
Salt Tolerance Potential in Onion: Confirmation through Physiological and Biochemical Traits

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization and Evaluation of Onion Accessions
2.2. Morphometric and Yield-Related Traits
2.3. Physio-Biochemical and Ion Estimation
2.3.1. Physiological Traits
2.3.2. Biochemical Traits
2.3.3. Ion Concentration
2.4. Statistical Analysis
3. Results
3.1. Effects of Salinity on Selected Onion Accessions
3.2. Impact of Salt Stress on Growth and Bulb Parameters
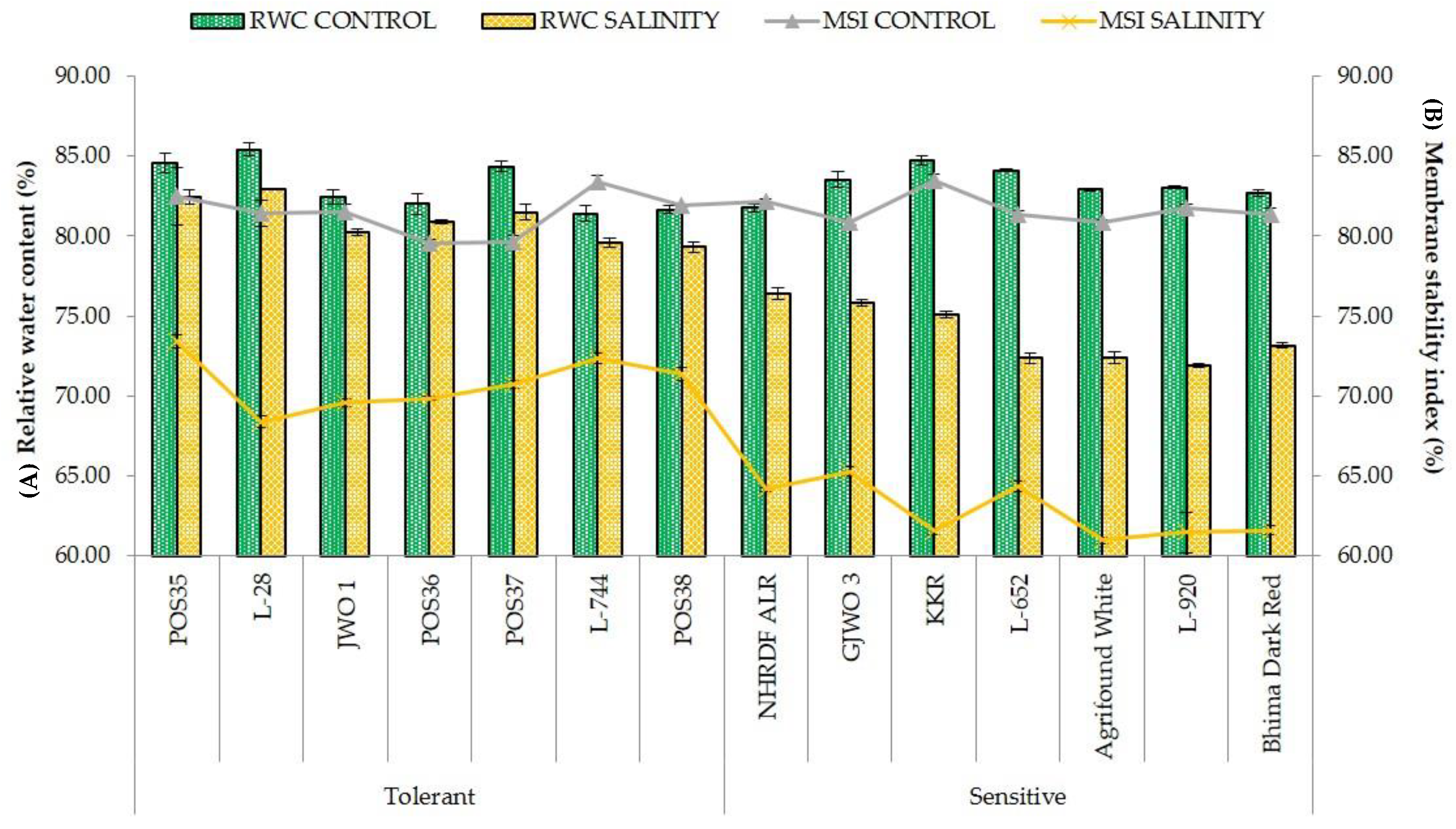
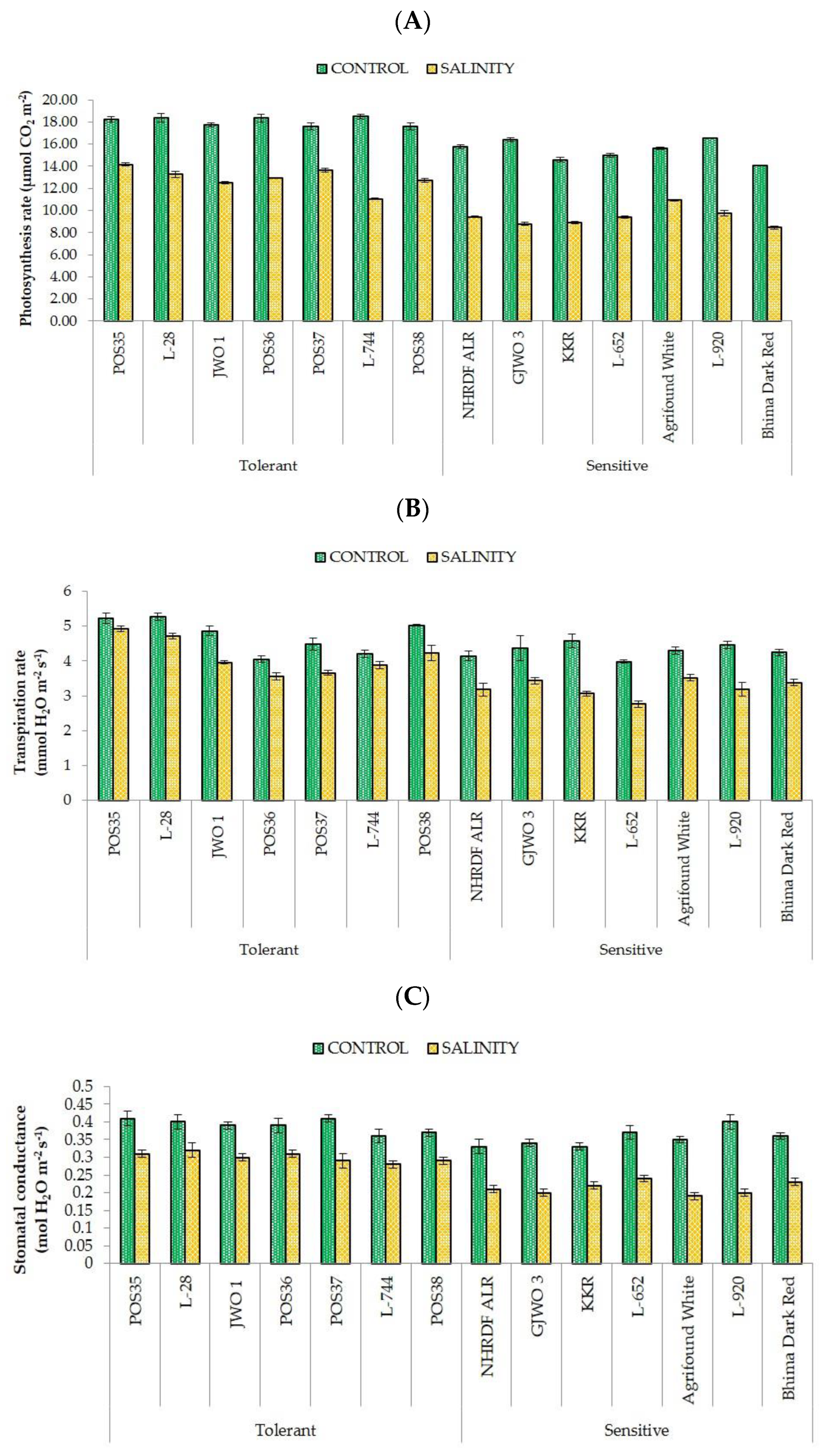
3.3. Physiological Responses
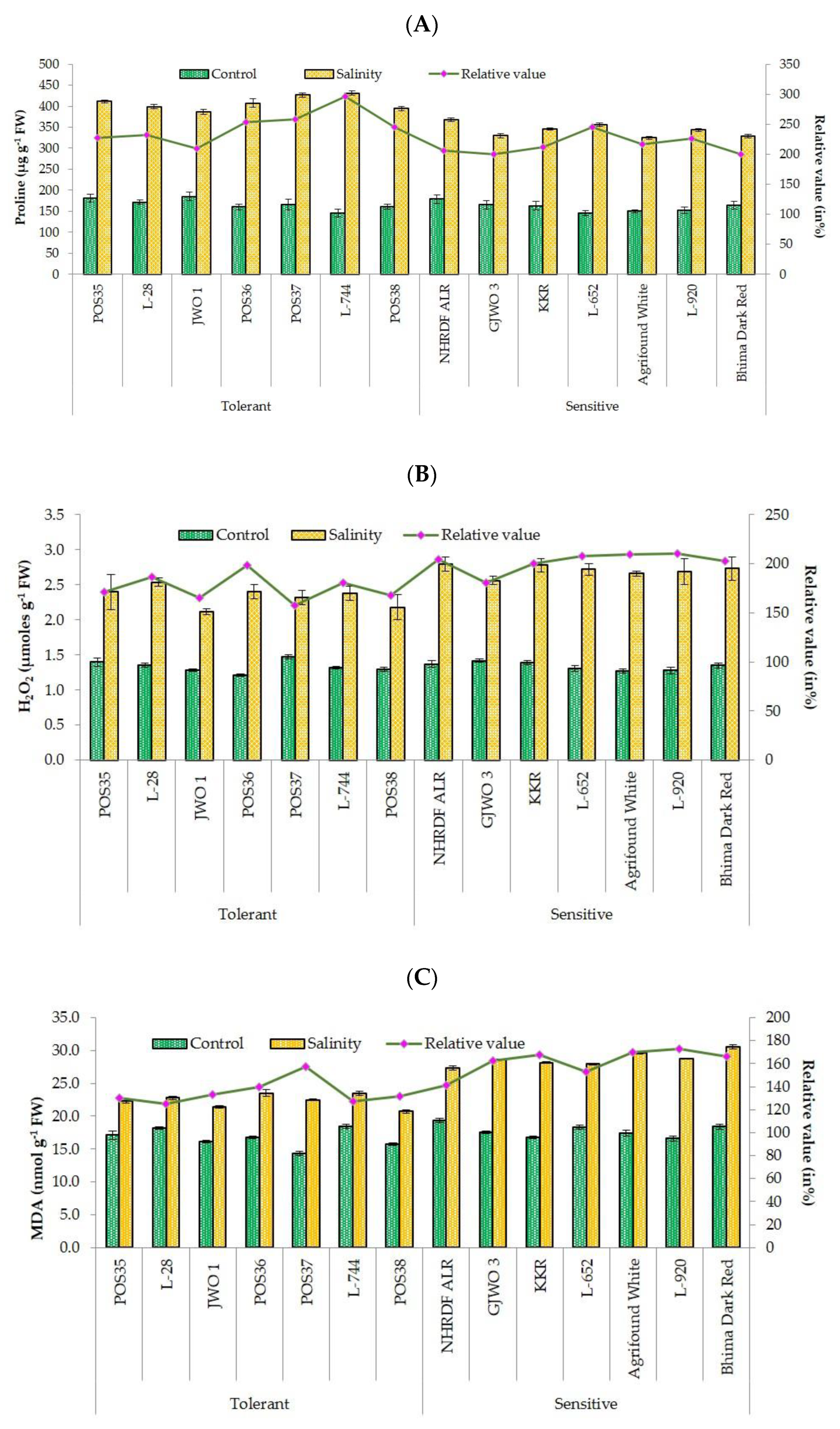
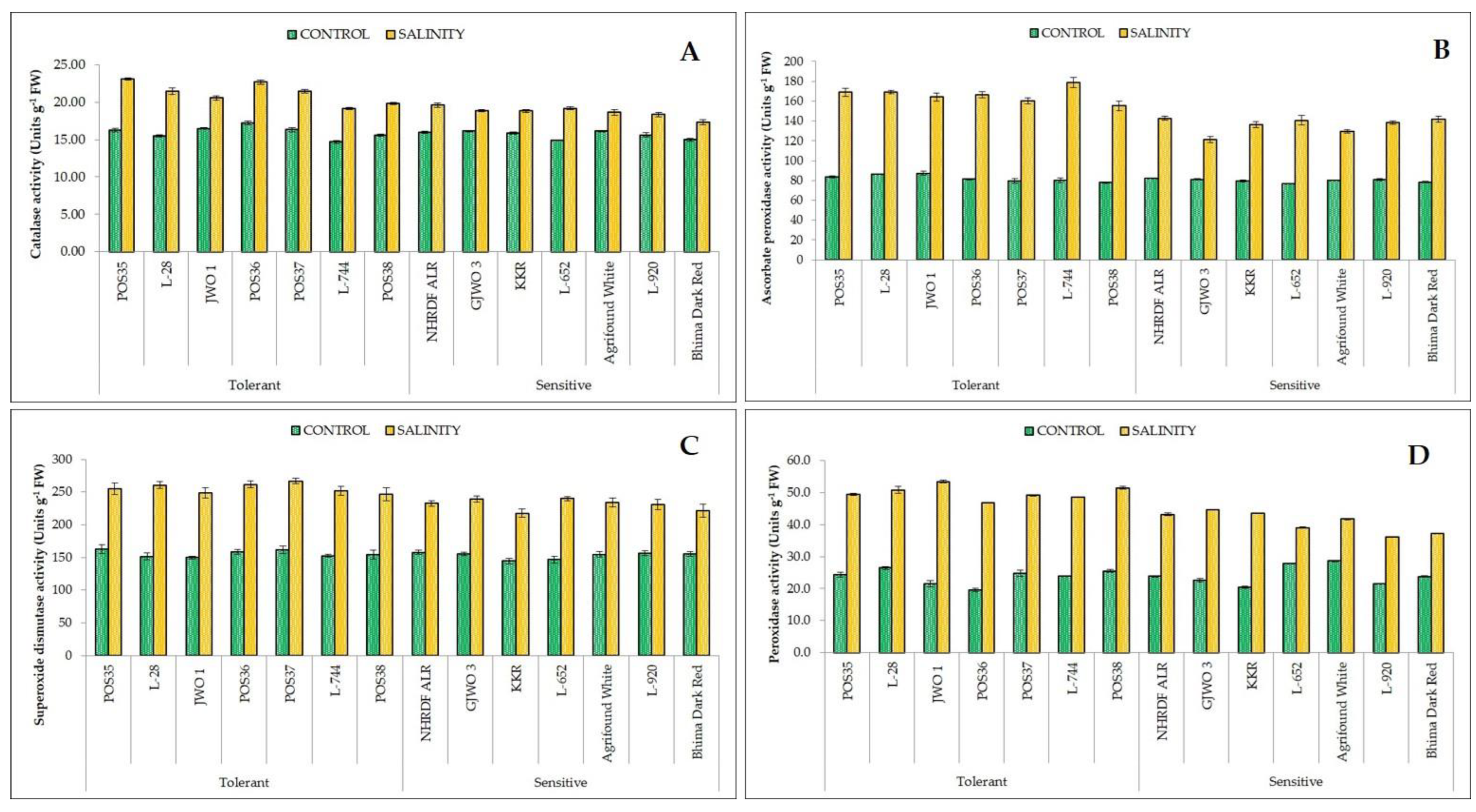
3.4. Biochemical Responses
3.5. Accumulation of Ion Concentrations
3.6. Traits Contributing towards Bulb Yield Divergence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2020. [Google Scholar]
- Khar, A.; Lawande, K.E.; Negi, K.S. Microsatellite marker based analysis of genetic diversity in short day tropical Indian onion and cross amplification in related Allium spp. Genet. Res. Crop Evol. 2011, 58, 741–752. [Google Scholar] [CrossRef]
- Ricciardi, L.; Mazzeo, R.; Marcotrigiano, A.R.; Rainaldi, G.; Iovieno, P.; Zonno, V.; Pavan, S.; Lotti, C. Assessment of genetic diversity of the “Acquaviva Red Onion” (Allium cepa L.) apulian landrace. Plants 2020, 9, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, L.; Alarcon, E.; Trevithick-Sutton, C.; Gandhi, N.; Scaiano, J.C. Effect of c- radiation on green onion DNA integrity: Role of ascorbic acid and polyphenols against nucleic acid damage. Food Chem. 2011, 128, 735–741. [Google Scholar] [CrossRef]
- El Balla, M.D.; Hamid, A.A.; Abdelmageed, A.H. Effects of time of water stress on flowering, seed yield and seed quality of common onion (Allium cepa L.) under the arid tropical conditions of Sudan. Agric. Water Manag. 2013, 121, 149–157. [Google Scholar] [CrossRef]
- Garcia, G.; Garcia, M.; Ramirez, H. Performance in seven Allium cepa L. cultivars at different salt stress levels. Bioagro 2015, 27, 93–102. [Google Scholar]
- Rahneshan, Z.; Nasibi, F.; Moghadam, A.A. Effects of salinity stress on some growth, physiological, biochemical parameters and nutrients in two pistachio (Pistacia vera L.) rootstocks. J. Plant Interact. 2018, 13, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kumar, J.; Singh, V.P.; Prasad, S.M. Plant tolerance mechanism against salt stress: The nutrient management approach. Biochem. Pharmacol. 2014, 3, e165. [Google Scholar] [CrossRef] [Green Version]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity Stress in Potato: Understanding physiological, biochemical and molecular responses. Life 2021, 11, 545. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. Sci. Hortic. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Venancio, J.B.; da Silva Dias, N.; de Medeiros, J.F.; de Moraes, P.L.D.; do Nascimento, C.W.A.; de Sousa Neto, O.N.; da Silva Sa, F.V. Yield and morphophysiology of onion grown under salinity and fertilization with silicon. Sci. Hortic. 2022, 301, 111095. [Google Scholar] [CrossRef]
- Silva, E.N.; Silveira, J.A.G.; Rodrigues, C.R.F.; Viegas, R.A. Physiological adjustment to salt stress in Jatropha curcas is associated with accumulation of salt ions, transport and selectivity of K+, osmotic adjustment and K+/Na+ homeostasis. Plant Biol. 2015, 17, 1023–1029. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–87. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Kumar, A.; Mishra, A.K.; Singh, K.; Lata, C.; Kumar, A.; Krishnamurthy, S.L.; Kumar, P. Diurnal changes and effect of elevated CO2 on gas exchange under individual and interactive salt and water stress in wheat (Triticum aestivum). Indian J. Agric. Sci. 2019, 89, 763. [Google Scholar]
- Bates, L.S.; Walden, R.P.; Teare, I.D. Rapid determination of proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Chawla, S.; Jain, S.; Jain, V. Salinity induced oxidative stress and antioxidant system in salt-tolerant and salt-sensitive cultivars of rice (Oryza sativa L.). J. Plant Biochem. Biotechnol. 2013, 22, 27–34. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Rao, M.V.; Watkins, C.B.; Brown, S.K.; Weeden, N.F. Active oxygen species metabolism in White Angel x Rome Beauty apple selections resistant and susceptible to superficial scald. J. Am. Soc. Hortic. Sci. 1998, 123, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- IRRI. Biometrics and Breeding Informatics, STAR version 2.0.1; International Rice Research Institute: Los Baños, PH, USA, 2013. [Google Scholar]
- Singh, D. The relative importance of characters affecting genetic divergence. Indian J. Genet. Plant Breed. 1981, 41, 237–245. [Google Scholar]
- Ghodke, P.H.; Andhale, P.S.; Gijare, U.M.; Thangasamy, A.; Khade, Y.P.; Mahajan, V.; Singh, M. Physiological and biochemical responses in onion crop to drought stress. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 2054–2062. [Google Scholar] [CrossRef]
- Sanwal, S.K.; Mann, A.; Kesh, H.; Kaur, G.; Kumar, R.; Rai, A.K. Genotype environment interaction analysis for fruit yield in okra (Abelmoschus esculentus L.) under alkaline environments. Indian J. Genet. 2021, 81, 101–110. [Google Scholar] [CrossRef]
- Diaz-Lopez, L.; Gimeno, V.; Lidon, V.; Simon, I.; Martinez, V.; García-Sanchez, F. The tolerance of Jatropha curcas seedlings to NaCl: An ecophysiological analysis. Plant Physiol. Biochem. 2012, 54, 34–42. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Liu, Z.; Ke, W.; Hou, H. Effect of Salt Stress on growth, physiological parameters, and ionic concentration of Water Dropwort (Oenanthe javanica) Cultivars. Front. Plant Sci. 2021, 12, 660409. [Google Scholar] [CrossRef]
- Rauf, S.; Adil, M.S.; Naveed, A.; Munir, H. Response of wheat species to the contrasting saline regimes. Pak. J. Bot. 2010, 42, 3039–3045. [Google Scholar]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of bread wheat genotypes for drought tolerance using phenotypic and proline analyses. Front. Plant Sci. 2016, 7, 1276. [Google Scholar] [CrossRef] [Green Version]
- Gedam, P.A.; Thangasamy, A.; Shirsat, D.V.; Ghosh, S.; Bhagat, K.P.; Sogam, O.A.; Gupta, A.J.; Mahajan, V.; Soumia, P.S.; Salunkhe, V.N.; et al. Screening of Onion (Allium cepa L.) Genotypes for drought tolerance using physiological and yield based indices through multivariate analysis. Front. Plant Sci. 2021, 12, 600371. [Google Scholar] [CrossRef] [PubMed]
- Geravandi, M.; Farshadfar, E.; Kahrizi, D. Evaluation of some physiological traits as indicators of drought tolerance in bread wheat genotypes. Russ. J. Plant Physl. 2011, 58, 69–75. [Google Scholar] [CrossRef]
- Sanwal, S.K.; Kaur, G.; Mann, A.; Kumar, R.; Rai, A.K. Response of okra (Abelmoschus esculentus L.) genotypes to salinity stress in relation to seedling stage. J. Soil Salin. Water Qual. 2019, 11, 217–223. [Google Scholar]
- Mahlooji, M.; Seyed Sharifi, R.; Razmjoo, J.; Sabzalian, M.R.; Sedghi, M. Effect of salt stress on photosynthesis and physiological parameters of three contrasting barley genotypes. Photosynthetica. 2018, 56, 549–556. [Google Scholar] [CrossRef]
- Meriem, B.F.; Kaouther, Z.; Cherif, H.; Tijani, M.; Andre, B. Effect of priming on growth, biochemical parameters and mineral composition of different cultivars of coriander (Coriandrum sativum L.) under salt stress. J. Stress Physiol. Biochem. 2014, 10, 84–109. [Google Scholar]
- Sharif, P.; Seyedsalehi, M.; Paladino, O.; Damme, P.; Van Sillanpa, M.; Sharif, A.A. Effect of drought and salinity stresses on morphological and physiological characteristics of canola. Int. J. Environ. Sci. Technol. 2017, 15, 1859–1866. [Google Scholar] [CrossRef]
- Elhakem, A.H. Growth, water relations, and photosynthetic activity are associated with evaluating salinity stress tolerance of wheat cultivars. Int. J. Agron. 2020, 2020, 8882486. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sanwal, S.K.; Kumar, A.; Mann, A.; Kaur, G. Differential response of pea (Pisum sativum) genotypes exposed to salinity in relation to physiological and biochemical attributes. Indian J. Agric. Sci. 2018, 88, 149–156. [Google Scholar]
- Gomes, M.A.C.; Suzuki, M.S.; Cunha, M.D.; Tulli, C.F. Effect of salt stress on nutrient concentration, photosynthetic pigments, proline and foliar morphology of Salvinia auriculata Aubl. Acta Limnol. Bras. 2011, 23, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Jamil, M.; Rehman, S.; Rha, E.S. Salinity effect on plant growth, PSII photochemistry and chlorophyll content in sugar beet (Beta vulgaris L.) and cabbage (Brassica oleracea capitata L.). Pak. J. Bot. 2007, 39, 753–760. [Google Scholar]
- Kanwal, H.; Ashraf, M.; Shahbaz, M. Assessment of salt tolerance of some newly developed and candidate wheat (Triticum aestivum L.) cultivars using gas exchange and chlorophyll fluorescence attributes. Pak. J. Bot. 2011, 43, 2693–2699. [Google Scholar]
- Perveen, S.; Shahbaz, M.; Ashraf, M. Influence of foliar-applied triacontanol on growth, gas exchange characteristics, and chlorophyll fluorescence at different growth stages in wheat under saline conditions. Photosynthetica. 2013, 51, 541–551. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Ranjbar, G.; Pakniyat, H.; Emam, Y. Physiological mechanisms of salt stress tolerance in plants: An overview. In Plant Environment Interaction: Responses and Approaches to Mitigate Stress; Chapter 8; Mahgoub Azooz, M., Ahmad, P., Eds.; John Wiley & Sons, Ltd.: Noida, India, 2016; pp. 141–160. [Google Scholar]
- Ali-Dinar, H.M.; Ebert, G.; Ludders, P. Growth, Chlorophyll Content, Photosynthesis and Water Relations in Guava (Psidium guajava L.) under Salinity and Different Nitrogen Supply. Gartenbauwissenschaft 1999, 64, 54–59. [Google Scholar]
- Sarker, U.; Oba, S. The response of salinity stress-induced A. tricolor to growth, anatomy, physiology, non-enzymatic and enzymatic antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Hasan, M.; Arora, A.; Gaikwad, K.; Kumar, S.; Rai, R.D.; Singh, A. Sodium chloride-induced spatial and temporal manifestation in membrane stability index and protein profiles of contrasting wheat (Triticum aestivum L.) genotypes under salt stress. Ind. J. Plant Physiol. 2015, 20, 271–275. [Google Scholar] [CrossRef]
- Rao, S.P.; Mishra, B.; Gupta, S.R.; Rathore, A. Physiological response to salinity and alkalinity of rice genotypes of varying salt tolerance grown in field Lysimeters. J. Stress Physiol. Biochem. 2013, 9, 54–65. [Google Scholar]
- Yassin, M.; El Sabagh, A.; Mekawy, A.M.; Islam, M.; Hossain, A.; Barutcular, C.; Alharby, H.; Bamagoos, A.; Liu, L.; Ueda, A.; et al. Comparative performance of two bread wheat (Triticum aestivum L.) genotypes under salinity stress. Appl. Ecol. Environ. Res. 2019, 17, 5029–5041. [Google Scholar] [CrossRef]
- Alzahrani, S.M.; Alaraidh, I.A.; Migdadi, H.; Alghamdi, S.; Altaf Khan, M.; Ahmad, P. Physiological, biochemical, and antioxidant properties of two genotypes of Vicia faba grown under salinity stress. Pak. J. Bot. 2019, 51, 786–798. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Beena, A.S.; Awana, M.; Singh, A. Physiological, biochemical, epigenetic and molecular analyses of wheat (Triticum aestivum) genotypes with contrasting salt tolerance. Front. Plant Sci. 2017, 8, 1151. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.; Daud, M.K.; Zulqurnain, M.; Ali, S. Seed priming by sodium nitroprusside improves salt tolerance in wheat (Triticum aestivum L.) by enhancing physiological and biochemical parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Polash, M.A.S.; Sakil, M.A.; Hossain, M.A. Plants responses and their physiological and biochemical defense mechanisms against salinity: A review. Trop. Plant Res. 2019, 6, 250–274. [Google Scholar] [CrossRef]
- Li, Y. Physiological responses of tomato seedlings (Lycopersicon esculentum) to salt stress. Mod. Appl. Sci. 2009, 3, 171–176. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of individual and combined effects of salinity and drought on physiological, nutritional and biochemical properties of cabbage (Brassica oleracea var. capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Sanwal, S.K.; Kumar, P.; Kesh, H.; Gupta, V.K.; Kumar, A.; Kumar, A.; Meena, B.L.; Colla, G.; Cardarelli, M.; Kumar, P. Salinity Stress Tolerance in Potato Cultivars: Evidence from Physiological and Biochemical Traits. Plants 2022, 11, 1842. [Google Scholar] [CrossRef]
- Sanwal, S.K.; Mann, A.; Kumar, A.; Kesh, H.; Kaur, G.; Rai, A.K.; Kumar, R.; Sharma, P.C.; Kumar, A.; Bahadur, A.; et al. Salt Tolerant Eggplant Rootstocks Modulate Sodium Partitioning in Tomato Scion and Improve Performance under Saline Conditions. Agriculture. 2022, 12, 183. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Behera, T.K.; Krishna, R.; Ansari, W.A.; Aamir, M.; Kumar, P.; Kashyap, S.P.; Pandey, S.; Kole, C. Approaches Involved in the Vegetable Crops Salt Stress Tolerance Improvement: Present Status and Way Ahead. Front. Plant Sci. 2022, 12, 787292. [Google Scholar] [CrossRef]
- Tsujii, M.; Kera, K.; Hamamoto, S.; Kuromori, T.; Shikanai, T.; Uozumi, N. Evidence for potassium transport activity of Arabidopsis KEA1-KEA6. Sci. Rep. 2019, 9, 10040. [Google Scholar] [CrossRef] [Green Version]
- Menezes, R.V.; Azevedo Neto, A.D.D.; Ribeiro, M.D.O.; Cova, A.M.W. Growth and contents of organic and inorganic solutes in amaranth under salt stress. Pesqui. Agropecuária Trop. 2017, 47, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Bojarian, M.; Asadi-Gharneh, H.A.; Golabadi, M. Factor analysis, stepwise regression and path coefficient analyses of yield, yield-associated traits, and fruit quality in tomato. Int. J. Veg. Sci. 2019, 25, 542–553. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Pessarakli, M.; Heidari, B. Comparing relationships among yield and its related traits in mycorrhizal and nonmycorrhizal inoculated wheat cultivars under different water regimes using multivariate statistics. Int. J. Agron. 2013, 2013, 682781. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Varieties | Parentage | Year of Release | Maturity (Days) | Bulb Color | Bulb Shape | Bulb Size | Storability (Months) |
|---|---|---|---|---|---|---|---|---|
| 1. | Pusa Red | Mass Selection (Local collection) | 1975 | 125–140 | Bronze red | Flat to globular | Medium | 3–4 |
| 2. | Kalyanpur Red Round (KKR) | Mass Selection (Collection from UP) | 1983 | 110–120 | Red | Round | Medium | 3–4 |
| 3. | ALR | Mass Selection (Collection from Nasik, Maharashtra) | 1988 | 110–120 | Light red | Globular | Medium | 3–4 |
| 4. | B-780 (Baswant780) | Mass Selection (Collection from Pimpalgaon, Maharashtra) | 1989 | 110–120 | Dark red | Round | Large | 2–3 |
| 5. | Agrifound White | Mass Selection (Local Collection) | 1994 | 160–165 | White | Round | Medium | 1–2 |
| 6. | Punjab Naroya | Mass selection of material collected from Maharashtra | 1997 | 115–120 | Red | Round | Medium to large | 2–3 |
| 7. | GJWO-1 | Mass Selection (Collection from Gujarat) | 2000 | 130–135 | White | Round | Medium | 2–3 |
| 8. | RO 59 (Arpita) | Mass Selection (Local Collection) | 2005 | 110–120 | Red | Globe | Medium | 2–3 |
| 9. | Arka Pitambar | Selection from U.D. 102 × IAR-396 | 2006 | 135–140 | Yellow | Globe | Medium | 3–4 |
| 10. | NHRDF Red (L-28) | Collected from Patiala, Punjab and developed at RRS, Karnal. | 2006 | 180–210 | Dark red | Round | Medium | 2–3 |
| 11. | Bhima Super | Rigorous mass selection for single centeredness and bulb shape | 2006 | 100–110 | Red | Flat globe | Large | 2–3 |
| 12. | Bhima Raj | Single bulb selection up to three generations | 2007 | 120–125 | Red | Oval | Medium | 2–3 |
| 13. | Bhima Red | Single bulb selection up to three generations | 2009 | 115–120 | Red | Round | Medium | 2–3 |
| 14. | Bhima Shweta | (White El. Comp. Selection/NRCWO2) (IC No. 572761) | 2010 | 110–120 | White | Round | Medium | 2–3 |
| 15. | Bhima Shakti | IC No. 572769 | 2010 | 125–135 | Red | Round | Large | 5–6 |
| 16. | POS38 | Selfing (two generations) and massing from Bhima Kiran | 2010 | 125–135 | Light red | Oval to round | Large | 5–6 |
| 17. | Arka Bheem | Triparental synthetic | 2011 | 125–130 | Red to pinkish red | Elongated globe | Medium | 1–2 |
| 18. | RO-252 | Mass Selection (Local Collection) | 2011 | - | Red | - | Medium | 2–3 |
| 19. | Bhima Dark Red | Single bulb selection up to three generations | 2012 | 100–110 | Dark red | Flat round | Medium | 1–1.5 |
| 20. | NHRDF Red-3 (L-652) | Mass Selection (Local Collection) | 2012 | 100–120 | Bronze red | Globular round | Medium | 2–3 |
| 21. | Bhima Safed | Mass Selection (Local Collection) | 2014 | 110–120 | White | Round to oval | Medium | 1–1.5 |
| 22. | JRO 11 (GJRO 11) | Local collection from Mahuva Taluka of Bhavnagar district | 2015 | 125–130 | Red | Flat globe | Medium | 3–4 |
| 23. | Bhima Light Red | Single bulb selection followed by mass selection | 2015 | 115–120 | Light red | Globe | Large | 2–3 |
| 24. | HOS 4 (Hisar onion 4) | Selection from local material collected from Bahadurgarh area | 2016 | 130–140 | Light Bronze | Globular | Medium | 3–4 |
| 25. | GJWO 3 | Local collection from Talaja Taluka of Bhavnagar district Germplasm No. 120 | 2016 | 125–130 | White | Flat globe | Medium | 2–3 |
| 26. | NHRDF Red-4 (L-744) | Mass Selection (Local Collection) | 2016 | 110–120 | Red | Globular round | Medium | 2–3 |
| 27. | Pusa Shobha | Mass selection from segregating material from local collection | 2018 | 140–160 | Brown | Flat globe | Large | 1–2 |
| 28. | Pusa Sona | Mass selection from segregating material of Early Grano | 2019 | 125–135 | Creamy yellow | Globular | Large | 1–2 |
| 29. | POS35 | Selection-Selfing (two generations) followed by massing from Pusa White Round selection | - | 125–130 | White | Round | Medium | 2–3 |
| 30. | POS36 | Selection- Selfing (two generations) followed by massing from Pusa Madavi | - | - | Brown | Globe | Large | 2–3 |
| 31. | POS39 | Single plant selection from Hisar 2 with waxy leaves (Hisar2 Waxy) | - | 125–130 | Bronze red | Flat globular | Medium | 3–4 |
| 32. | POS37 | Selfing (two generations) followed by massing from Pusa White Flat | - | 120–130 | White | Flat round | Medium large | 2–3 |
| 33. | Sukhsagar | Local landrace from West Bengal | - | 90–100 | Dark red | Globe | Medium | 3–4 |
| 34. | PRO 6 | Mass Selection (Collection from Punjab) | 115–120 | Deep red | Round | Medium large | 2–3 | |
| 35. | JNDWO-85 | Mass Selection (Collection from Gujarat) | - | - | White | - | - | 3–4 |
| 36. | NHRDF (L-920) | Mass Selection (Local Collection) | - | - | - | - | Medium | 2–3 |
| Parameters | Initial Soil Status | Final Soil Status | |
|---|---|---|---|
| Control | After Saline Treatment | ||
| 2019–20 | |||
| ECe (dS m−1) | 1.65 | 1.85 | 6.90 |
| pHs | 7.74 | 7.67 | 7.82 |
| 2020–21 | |||
| ECe (dS m−1) | 1.48 | 1.62 | 7.04 |
| pHs | 7.61 | 7.65 | 7.93 |
| Variables | Mean Squares | F Values | Significance | |||
|---|---|---|---|---|---|---|
| Genotypes | G X E | Genotypes | G X E | Genotypes | G X E | |
| Df | 13 | 13 | 13 | 13 | Pr(>F) | Pr(>F) |
| #SPAD Index | 21.66 | 3.26 | 54.97 | 8.26 | 0.000 | 0.000 |
| RWC | 28.51 | 24.69 | 233.79 | 202.46 | 0.000 | 0.000 |
| MSI | 31.88 | 32.85 | 112.93 | 116.38 | 0.000 | 0.000 |
| Pn | 17.51 | 1.88 | 451.38 | 48.55 | 0.000 | 0.000 |
| E | 1.49 | 0.19 | 83.42 | 10.41 | 0.000 | 0.000 |
| gS | 0.01 | 0.00 | 36.77 | 9.25 | 0.000 | 0.000 |
| Proline | 2596.14 | 2045.07 | 47.65 | 37.54 | 0.000 | 0.000 |
| H2O2 | 0.09 | 0.08 | 9.44 | 8.25 | 0.000 | 0.000 |
| MDA | 26.56 | 13.90 | 315.53 | 165.17 | 0.000 | 0.000 |
| CAT | 7.22 | 2.89 | 154.75 | 61.89 | 0.000 | 0.000 |
| APX | 524.63 | 403.66 | 89.91 | 69.18 | 0.000 | 0.000 |
| SOD | 481.60 | 279.97 | 15.71 | 9.13 | 0.000 | 0.000 |
| POX | 53.36 | 58.10 | 287.12 | 312.58 | 0.000 | 0.000 |
| Root Na+/K+ | 0.51 | 0.20 | 51.19 | 20.36 | 0.000 | 0.000 |
| Shoot Na+/K+ | 1.50 | 0.92 | 578.09 | 354.26 | 0.000 | 0.000 |
| Bulb Na+/K+ | 0.00 | 0.00 | 9.74 | 8.97 | 0.000 | 0.000 |
| PH | 31.05 | 3.86 | 82.25 | 10.23 | 0.000 | 0.000 |
| NL | 0.82 | 0.02 | 44.17 | 1.01 | 0.000 | 0.453 |
| BD | 1.64 | 0.18 | 86.95 | 9.28 | 0.000 | 0.000 |
| BYP | 1361.39 | 134.57 | 11990.75 | 1185.22 | 0.000 | 0.000 |
| TSS | 6.40 | 3.28 | 284.75 | 146.11 | 0.000 | 0.000 |
| Salt-Tolerant Genotypes | Bulb Yield Reduction (<20%) | Moderately Tolerant Genotypes | Bulb Yield Reduction (20–40%) | Salt-Sensitive Genotypes | Bulb Yield Reduction (>40%) |
|---|---|---|---|---|---|
| POS35 | 14.88 | Bhima Red | 20.20 | ALR | 42.48 |
| L-28 | 15.13 | POS39 | 20.55 | GJWO 3 | 42.63 |
| GJWO 1 | 16.35 | Bhima Shweta | 24.72 | KKR | 43.50 |
| POS36 | 17.19 | JNDWO-85 | 26.65 | L-652 | 43.65 |
| L-744 | 18.80 | HOS 4 (Hisar onion 4) | 29.43 | Agrifound White | 43.89 |
| POS37 | 19.48 | Arka Pitamber | 29.52 | L-920 | 48.60 |
| POS38 | 19.76 | Pusa Red | 29.56 | Bhima Dark Red | 62.86 |
| RO 59 (Arpita) | 29.99 | ||||
| Pusa Shobha | 31.47 | ||||
| Bhima Light Red | 32.15 | ||||
| Punjab Naroya | 32.31 | ||||
| GJRO 11 | 32.92 | ||||
| Bhima Raj | 34.60 | ||||
| PRO 6 | 34.60 | ||||
| B-780 (Baswant780) | 34.69 | ||||
| Bhima Shakti | 34.96 | ||||
| Bhima Super | 35.25 | ||||
| Bhima Safed | 35.27 | ||||
| RO-252 | 37.11 | ||||
| Sukhsagar | 38.22 | ||||
| Pusa Sona | 38.44 | ||||
| Arka Bheem | 39.62 |
| Cultivars Name | Plant Height (cm) | Number of Leaves | Bulb Diameter (cm) | Bulb Yield (g plant−1) | TSS (°Brix) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Salinity | Control | Salinity | Control | Salinity | Control | Salinity | Control | Salinity | |
| POS35 | 43.22 ± 0.31 de | 42.20 ± 0.43 de | 8.30 ± 0.04 | 8.27 ± 0.25 | 6.18 ± 0.04 a | 5.62 ± 0.13 a | 77.48 ± 0.59 a | 65.95 ± 0.05 a | 12.43 ± 0.03 e | 12.00 ± 0.06 d |
| L-28 | 43.95 ± 0.64 cd | 43.48 ± 0.73 abc | 9.01 ± 0.11 | 8.98 ± 0.08 | 5.80 ± 0.08 b | 5.44 ± 0.10 ab | 77.00 ± 0.20 a | 65.35 ± 0.35 b | 12.07 ± 0.07 g | 10.67 ± 0.13 g |
| GJWO 1 | 43.82 ± 0.43 d | 42.62 ± 0.81 bcd | 8.17 ± 0.24 | 7.96 ± 0.06 | 5.31 ± 0.04 cd | 4.94 ± 0.06 c | 58.70 ± 0.30 g | 49.10 ± 0.10 f | 12.37 ± 0.10 ef | 10.37 ± 0.07 h |
| POS36 | 43.25 ± 0.78 de | 42.26 ± 0.55 de | 8.35 ± 0.10 | 8.15 ± 0.13 | 5.87 ± 0.13 b | 5.02 ± 0.08 c | 66.95 ± 0.20 f | 55.44 ± 0.44 e | 13.37 ± 0.10 e | 12.30 ± 0.30 c |
| POS37 | 43.20 ± 0.40 de | 42.55 ± 0.44 cde | 8.70 ± 0.15 | 8.48 ± 0.29 | 5.69 ± 0.52 b | 5.54 ± 0.07 a | 70.65 ± 0.15 d | 56.89 ± 0.11 d | 12.77 ± 0.10 d | 12.07 ± 0.07 cd |
| L-744 | 45.43 ± 0.58 ab | 44.03 ± 0.62 a | 8.85 ± 0.05 | 8.71 ± 0.10 | 5.92 ± 0.10 b | 5.29 ± 0.10 b | 74.38 ± 1.20 b | 60.40 ± 0.40 c | 12.17 ± 0.17 fg | 11.63 ± 0.20 e |
| POS38 | 38.75 ± 0.32 h | 37.65 ± 0.78 g | 9.06 ± 0.11 | 8.91 ± 0.09 | 5.28 ± 0.14 cd | 4.87 ± 0.07 c | 69.67 ± 0.10 e | 55.90 ± 0.10 e | 12.10 ± 0.10 g | 11.50 ± 0.20 e |
| ALR | 44.88 ± 1.18 bc | 41.45 ± 0.69 e | 8.76 ± 0.10 | 8.56 ± 0.20 | 4.70 ± 0.10 e | 4.17 ± 0.13 ef | 41.53 ± 0.10 k | 23.89 ± 0.11 j | 12.90 ± 0.10 d | 12.20 ± 0.20 cd |
| GJWO 3 | 41.93 ± 0.60 f | 37.46 ± 0.55 g | 7.99 ± 0.06 | 7.84 ± 0.16 | 5.38 ± 0.06 c | 4.21 ± 0.09 def | 49.30 ± 0.30 j | 28.28 ± 0.28 h | 12.50 ± 0.10 e | 14.85 ± 0.06 a |
| KKR | 42.25 ± 0.73 ef | 37.65 ± 0.78 g | 7.90 ± 0.08 | 7.87 ± 0.20 | 5.06 ± 0.04 d | 4.43 ± 0.13 d | 40.00 ± 0.25 l | 22.60 ± 0.60 k | 15.77 ± 0.10 b | 11.00 ± 0.15 f |
| L-652 | 42.37 ± 0.65 ef | 39.82 ± 0.83 f | 8.45 ± 0.15 | 8.30 ± 0.09 | 5.26 ± 0.11 cd | 4.32 ± 0.12 de | 55.10 ± 0.10 h | 31.05 ± 0.05 g | 12.93 ± 0.07 d | 12.07 ± 0.07 cd |
| Agrifound White | 40.66 ± 0.62 g | 35.49 ± 1.06 h | 8.25 ± 0.05 | 8.10 ± 0.09 | 5.10 ± 0.06 d | 4.86 ± 0.06 c | 50.22 ± 0.22 i | 28.18 ± 0.18 h | 13.60 ± 0.30 c | 11.97 ± 0.03 d |
| L-920 | 46.42 ± 0.56 a | 43.67 ± 0.90 ab | 8.08 ± 0.08 | 8.06 ± 0.06 | 4.72 ± 0.06 e | 3.50 ± 0.06 g | 41.56 ± 0.20 k | 21.36 ± 0.36 l | 16.10 ± 0.05 a | 14.30 ± 0.30 b |
| Bhima Dark Red | 45.26 ± 0.75 b | 42.08 ± 0.35 de | 8.74 ± 0.14 | 8.29 ± 0.07 | 5.09 ± 0.09 d | 4.06 ± 0.08 f | 72.10 ± 0.10 c | 26.78 ± 0.10 i | 12.17 ± 0.17 fg | 11.10 ± 0.10 f |
| Trait | Unit | Groups Mean | Mean Square | Significance | |
|---|---|---|---|---|---|
| Tolerant Genotypes | Sensitive Genotypes | Tolerant vs. Sensitive | Pr(>F) | ||
| SPAD Index | Value | 44.90 ± 1.40 a | 42.29 ± 2.05 b | 33.571 | 0.000 |
| Relative Water Content (RWC) | % | 80.98 ± 1.36 a | 73.87 ± 1.77 b | 116.433 | 0.000 |
| Membrane Stability Index (MSI) | % | 70.81 ± 1.66 a | 62.76 ± 1.72 b | 215.15 | 0.000 |
| Photosynthesis rate (Pn) | µmol CO2 m−2 s−1 | 12.91 ± 0.95 a | 9.38 ± 0.79 b | 8.397 | 0.000 |
| Transpiration rate (E) | mmol H2O m−2 s−1 | 4.13 ± 0.50 a | 3.22 ± 0.27 a | 0.024 | 0.160 |
| Stomatal conductance (gS) | mol H2O m−2 s−1 | 0.30 ± 0.02 a | 0.21 ± 0.02 b | 0.006 | 0.000 |
| Proline content (Pro) | µg g−1 FW | 407.88 ± 17.29 a | 342.62 ± 16.77 b | 11853.07 | 0.000 |
| Hydrogen peroxide (H2O2) | µmoles g−1 FW | 2.33 ± 0.18 b | 2.71 ± 0.12 a | 0.666 | 0.000 |
| Malondialdehyde (MDA) | nmol g−1 FW | 22.38 ± 0.99 b | 28.70 ± 1.06 a | 108.58 | 0.000 |
| Catalase (CAT) | units g−1 FW | 21.18 ± 1.39 a | 18.68 ± 0.72 b | 5.838 | 0.000 |
| Ascorbate peroxidase (APX) | units g−1 FW | 166.11 ± 7.70 a | 135.77 ± 7.75 b | 870.59 | 0.000 |
| Super oxide dismutase (SOD) | units g−1 FW | 255.76 ± 9.20 a | 230.75 ± 9.70 b | 1687.96 | 0.000 |
| Peroxidase (POX) | units g−1 FW | 49.96 ± 2.07 a | 40.78 ± 3.18 b | 333.26 | 0.000 |
| Root Na+/K+ | Index | 3.22 ± 0.13 b | 3.50 ± 0.25 a | 0.367 | 0.000 |
| Shoot Na+/K+ | Index | 2.11 ± 0.43 b | 3.57 ± 0.37 a | 1.765 | 0.000 |
| Bulb Na+/K+ | Index | 0.32 ± 0.05 b | 0.34 ± 0.04 a | 0.004 | 0.007 |
| Plant height (PH) | Cm | 41.82 ± 1.97 a | 39.95 ± 3.12 b | 8.52 | 0.000 |
| Number of leaves/plant (NL) | Nos. | 8.43 ± 0.35 a | 8.21 ± 0.38 a | 0.065 | 0.101 |
| Bulb diameter (BD) | Cm | 5.07 ± 0.47 a | 4.40 ± 0.59 b | 0.138 | 0.001 |
| Total soluble solids (TSS) | °Brix | 11.72 ± 0.65 b | 12.28 ± 1.58 a | 2.024 | 0.000 |
| Bulb yield/plant (BYP) | g plant−1 | 52.51 ± 12.92 a | 31.94 ± 14.34 b | 52.371 | 0.000 |
| Traits | Traits Contribution (%) under Control Conditions | Traits Contribution (%) under Salinity Stress Conditions | Traits Mean ± SD | Magnitude # (%) | |
|---|---|---|---|---|---|
| Control | Salinity Stress | ||||
| SPAD Index | 0.414 | 0.293 | 45.49 ± 0.49 | 43.60 ± 0.34 | −4.15 |
| Relative Water Content (RWC) | 0.707 | 2.580 | 83.18 ± 0.33 | 77.43 ± 0.26 | −6.92 |
| Membrane Stability Index (MSI) | 0.227 | 1.617 | 81.54 ± 0.43 | 66.79 ± 0.35 | −18.09 |
| Photosynthesis rate (Pn) | 2.637 | 3.018 | 16.74 ± 0.21 | 11.15 ± 0.14 | −33.42 |
| Transpiration rate (E) | 0.478 | 0.484 | 4.51 ± 0.13 | 3.67 ± 0.11 | −18.53 |
| Stomatal conductance (gS) | 0.191 | 0.256 | 0.37 ± 0.02 | 0.26 ± 0.01 | −31.09 |
| Proline content (Pro) | 0.374 | 0.256 | 163.63 ± 4.75 | 375.25 ± 8.42 | +129.33 |
| Hydrogen peroxide (H2O2) | 0.263 | 0.043 | 1.34 ± 0.03 | 2.52 ± 0.11 | +88.38 |
| Malondialdehyde (MDA) | 0.960 | 3.266 | 17.23 ± 0.30 | 25.54 ± 0.22 | +48.27 |
| Catalase (CAT) | 1.423 | 0.607 | 15.83 ± 0.18 | 19.93 ± 0.25 | +25.90 |
| Ascorbate peroxidase (APX) | 0.498 | 0.430 | 81.07 ± 0.98 | 150.94 ± 3.24 | +86.18 |
| Super oxide dismutase (SOD) | 0.124 | 0.072 | 154.42 ± 4.25 | 243.25 ± 6.48 | +57.52 |
| Peroxidase (POX) | 1.811 | 3.832 | 23.88 ± 0.43 | 45.37 ± 0.30 | +90.01 |
| Root Na+/K+ | 1.878 | 0.058 | 1.66 ± 0.07 | 3.36 ± 0.10 | +102.54 |
| Shoot Na+/K+ | 5.786 | 2.536 | 1.20 ± 0.05 | 2.84 ± 0.07 | +136.61 |
| Bulb Na+/K+ | 0.120 | 0.058 | 0.23 ± 0.02 | 0.33 ± 0.02 | +41.54 |
| Plant height (PH) | 1.140 | 0.217 | 43.24 ± 0.61 | 40.89 ± 0.68 | −5.45 |
| Number of leaves/plant (NL) | 0.653 | 0.088 | 8.47 ± 0.10 | 8.32 ± 0.13 | −1.79 |
| Bulb diameter (BD) | 0.462 | 0.640 | 5.38 ± 0.11 | 4.73 ± 0.09 | −12.04 |
| Bulb yield/plant (BYP) | 73.662 | 78.800 | 60.33 ± 0.29 | 42.23 ± 0.23 | −30.01 |
| Total soluble solids (TSS) | 6.193 | 0.849 | 13.09 ± 0.11 | 12.00 ± 0.14 | −8.30 |
| Dependent Variable | Step and Variables | R2 | F-Stat. | Probability |
|---|---|---|---|---|
| BYP (Bulb yield/plant) | 1. BD | 0.805 | 165.014 | 0.000 |
| 2. BD + PH | 0.872 | 132.617 | 0.000 | |
| 3. BD + PH + APX | 0.890 | 102.647 | 0.000 | |
| 4. BD + PH + APX + gS | 0.926 | 115.676 | 0.000 | |
| 5. BD + PH + APX + gS + POX | 0.944 | 122.048 | 0.000 | |
| 6. BD + PH + APX + gS + POX + CAT | 0.961 | 141.819 | 0.000 | |
| 7. BD + PH + APX + gS + POX + CAT + MDA | 0.967 | 140.697 | 0.000 | |
| 8. BD + PH + APX + gS + POX + CAT + MDA + MSI | 0.972 | 141.141 | 0.000 | |
| 9. BD + PH + APX + gS + POX + CAT + MDA + MSI + Bulb Na+/K+ | 0.976 | 143.261 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanwal, S.K.; Kesh, H.; Kumar, A.; Dubey, B.K.; Khar, A.; Rouphael, Y.; Kumar, P. Salt Tolerance Potential in Onion: Confirmation through Physiological and Biochemical Traits. Plants 2022, 11, 3325. https://doi.org/10.3390/plants11233325
Sanwal SK, Kesh H, Kumar A, Dubey BK, Khar A, Rouphael Y, Kumar P. Salt Tolerance Potential in Onion: Confirmation through Physiological and Biochemical Traits. Plants. 2022; 11(23):3325. https://doi.org/10.3390/plants11233325
Chicago/Turabian StyleSanwal, Satish Kumar, Hari Kesh, Arvind Kumar, Bhanu Kumar Dubey, Anil Khar, Youssef Rouphael, and Pradeep Kumar. 2022. "Salt Tolerance Potential in Onion: Confirmation through Physiological and Biochemical Traits" Plants 11, no. 23: 3325. https://doi.org/10.3390/plants11233325
APA StyleSanwal, S. K., Kesh, H., Kumar, A., Dubey, B. K., Khar, A., Rouphael, Y., & Kumar, P. (2022). Salt Tolerance Potential in Onion: Confirmation through Physiological and Biochemical Traits. Plants, 11(23), 3325. https://doi.org/10.3390/plants11233325