in winter wheat in a cool Atlantic climate. Eur. J. Agron. 2014, 54, 61–69. [Google Scholar] [CrossRef]
- Pelosi, C.; Grandeau, G.; Capowiez, Y. Temporal dynamics of earthworm-related macroporosity in tilled and non-tilled cropping systems. Geoderma 2017, 289, 169–177. [Google Scholar] [CrossRef]
- Feiziene, D.; Feiza, V.; Karklins, A.; Versuliene, A.; Janusauskaite, D.; Antanaitis, S. After-effects of long-term tillage and residue management on topsoil state in Boreal conditions. Eur. J. Agron. 2018, 94, 2–24. [Google Scholar] [CrossRef]
- Hofmeijer, M.A.J.; Krauss, M.; Berner, A.; Peigné, J.; Mäder, P.; Armengot, L. Effects of reduced tillage on weed pressure, nitrogen availability and winter wheat yields under organic management. Agronomy 2019, 9, 180. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.A.; Tedone, L.; Verdini, L.; Cazzato, E.; De Mastro, G. Wheat response to no-tillage and nitrogen fertilization in a long-term faba bean-based rotation. Agronomy 2019, 9, 50. [Google Scholar] [CrossRef] [Green Version]
- Macák, M.; Candráková, E.; Đalović, I.; Prasad, P.V.V.; Farooq, M.; Korczyk-Szabó, J.; Kováčik, P.; Šimanský, V. The influence of different fertilization strategies on the grain yield of field peas (Pisum sativum L.) under conventional and conservation tillage. Agronomy 2020, 10, 1728. [Google Scholar] [CrossRef]
- Li, Y.; Hou, R.; Tao, F. Wheat morphophysiological traits and radiation use efficiency under interactive effects of warming and tillage management. Plant Cell Environ. 2021, 44, 2386–2401. [Google Scholar] [CrossRef]
- Tolon-Becerra, A.; Tourn, M.; Botta, G.F.; Lastra-Bravo, X. Effects of different tillage regimes on soil compaction, maize (Zea mays L.) seedling emergence and yields in the eastern Argentinean Pampas region. Soil Tillage Res. 2011, 117, 184–190. [Google Scholar] [CrossRef]
- Singh, V.K.; Dwivedi, B.S.; Singh, S.K.; Majumdar, K.; Jat, M.L.; Mishra, R.P.; Rani, M. Soil physical properties, yield trends and economics after five years of conservation agriculture based rice-maize system in north-western India. Soil Tillage Res. 2016, 155, 133–148. [Google Scholar] [CrossRef]
- Schneider, F.; Don, A.; Hennings, I.; Schmittmann, O.; Seidel, S.J. The effect of deep tillage on crop yield–what do we really know? Soil Tillage Res. 2017, 174, 193–204. [Google Scholar] [CrossRef]
- Grzesiak, S.; Grzesiak, M.T.; Hura, T.; Marcinska, I.; Rzepka, A. Changes in root system structure, leaf water potential and gas exchange of maize and triticale seedlings affected by soil compaction. Environ. Exp. Bot. 2013, 88, 2–10. [Google Scholar] [CrossRef]
- Grzesiak, M.T. Impact of soil compaction on root architecture, leaf water status, gas exchange and growth of maize and triticale seedlings. Plant Root 2009, 3, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Grzesiak, M.T.; Janowiak, F.; Szczyrek, P.; Kaczanowska, K.; Ostrowska, A.; Rut, G.; Hura, T.; Rzepka, A.; Grzesiak, S. Impact of soil compaction stress combined with drought or waterlogging on physiological and biochemical markers in two maize hybrids. Acta Physiol. Plant. 2016, 38, 109. [Google Scholar] [CrossRef] [Green Version]
- Wasaya, A.; Tahir, M.; Ali, H.; Hussain, M.; Yasir, T.A.; Sher, A.; Ijaz, M.; Sattar, A. Influence of varying tillage systems and nitrogen application on crop allometry, chlorophyll contents, biomass production and net returns of maize (Zea mays L.). Soil Tillage Res. 2017, 170, 18–26. [Google Scholar] [CrossRef]
- Kan, Z.; Qi, J.; Liu, Q.; He, C.; Virk, A.L.; Lal, R.; Zhang, H. Effects of conservation tillage on wheat growth duration and grain yield in the North China Plain. Arch. Agron. Soil Sci. 2020, 68, 1019–1033. [Google Scholar] [CrossRef]
- Noulas, C.; Herrera, J.M.; Tziouvalekas, M.; Qin, R. Agronomic assessment of nitrogen use efficiency in spring wheat and interrelations with leaf greenness under field conditions. Commun. Soil Sci. Plant Anal. 2018, 49, 763–781. [Google Scholar] [CrossRef]
- Viljevac Vuletic, M.; Marcek, T.; Španic, V. Photosynthetic and antioxidative strategies of flag leaf maturation and its impact to grain yield of two field-grown wheat varieties. Theor. Exp. Plant Physiol. 2019, 31, 387–399. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, H.J.; Lim, P.O.; Nam, H.G. Leaf senescence: Systems and dynamics aspects. Ann. Rev. Plant Biol. 2019, 70, 347–376. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Early anthesis and delayed but fast leaf senescence contribute to individual grain dry matter and water accumulation in wheat. Field Crop. Res. 2016, 187, 24–34. [Google Scholar] [CrossRef]
- Gaju, O.; DeSilva, J.; Carvahlo, P.; Hawkesford, M.J.; Griffiths, S.; Greenland, A.; Foulkes, M.J. Leaf photosynthesis and associations with grain yield, biomass and nitrogen-use efficiency in landraces, synthetic-derived lines and cultivars in wheat. Field Crop. Res. 2016, 193, 1–15. [Google Scholar] [CrossRef]
- Li, Y.; Hou, R.; Tao, F. Interactive effects of different warming levels and tillage managements on winter wheat growth, physiological processes, grain yield and quality in the North China Plain. Agric. Ecosyst. Environ. 2020, 295, 106923. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledoneus Plants: BBCH Monograph, 2nd ed.; Blackwell Wissenschafts-Verlag: Braunschweig, Germany, 2001. [Google Scholar]
- Rohaček, K.; Soukupova, J.; Bartak, M. Chlorophyll fluorescence: A wonderful tool to study plant physiology and plant stress. In Plant Cell Compartments—Selected Topisc; Schoefs, B., Ed.; Res Signpost: Kerala, India, 2008; pp. 41–104. [Google Scholar]
- Dirsė, A.; Taparauskienė, L. Humidity fluctuations in plant vegetation periods and a comparison of its assessment methods. Žemės Ūkio Moksl. 2010, 17, 9–17. (In Lithuanian) [Google Scholar]
- del Pozo, A.; Yáñez, A.; Matus, I.A.; Tapia, G.; Castillo, D.; Sanchez-Jardón, L.; Araus, J.L. Physiological traits associated with wheat yield potential and performance under water-stress in a Mediterranean environment. Front. Plant Sci. 2016, 7, 987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibane, N.; Caicedo, M.; Martinez, S.; Marcet, P.; Revilla, P.; Ordas, B. Relationship between delayed leaf senescence (stay-green) and agronomic and physiological characters in maize (Zea mays L.). Agronomy 2021, 11, 276. [Google Scholar] [CrossRef]
- Hussain, S.; Ulhassan, Z.; Brestic, M.; Zivcak, M.; Zhou, W.I.; Allakhverdiev, S.I.; Yang, X.; Safdar, M.E.; Yang, W.; Liu, W. Photosynthesis research under climate change. Photosynth. Res. 2021, 150, 5–19. [Google Scholar] [CrossRef]
- Liu, J.; Fan, Y.; Ma, Y.; Li, Q. Response of photosynthetic active radiation interception, dry matter accumulation, and grain yield to tillage in two winter wheat genotypes. Arch. Agron. Soil Sci. 2019, 66, 1103–1114. [Google Scholar] [CrossRef]
- Kulig, B.; Lepiarczyk, A.; Oleksy, A.; Kolodziejczyk, M. The effect of tillage system and forecrop on the yield and values of LAI and SPAD indices of spring wheat. Eur. J. Agron. 2010, 33, 43–51. [Google Scholar] [CrossRef]
- Chen, S.; Xia, G.; Zhao, W.; Wu, F.; Zhang, G. Characterization of leaf photosynthetic properties for no-tillage rice. Rice Sci. 2007, 14, 283–288. [Google Scholar] [CrossRef]
- Liu, K.; Wiatrak, P. Corn production response to tillage and nitrogen application in dry-land environment. Soil Tillage Res. 2012, 124, 138–143. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Physiol. Biochem. 2014, 81, 6–25. [Google Scholar] [CrossRef]
- Feng, W.; He, L.; Zhang, H.; Guo, B.; Zhu, Y.; Wang, C.; Guo, T. Assessment of plant nitrogen status using chlorophyll fluorescence parameters of the upper leaves in winter wheat. Eur. J. Agron. 2015, 64, 78–87. [Google Scholar] [CrossRef]
- Sonko, R.; Marchenko, O.; Kolomijez, V.; Starodub, N. Express Biosensor control of maize plants under different agrotechnical procedures. In Proceedings of the 14th International Meeting on Chemical Sensors, Nürnberg, Germany, 20–23 May 2012; pp. 1328–1331. [Google Scholar] [CrossRef]
- Luoni, S.B.; Astigueta, F.H.; Nicosia, S.; Moschen, S.; Fernandez, P.; Heinz, R. Transcription factors associated with leaf senescence in crops. Plants 2019, 8, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyneke, E.; Watanabe, M.; Erban, A.; Duan, G.; Buchner, P.; Walther, D.; Kopka, J.; Hawkesford, M.J.; Hoefgen, R. Effect of senescence phenotypes and nitrate availability on wheat leaf metabolome during grain filling. Agronomy 2019, 9, 305. [Google Scholar] [CrossRef] [Green Version]
- Borrill, P.; Fahy, B.; Smith, A.M.; Uauy, C. Wheat grain filling is limited by grain filling capacity rather than the duration of flag leaf photosynthesis: A case study using NAM RNAi plants. PLoS ONE 2015, 10, e0134947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Tang, Y.; Li, C.; McHugh, A.D.; Li, Z.; Wu, C. Individual and combined effects of soil waterlogging and compaction on physiological characteristics of wheat in southwestern China. Field Crop. Res. 2018, 215, 163–172. [Google Scholar] [CrossRef]
- He, J.; Shi, Y.; Zhao, J.; Yu, Z. Strip rotary tillage with subsoiling increases winter wheat yield by alleviating leaf senescence and increasing grain filling. Crop J. 2020, 8, 327–340. [Google Scholar] [CrossRef]
- Copec, K.; Filipovic, D.; Husnjak, S.; Kovacev, I.; Kosutic, S. Effects of tillage systems on soil water content and yield in maize and winter wheat production. Plant Soil Environ. 2015, 61, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Kovačev, I.; Bilandžija, N.; Čopec, K.; Filipovič, D. Impact of soil tillage reduction on cultivation productivity of wheat, barley and soybean. Sci. Pap. Ser. A Agron. 2018, 61, 291–295. [Google Scholar]
- Muñoz-Romero, V.; López-Bellido, L.; López-Bellido, R.J. Effect of tillage system on soil temperature in a rainfed Mediterranean Vertisol. Int. Agrophys. 2015, 29, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77. [Google Scholar] [CrossRef]
- Drebenstedt, I.; Schmidt, I.; Poll, C.; Marhan, S.; Kahle, R.; Kandeler, E.; Högy, P. Effects of soil warming and altered precipitation patterns on photosynthesis, biomass production and yield of barley. J. Appl. Bot. Food Qual. 2020, 93, 44–53. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Indices | Growth Stage (A) | Tillage Intensity (B) | A × B | Total |
|---|---|---|---|---|---|
| Winter oilseed rape | SPAD | 45.1 ** | 5.3 * | 5.2 | 55.6 |
| Fv/Fm | 74.9 ** | 0.2 | 5.5 | 80.6 | |
| Spring wheat | SPAD | 63.0 ** | 5.5 ** | 3.6 | 72.1 |
| Fv/Fm | 70.9 ** | 1.6 | 7.1 | 79.6 | |
| Spring barley | SPAD | 43.1 ** | 1.8 | 13.0 | 57.8 |
| Fv/Fm | 16.0 ** | 3.9 | 13.3 | 33.2 | |
| Field pea | SPAD | 83.4 ** | 1.3 | 2.5 | 87.3 |
| Fv/Fm | 45.7 ** | 3.2 | 12.1 | 31.0 | |
| Winter wheat | SPAD | 14.7 ** | 10.4 * | 12.1 | 37.1 |
| Fv/Fm | 63.6 ** | 2.2 | 8.4 | 74.2 |
| Crop | Tillage | Grain Yield (t ha−1) | Protein (%) | TGW (g) | HLW (kg hl−1) |
|---|---|---|---|---|---|
| Winter oilseed rape | DP | 3.72 ± 0.10 b | § 45.1 ± 0.24 b | 6.37 ± 0.10 b | - |
| SP | 3.54 ± 0.11 b | § 45.2 ± 0.29 b | 6.35 ± 0.08 b | - | |
| SH | 3.26 ± 0.15 c | § 45.7 ± 0.17 a | 6.37 ± 0.07 b | - | |
| DH | 3.57 ± 0.01 b | § 45.9 ± 0.22 a | 6.38 ± 0.05 b | - | |
| DD | 3.33 ± 0.10 c | § 45.6 ± 0.19 b | 6.35 ± 0.02 b | - | |
| LSD05 | 0.294 | 0.60 | 0.230 | ||
| Spring wheat | DP | 5.86 ± 0.11 b | 13.1 ± 0.26 b | 40.0 ± 0.31 b | 75.4 ± 0.56 b |
| SP | 6.02 ± 0.17 b | 13.1 ± 0.23 b | 39.9 ± 0.60 b | 75.8 ± 0.50 b | |
| SH | 5.98 ± 0.18 b | 13.2 ± 0.18 b | 40.6 ± 0.59 b | 75.3 ± 1.03 b | |
| DH | 5.97 ± 0.11 b | 12.5 ± 0.17 c | 39.5 ± 0.34 b | 74.5 ± 0.84 b | |
| DD | 5.11 ± 0.22 c | 13.2 ± 0.19 b | 39.6 ± 0.30 b | 73.8 ± 0.77 b | |
| LSD05 | 0.530 | 0.44 | 1.30 | 2.56 | |
| Spring barley | DP | 6.91 ± 0.33 b | 11.8 ± 0.18 b | 43.9 ± 0.41 b | 61.0 ± 0.51 b |
| SP | 6.72 ± 0.26 b | 12.5 ± 0.27 a | 44.0 ± 0.29 b | 60.8 ± 0.50 b | |
| SH | 7.01 ± 0.17 b | 11.8 ± 0.13 b | 42.9 ± 0.24 c | 61.0 ± 0.17 b | |
| DH | 6.51 ± 0.07 b | 11.3 ± 0.2 b | 41.8 ± 0.11 c | 60.3 ± 0.42 b | |
| DD | 5.72 ± 0.49 c | 10.9 ± 0.14 c | 43.6 ± 0.16 b | 61.0 ± 0.45 b | |
| LSD05 | 0.981 | 0.63 | 0.72 | 1.44 | |
| Field pea | DP | 2.29 ± 0.18 b | - * | 223.7 ± 4.46 b | 87.8 ± 0.56 b |
| SP | 2.57 ±0.18 b | - | 222.9 ± 3.09 b | 86.8 ± 0.52 b | |
| SH | 2.07 ± 0.09 b | - | 222.7 ± 3.55 b | 87.2 ± 0.41 b | |
| DH | 2.40 ± 0.17 b | - | 227.5 ± 2.32 b | 88.2 ± 0.29 b | |
| DD | 2.00 ± 0.26 b | - | 229.8 ± 5.81 b | 87.0 ± 1.01 b | |
| LSD05 | 0.499 | 12.04 | 1.85 | ||
| Winter wheat | DP | 8.26 ± 0.15 b | 12.8 ± 0.05 b | 37.4 ± 0.54 b | 81.2 ± 0.43 b |
| SP | 7.92 ± 0.13 b | 12.7 ± 0.13 b | 38.4 ± 0.92 b | 80.5 ± 1.19 b | |
| SH | 7.89 ± 0.04 b | 12.5 ± 0.09 c | 39.2 ± 0.86 b | 80.3 ± 0.98 b | |
| DH | 7.90 ± 0.27 b | 12.4 ± 0.11 c | 38.7 ± 1.13 b | 80.2 ± 0.71 b | |
| DD | 8.21 ± 0.16 b | 12.3 ± 0.05 c | 38.6 ± 0.63 b | 81.7 ± 0.54 b | |
| LSD05 | 0.438 | 0.20 | 2.32 | 2.21 |
| Tillage Intensity | Indices | Fv/Fm | GY | Protein † | TGW | HLM § | T0–5 cm |
|---|---|---|---|---|---|---|---|
| DP | SPAD | 0.331 ** | 0.576 ** | 0.413 ** | −0.550 ** | −0.113 | −0.297 ** |
| Fv/Fm | 0.058 | 0.234 * | 0.057 | 0.287 ** | −0.242 * | ||
| GY | −0.844 ** | −0.912 ** | −0.502 ** | −0.538 ** | |||
| Protein † | −0.99 ** | 0.739 ** | −0.391 ** | ||||
| TGW | 0.645 ** | 0.470 ** | |||||
| HLM | 0.290 ** | ||||||
| SP | SPAD | 0.198 | 0.584 ** | 0.314 ** | −0.572 ** | −0.092 | −0.213 |
| Fv/Fm | 0.081 | 0.204 | 0.073 | 0.368 ** | −0.210 | ||
| GY | −0.892 ** | −0.929 ** | −0.509 ** | −0.540 ** | |||
| Protein † | −0.989 ** | 0.285 * | −0.413 ** | ||||
| TGW | 0.626 ** | 0.484 ** | |||||
| HLM | 0.331 ** | ||||||
| SH | SPAD | 0.252 * | 0.624 ** | 0.204 | −0.624 ** | −0.170 | −0.338 ** |
| Fv/Fm | −0.060 | 0.204 | 0.161 | 0.290 ** | −0.114 | ||
| GY | −0.921 ** | −0.948 ** | −0.596 ** | −0.504 ** | |||
| Protein † | −0.995 ** | 0.672 ** | −0.391 ** | ||||
| TGW | 0.662 ** | 0.426 ** | |||||
| HLM | 0.316 ** | ||||||
| DH | SPAD | 0.205 | 0.554 ** | 0.440 ** | −0.513 ** | −0.155 | −0.262 * |
| Fv/Fm | 0.029 | 0.164 | 0.139 | 0.297 ** | −0.143 | ||
| GY | −0.878 ** | −0.929 ** | 0.526 ** | −0.546 ** | |||
| Protein † | −0.996 ** | 0.854 ** | −0.406 ** | ||||
| TGW | 0.690 ** | 0.500 ** | |||||
| HLM | 0.320 ** | ||||||
| DD | SPAD | 0.192 | 0.602 ** | 0.097 | −0.623 ** | −0.195 | −0.270 * |
| Fv/Fm | 0.184 | 0.191 | −0.110 | 0.042 | −0.187 | ||
| GY | −0.720 ** | −0.825 ** | −0.277 ** | −0.530 ** | |||
| Protein † | −0.995 ** | 0.664 ** | −0.423 ** | ||||
| TGW | 0.639 ** | 0.413 ** | |||||
| HLM | 0.148 |
| Crops | Sowing Time | Cultivar | Seed Rate (Viable Seeds mln ha−1) | Crop Density (Plants m2) | Fertilizer NPK (kg ha−1) |
|---|---|---|---|---|---|
| Winter oilseed rape | 24 August 2012 | Komando | 0.7 | 45 | N194 P53 K105 |
| Spring wheat | 2 May 2013 | Granary | 5.0 | 256 | N152 P48 K102 |
| Spring barley | 17 April 2014 | Grace | 4.0 | 294 | N132 P48 K102 |
| Field pea | 14 April 2015 | Pinocchio | 1.0 | 69 | N14 P56 K119 |
| Winter wheat | 10 September 2016 | Ada | 4.5 | 395 | N184 P56 K119 |
| Measurement No. | |||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Crop | Growth Stage (BBCH) | ||||
| Winter oilseed rape | 60–61 | 63–65 | 69 | 71–73 | 75–78 |
| Spring wheat | 32 | 39–41 | 51–53 | 59 | 71 |
| Spring barley | 31 | 39 | 49 | 59 | 69 |
| Field pea | 15 | 17–18 | 39 | 61 | 69 |
| Winter wheat | 32 | 41–43 | 55 | 61 | 63 |
| Crops | Growth Stage (BBCH) | ||||
|---|---|---|---|---|---|
| Winter oilseed rape | 69 | 71–73 | 75 | 77 | 79 |
| Spring wheat | 59 | 73 | 75 | 75–77 | 77 |
| Spring barley | 69 | 71 | 75 | - | - |
| Field pea | 69 | 71 | 77 | - | - |
| Winter wheat | 73 | 75 | 77 | 83 | 85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janusauskaite, D.; Kadziene, G. Influence of Different Intensities of Tillage on Physiological Characteristics and Productivity of Crop-Rotation Plants. Plants 2022, 11, 3107. https://doi.org/10.3390/plants11223107
Janusauskaite D, Kadziene G. Influence of Different Intensities of Tillage on Physiological Characteristics and Productivity of Crop-Rotation Plants. Plants. 2022; 11(22):3107. https://doi.org/10.3390/plants11223107
Chicago/Turabian StyleJanusauskaite, Daiva, and Grazina Kadziene. 2022. "Influence of Different Intensities of Tillage on Physiological Characteristics and Productivity of Crop-Rotation Plants" Plants 11, no. 22: 3107. https://doi.org/10.3390/plants11223107
APA StyleJanusauskaite, D., & Kadziene, G. (2022). Influence of Different Intensities of Tillage on Physiological Characteristics and Productivity of Crop-Rotation Plants. Plants, 11(22), 3107. https://doi.org/10.3390/plants11223107