
Pollen Production of Selected Grass Species in Russia and India at the Levels of Anther, Flower and Inflorescence

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling Areas
2.2. Species Identification
2.3. Estimation of Pollen Production
3. Results
3.1. ITS1-2 Sequencing
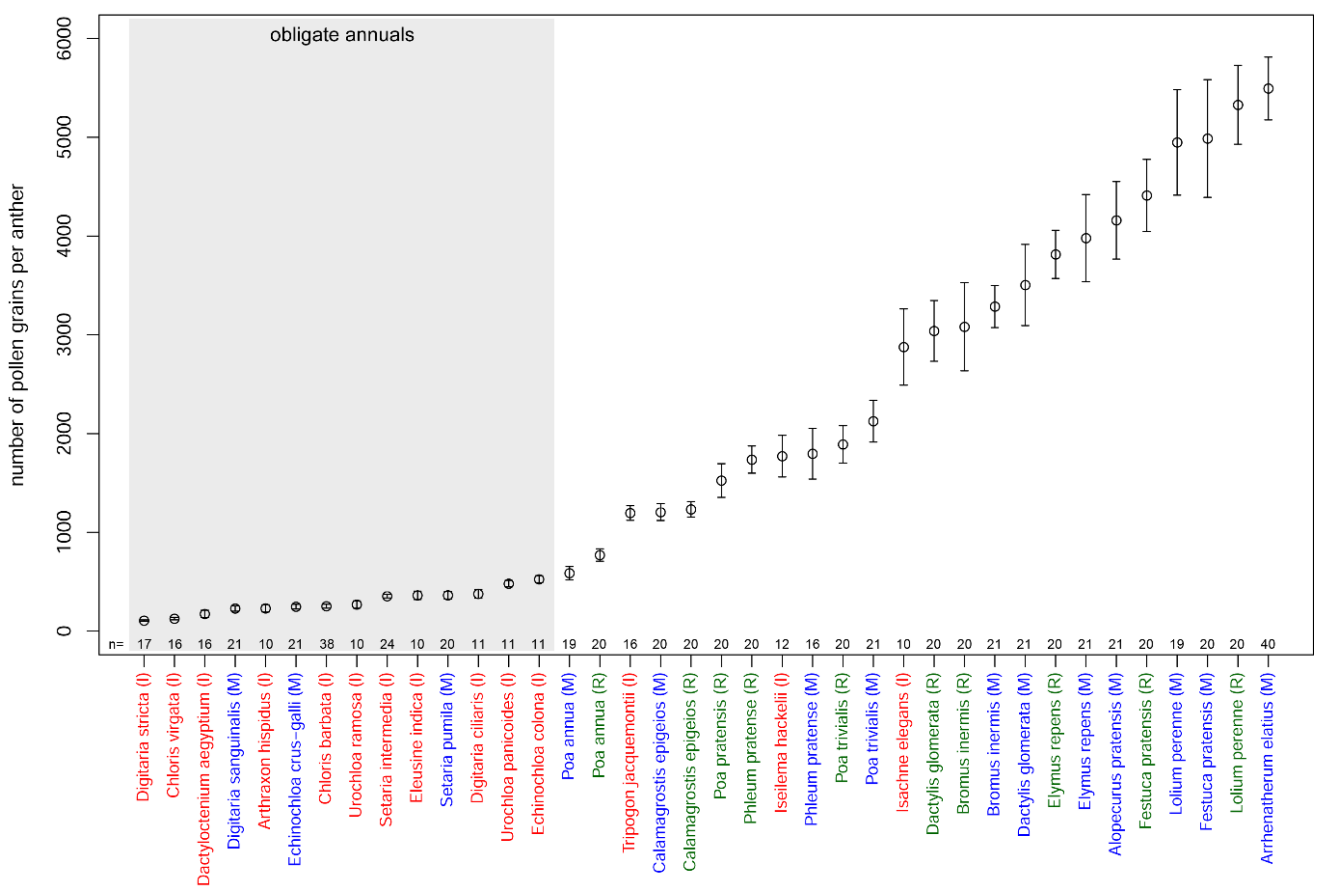
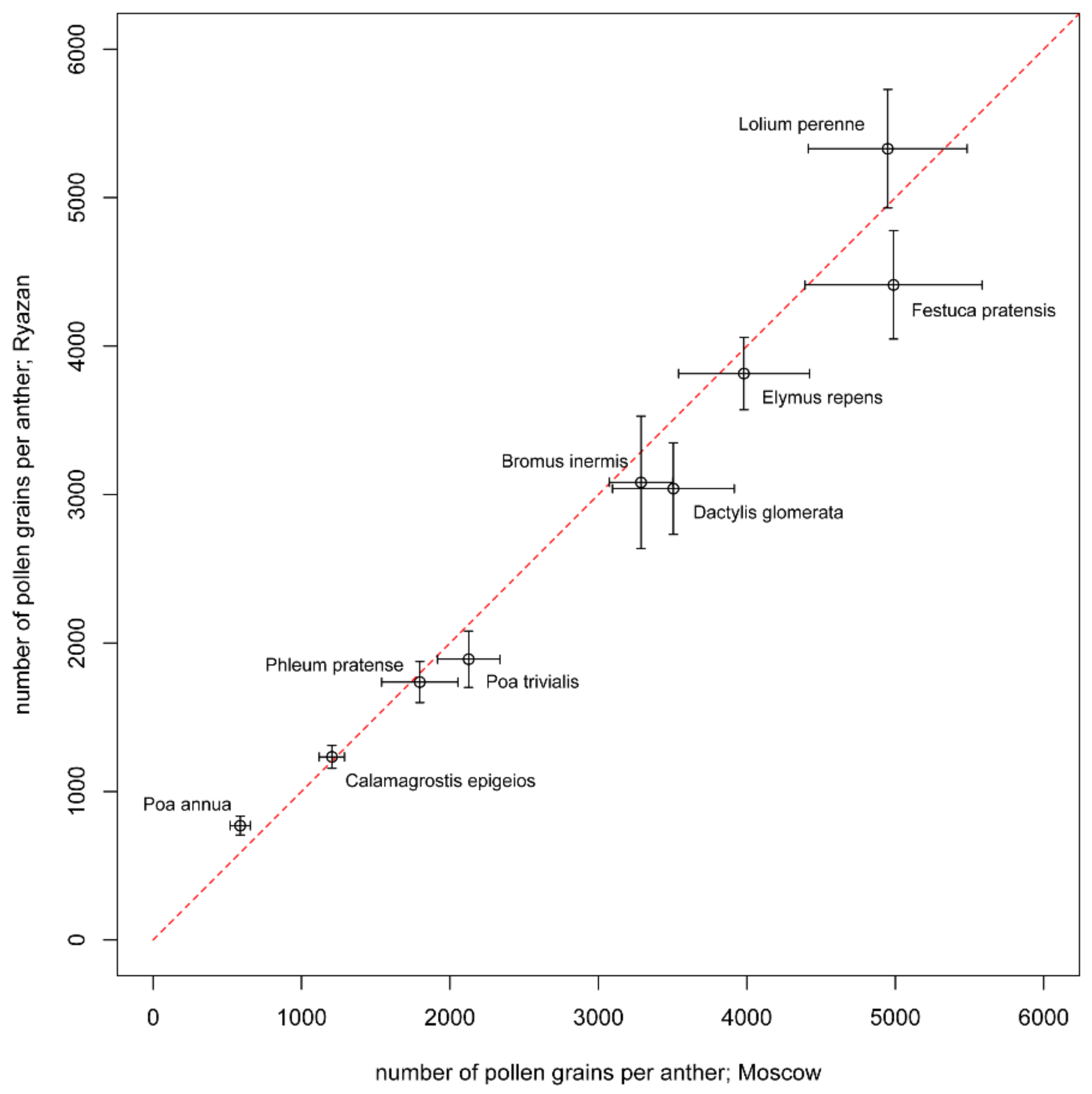
3.2. Number of Pollen Grains per Anther
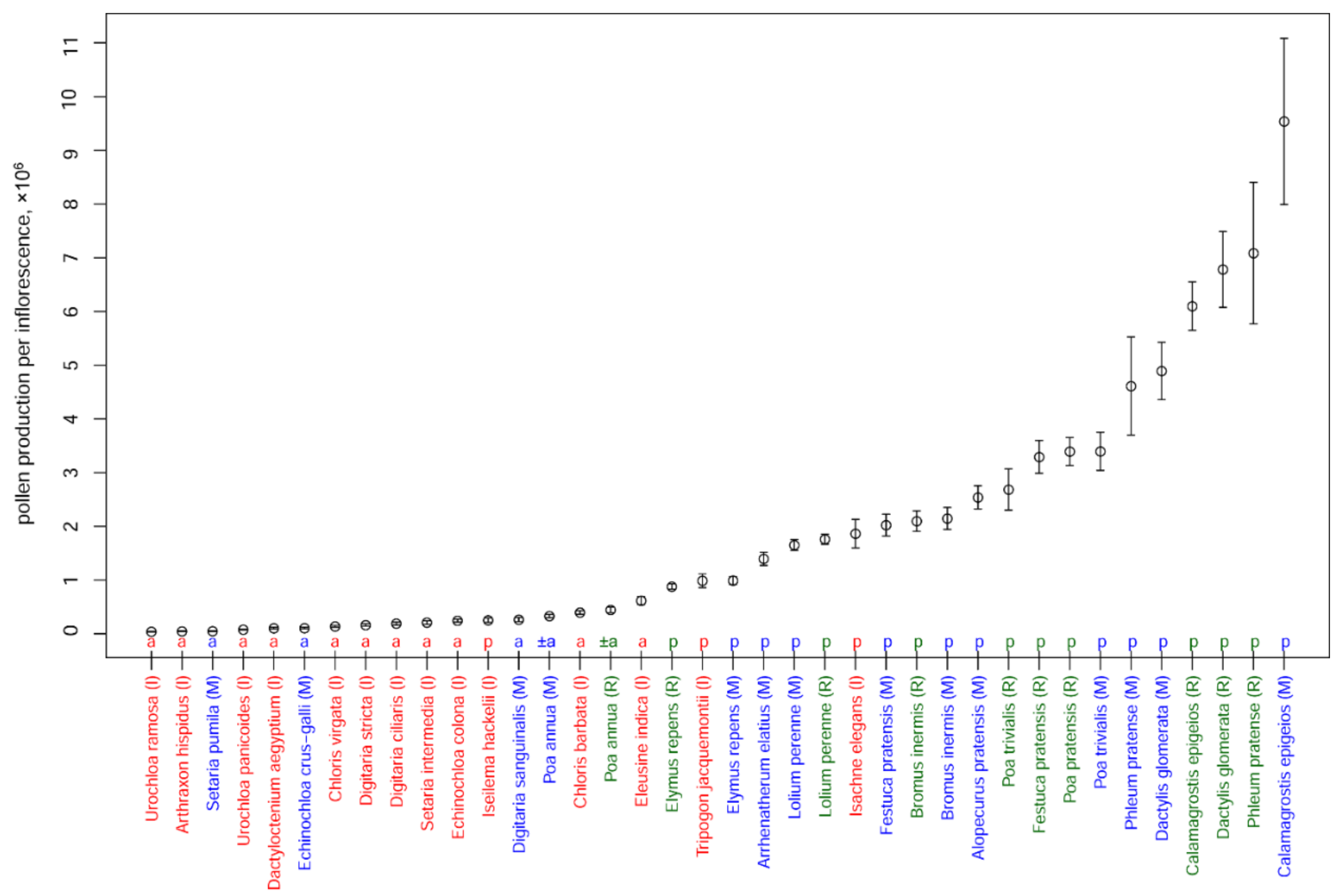
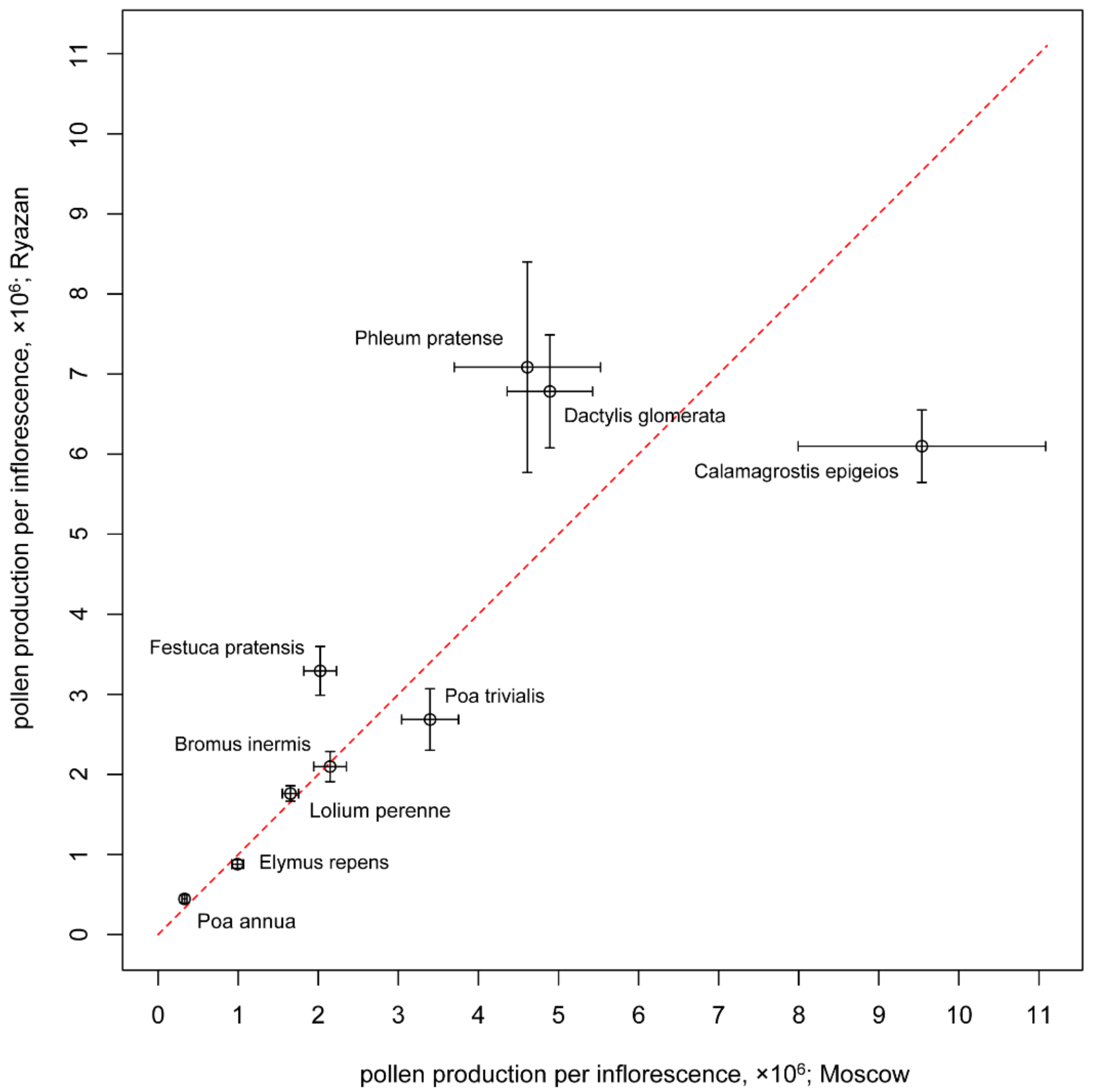
3.3. Pollen Production per Inflorescence
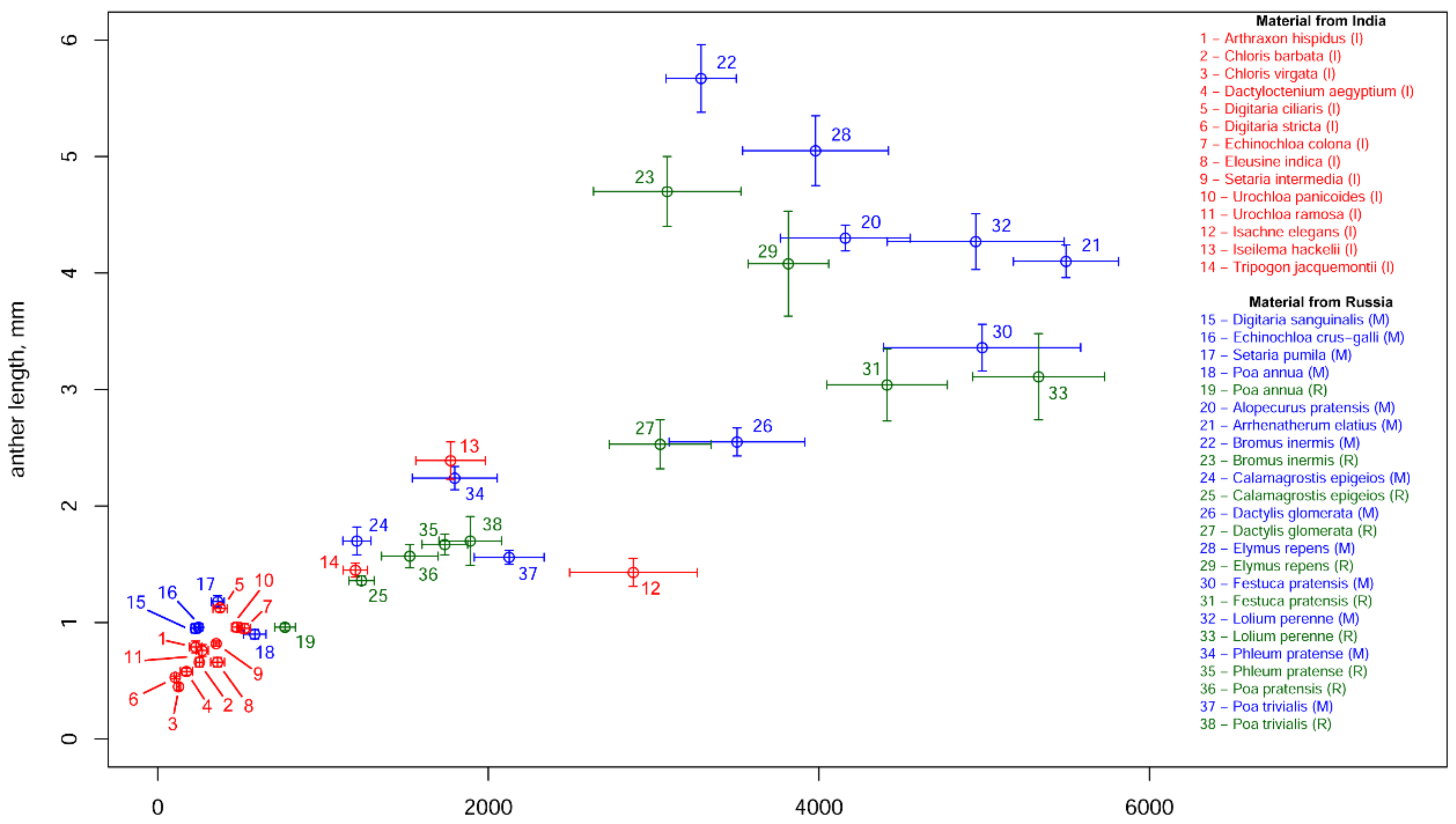
3.4. Anther Length, Diameter of Pollen Grains and Pollen Production per Anther
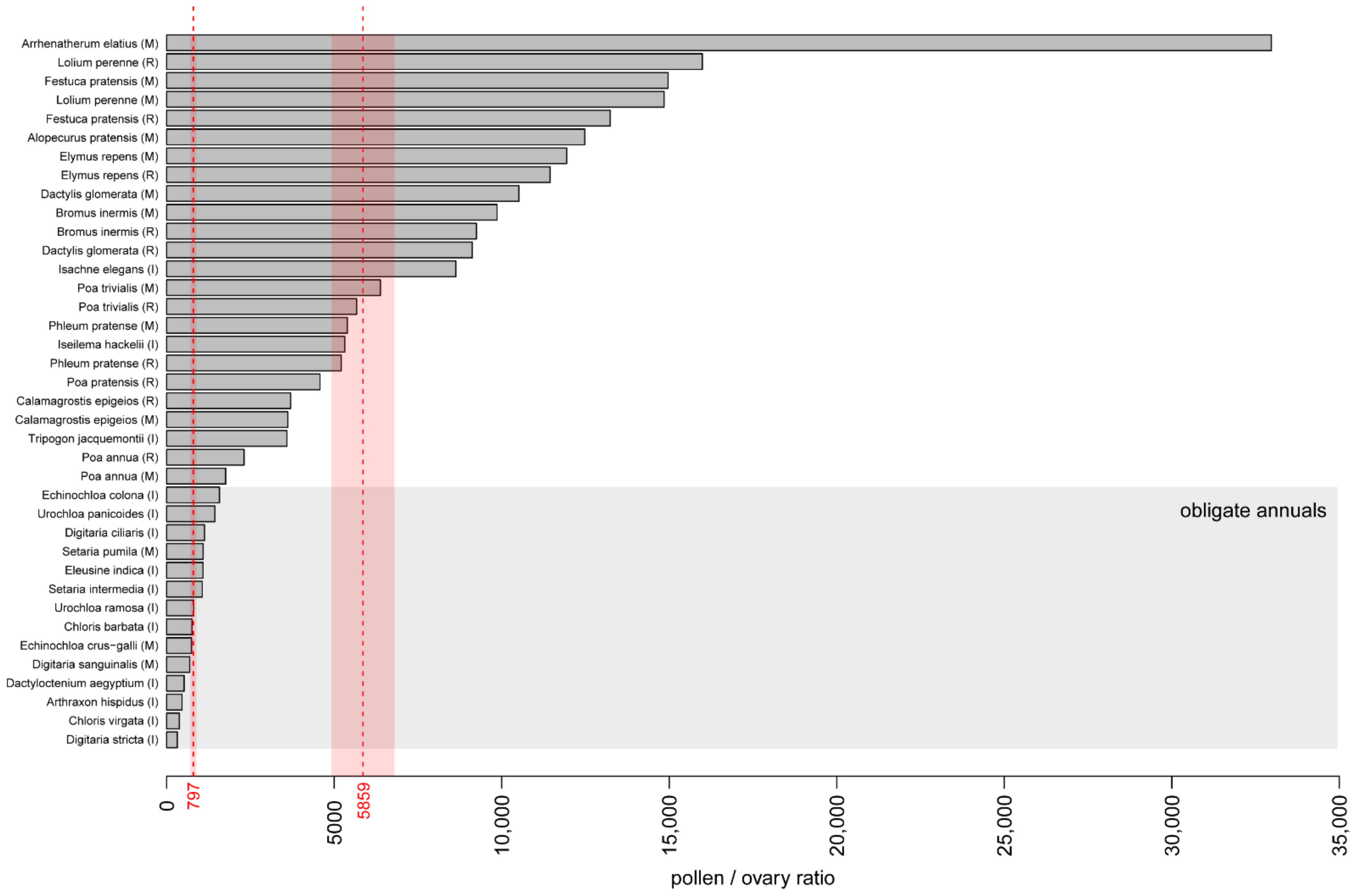
3.5. Pollen/Ovule Ratio
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Teisher, J.K.; Clark, L.G.; Barberá, P.; Gillespie, L.J.; Zuloaga, F.O. A worldwide phylogenetic classification of the Poaceae (Gramineae) II: An update and a comparison of two 2015 classifications. J. Syst. Evol. 2017, 55, 259–290. [Google Scholar] [CrossRef] [Green Version]
- McKevith, B. Nutritional aspects of cereals. Nutr. Bull. 2004, 29, 111–142. [Google Scholar] [CrossRef]
- Laskowski, W.; Górska-Warsewicz, H.; Rejman, K.; Czeczotko, M.; Zwolińska, J. How important are cereals and cereal products in the average polish diet? Nutrients 2019, 11, 679. [Google Scholar] [CrossRef] [Green Version]
- Watson, L. The grass family, Poaceae. In Reproductive Versatility in the Grasses; Cambridge University Press: Cambridge, UK, 1990; pp. 1–31. [Google Scholar]
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Liccardi, G.; Popov, T.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy 2007, 62, 976–990. [Google Scholar] [CrossRef] [PubMed]
- García-Mozo, H. Poaceae pollen as the leading aeroallergen worldwide: A review. Allergy 2017, 72, 1849–1858. [Google Scholar] [CrossRef] [Green Version]
- Andersson, K.; Lidholm, J. Characteristics and immunobiology of grass pollen allergens. Int. Arch. Allergy Immunol. 2003, 130, 87–107. [Google Scholar] [CrossRef]
- Linder, H.P.; Rudall, P.J. Evolutionary history of Poales. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 107–124. [Google Scholar] [CrossRef]
- Kellogg, E.A. Poaceae. In The Families and Genera of Vascular Plants; Springer: Berlin/Heidelberg, Germany, 2015; Volume XIII, pp. 1–416. [Google Scholar]
- Adams, D.E.; Perkins, W.E.; Estes, J.R. Pollination systems in Paspalum dilatatum Poir. (Poaceae): An example of insect pollination in a temperate grass. Am. J. Bot. 1981, 68, 389–394. [Google Scholar] [CrossRef]
- Campbell, C.S.; Quinn, J.A.; Cheplick, G.P.; Bell, T.J. Cleistogamy in grasses. Annu. Rev. Ecol. Syst. 1983, 14, 411–441. [Google Scholar] [CrossRef]
- Chhabra, A.K.; Sethi, S.K. Inheritance of cleistogamic flowering in durum wheat (Triticum durum). Euphytica 1991, 55, 147–150. [Google Scholar] [CrossRef]
- Ueno, K.; Itoh, H. Cleistogamy in wheat: Genetic control and the effect of environmental conditions. Cereal Res. Commun. 1997, 25, 185–189. [Google Scholar] [CrossRef]
- Connor, H.E. Breeding systems in the grasses: A survey. N. Z. J. Bot. 1979, 17, 547–574. [Google Scholar] [CrossRef]
- Heslop-Harrison, J. The function of the glume pit and the control of cleistogamy in Bothriochloa decipiens (Hack.) C.E. Hubbard. Phytomorphology 1961, 11, 378–383. [Google Scholar]
- Knox, R.B.; Heslop-Harrison, J. Experimental control of aposporous apomixis in a grass of the Andropogoneae. Bot. Not. 1963, 116, 127141. [Google Scholar]
- Quarin, C.L. Seasonal changes in the incidence of apomixis of diploid, triploid, and tetraploid plants of Paspalum cromyorrhizon. Euphytica 1986, 35, 515–522. [Google Scholar] [CrossRef]
- Soderstrom, T.R.; Calderón, C.E. Insect pollination in tropical rain forest grasses. Biotropica 1971, 3, 1–16. [Google Scholar] [CrossRef]
- Janzen, D.H. Why bamboos wait so long to flower. Annu. Rev. Ecol. Syst. 1976, 7, 347–391. [Google Scholar] [CrossRef]
- Jackson, G.C.; Woodbury, R.O. Host plants of the carpenter bee, Xylocopa brasilianorum L. (Hymenoptera: Apoidea) in Puerto Rico. J. Agric. Univ. Puerto Rico 1976, 60, 639–660. [Google Scholar] [CrossRef]
- Wong, K.M. Flowering, fruiting and germination of the bamboo Schizostachyum zollingeri in Perlis. Malays. For. 1981, 44, 453–463. [Google Scholar]
- Bogdan, A.V. Grass pollination by bees in Kenya. Proc. Linnean Soc. 1962, 173, 57–60. [Google Scholar] [CrossRef]
- Koshy, K.C.; Harikumar, D.; Narendran, T.C. Insect visits to some bamboos of the Western Ghats, India. Curr. Sci. 2001, 81, 833–838. [Google Scholar]
- Wolowski, M.; Freitas, L. An overview on pollination of the neotropical Poales. Rodriguésia 2015, 66, 329–336. [Google Scholar] [CrossRef]
- Ruiz-Sanchez, E.; Castro-Castro, A.; Clark, L.G. Chusquea septentrionalis sp. nov. (Poaceae: Bambusoideae) from the Madrean region in Durango, Mexico. Nord. J. Bot. 2017, 35, 546–551. [Google Scholar] [CrossRef]
- Saunders, M.E. Insect pollinators collect pollen from wind-pollinated plants: Implications for pollination ecology and sustainable agriculture. Insect Conserv. Divers. 2018, 11, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Baena, J.C.; Hidalgo, P.J.; Domínguez, E.; Galán, C. Pollen production in the Poaceae family. Grana 2003, 42, 153–159. [Google Scholar] [CrossRef]
- Aboulaich, N.; Bouziane, H.; Kadiri, M.; del Mar Trigo, M.; Riadi, H.; Kazzaz, M.; Merzouki, A. Pollen production in anemophilous species of the Poaceae family in Tetouan (NW Morocco). Aerobiologia 2009, 25, 27–38. [Google Scholar] [CrossRef]
- Ghitarrini, S.; Galán, C.; Frenguelli, G.; Tedeschini, E. Phenological analysis of grasses (Poaceae) as a support for the dissection of their pollen season in Perugia (central Italy). Aerobiologia 2017, 33, 339–349. [Google Scholar] [CrossRef]
- Cruden, R.W. Pollen-ovule ratios: A conservative indicator of breeding systems in flowering plants. Evolution 1977, 31, 32–46. [Google Scholar] [CrossRef]
- Subba Reddi, C.S.; Reddi, N.S. Pollen production in some anemophilous angiosperms. Grana 1986, 25, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Kaybeleva, E.I.; Yudakova, O.I. Pollen-ovule ratios in cereals with different mode of reproduction. Bull. Saratov Univ. Bot. Gard. 2015, 13, 148–154. [Google Scholar]
- Bai, A.J.; Subba Reddi, C.S. Pollen productivity and pollen incidence in some potentially allergenic plants of Visakhapatnam. Adv. Pollen Spore Res. 1980, 5, 217–224. [Google Scholar]
- McKone, M.J. Intraspecific variation in pollen yield in bromegrass (Poaceae: Bromus). Am. J. Bot. 1989, 76, 231–237. [Google Scholar] [CrossRef]
- De Vries, A.P. Flowering biology of wheat, particularly in view of hybrid seed production—A review. Euphytica 1971, 20, 152–170. [Google Scholar] [CrossRef]
- Sapra, V.T.; Hughes, J.L. Pollen production in hexaploid Triticale. Euphytica 1975, 24, 237–243. [Google Scholar] [CrossRef]
- Nguyen, V.; Fleury, D.; Timmins, A.; Laga, H.; Hayden, M.; Mather, D.; Okada, T. Addition of rye chromosome 4R to wheat increases anther length and pollen grain number. Theor. Appl. Genet. 2015, 128, 953–964. [Google Scholar] [CrossRef]
- Song, X.; Feng, J.; Cui, Z.; Zhang, C.; Sun, D. Genome-wide association study for anther length in some elite bread wheat germplasm. Czech J. Genet. Plant Breed. 2018, 54, 109–114. [Google Scholar]
- De Vries, A.P. Some aspects of cross-pollination in wheat (Triticum aestivum L.). 3. Anther length and number of pollen grains per anther. Euphytica 1974, 23, 11–19. [Google Scholar] [CrossRef]
- Smart, I.J.; Tuddenham, W.G.; Knox, R.B. Aerobiology of grass pollen in the city atmosphere of Melbourne: Effects of weather parameters and pollen sources. Aust. J. Bot. 1979, 27, 333–342. [Google Scholar] [CrossRef]
- Piotrowska, K. Pollen production in selected species of anemophilous plants. Acta Agrobot. 2008, 61, 41–52. [Google Scholar] [CrossRef]
- Tormo-Molina, R.; Maya-Manzano, J.-M.; Silva-Palacios, I.; Fernández-Rodríguez, S.; Gonzalo-Garijo, Á. Flower production and phenology in Dactylis glomerata. Aerobiologia 2015, 31, 469–479. [Google Scholar] [CrossRef]
- Driessen, M.N.B.M.; Willemse, M.T.M.; Van Luijn, J.A.G. Grass pollen grain determination by light- and UV-microscopy. Grana 1989, 28, 115–122. [Google Scholar] [CrossRef]
- Weber, R.W. Patterns of pollen cross-allergenicity. J. Allergy Clin. Immunol. 2003, 112, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Kraaijeveld, K.; De Weger, L.A.; Ventayol García, M.; Buermans, H.; Frank, J.; Hiemstra, P.S.; Den Dunnen, J.T. Efficient and sensitive identification and quantification of airborne pollen using next-generation DNA sequencing. Mol. Ecol. Resour. 2015, 15, 8–16. [Google Scholar] [CrossRef]
- Bell, K.L.; De Vere, N.; Keller, A.; Richardson, R.T.; Gous, A.; Burgess, K.S.; Brosi, B.J. Pollen DNA barcoding: Current applications and future prospects. Genome 2016, 59, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Ghitarrini, S.; Pierboni, E.; Rondini, C.; Tedeschini, E.; Tovo, G.R.; Frenguelli, G.; Albertini, E. New biomolecular tools for aerobiological monitoring: Identification of major allergenic Poaceae species through fast real-time PCR. Ecol. Evol. 2018, 8, 3996–4010. [Google Scholar] [CrossRef] [PubMed]
- Leontidou, K.; Vernesi, C.; De Groeve, J.; Cristofolini, F.; Vokou, D.; Cristofori, A. DNA metabarcoding of airborne pollen: New protocols for improved taxonomic identification of environmental samples. Aerobiologia 2018, 34, 63–74. [Google Scholar] [CrossRef]
- Christensen, J.E.; Horner, H.T., Jr. Pollen pore development and its spatial orientation during microsporogenesis in the grass Sorghum bicolor. Am. J. Bot. 1974, 61, 604–623. [Google Scholar] [CrossRef]
- Kirpes, C.C.; Clark, L.G.; Lersten, N.R. Systematic significance of pollen arrangement in microsporangia of Poaceae and Cyperaceae. Review and observations on representative taxa. Am. J. Bot. 1996, 83, 1609–1622. [Google Scholar] [CrossRef]
- Furness, C.A.; Rudall, P.J. The tapetum and systematics in monocotyledons. Bot. Rev. 1998, 64, 201–239. [Google Scholar] [CrossRef]
- Friedman, J.; Harder, L.D. Functional associations of floret and inflorescence traits among grass species. Am. J. Bot. 2005, 92, 1862–1870. [Google Scholar] [CrossRef] [PubMed]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [Green Version]
- Potdar, G.G.; Salunkhe, C.B.; Yadav, S.R. Grasses of Maharashtra; Shivaji University: Kolhapur, India, 2012; ISBN 81-8486-450-7. [Google Scholar]
- Alexeev, Y.E. The family Poaceae. In Maevskyi P.F. Flora of the Middle Part of European Russia; KMK: Moscow, Russia, 2014; pp. 509–569. ISBN 5-04-099221-1. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Adobe Systems Inc. Adobe Photoshop, Version 22.5.1; Adobe Systems Inc.: Mountain View, CA, USA, 2021. [Google Scholar]
- Connor, H.E. Flowers and floral biology of the holy grasses (Hierochloe and Anthoxanthum: Aveneae, Gramineae). Flora 2012, 207, 323–333. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Stanley, R.G.; Linskens, H.F. Pollen: Biology, Biochemistry, Management; Springer: Berlin/Heidelberg, Germany, 1974; ISBN 978-642-65907-2. [Google Scholar]
- Beri, S.M.; Anand, S.C. Factors affecting pollen shedding capacity in wheat. Euphytica 1971, 20, 327–332. [Google Scholar] [CrossRef]
- Erbar, C.; Langlotz, M. Pollen to ovule ratios: Standard or variation-a Compilation. Bot. Jahrb. Syst. 2004, 126, 71–132. [Google Scholar] [CrossRef]
- Smith, D.C. Pollination and seed formation in grasses. J. Agric. Res. 1944, 68, 79–95. [Google Scholar]
- Lehmann, C.; Rebele, F. Zum potential sexueller Fortpflanzung bei Calamagrostis epigejos (L.) Roth. Verh. Ges. Ökol. 1994, 23, 445–450. [Google Scholar]
- Rebele, F.; Lehmann, C. Biological flora of Central Europe: Calamagrostis epigejos (L.) Roth. Flora 2001, 196, 325–344. [Google Scholar] [CrossRef]
- Aiken, S.G.; Dore, W.G.; Lefkovitch, L.P.; Armstrong, K.C. Calamagrostis epigejos (Poaceae) in North America, especially Ontario. Can. J. Bot. 1989, 67, 3205–3218. [Google Scholar] [CrossRef]
- Thoiba, K.; Pradeep, A.K. A Revision of Tripogon (Poaceae: Chloridoideae) in India. Rheedea 2020, 30, 325. [Google Scholar] [CrossRef]
- Gaff, D.F.; Bole, P.V. Resurrection grasses in India. Oecologia 1986, 71, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Baumann, U.; Juttner, J.; Bian, X.; Langridge, P. Self-incompatibility in the grasses. Ann. Bot. 2000, 85, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Pfitzenmeyer, C.D.C. Arrhenatherum elatius (L.) J. &. C. Presl (A. avenaceum Beauv.). J. Ecol. 1962, 50, 235–245. [Google Scholar]
- Cuguen, J.; Acheroy, M.; Loutfi, A.L.; Petit, D.; Vernet, P. Breeding system differentiation in Arrhenatherum elatius populations: Evolution toward selfing. Evol Trends Plants 1989, 3, 17–24. [Google Scholar]
- Petit, C.; Lesbros, P.; Ge, X.; Thompson, J.D. Variation in flowering phenology and selfing rate across a contact zone between diploid and tetraploid Arrhenatherum elatius (Poaceae). Heredity 1997, 79, 31–40. [Google Scholar] [CrossRef]
- Jürgens, A.; Witt, T.; Gottsberger, G. Pollen grain numbers, ovule numbers and pollen-ovule ratios in Caryophylloideae: Correlation with breeding system, pollination, life form, style number, and sexual system. Sex. Plant Reprod. 2002, 14, 279–289. [Google Scholar]
- Galloni, M.; Podda, L.; Vivarelli, D.; Cristofolini, G. Pollen presentation, pollen-ovule ratios, and other reproductive traits in Mediterranean legumes (fam. Fabaceae-subfam. Faboideae). Plant Syst. Evol. 2007, 266, 147–164. [Google Scholar] [CrossRef]
- Alarcón, M.L.; Roquet, C.; Aldasoro, J.J. Evolution of pollen/ovule ratios and breeding system in Erodium (Geraniaceae). Syst. Bot. 2011, 36, 661–676. [Google Scholar] [CrossRef]
- Ramirez, N.; Seres, A. Plant reproductive biology of herbaceous monocots in a Venezuelan tropical cloud forest. Plant Syst. Evol. 1994, 190, 129–142. [Google Scholar] [CrossRef]
- Chorghe, A.R.; Tiwari, A.P. Effective lectotypification of three names in the genus Iseilema (Poaceae: Panicoideae). Kew Bull. 2021, 76, 857–858. [Google Scholar] [CrossRef]
- Chandramohan, K.; Mahesh, Y. A taxonomic revision of the genus Iseilema Andersson (Poaceae) in India. J. Global Biosci. 2020, 9, 7836–7868. [Google Scholar]
- Kabeer, K.A.A.; Nair, V.J. Flora of Tamil Nadu—Grasses; Botanical Survey of India: Kolkata, India, 2009; pp. 1–524. ISBN 81-8177-031-5.
- Hutchinson, C.S.; Seymour, G.B. Poa annua L. J. Ecol. 1982, 70, 887–901. [Google Scholar] [CrossRef]
- Bell, C.R. Mineral nutrition and flower to flower pollen size variation. Am. J. Bot. 1959, 46, 621–624. [Google Scholar] [CrossRef]
- Lau, T.-C.; Stephenson, A.G. Effects of soil nitrogen on pollen production, pollen grain size, and pollen performance in Cucurbita pepo (Cucurbitaceae). Am. J. Bot. 1993, 80, 763–768. [Google Scholar] [CrossRef]
- Severova, E.E.; Yadav, S.R.; Sokoloff, D.D. Pollen morphology of Indian Aponogeton (Aponogetonaceae, Alismatales) and the problem of recognizing palynotypes in a taxonomically diverse and ancient genus. Phytotaxa 2020, 475, 187–200. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher Information and GenBank Accession Numbers for ITS1-2 Sequences | Locality, Sampling Date | Life Form |
|---|---|---|---|
| Alopecurus pratensis L. | Severova s.n. (MW 1072500) | Lomonosov Moscow State University, Moscow, Russia, 28 May 2021 | perennial |
| Arrhenatherum elatius (L.) P. Beauv. ex J. & C. Presl. | Severova s.n. (MW 1072498) | as above, 11 June 2021 | perennial |
| Arthraxon hispidus (Thunb.) Makino | Sokoloff et al. 127 (MW 0758387), OL752458 | Shivaji University, Botanical Garden, Kolhapur, India, 31 August 2019 | annual |
| Bromus inermis Leyss. | Severova s.n. (MW 1072504) | 1. Lomonosov Moscow State University, Moscow, Russia, 11 June 2021 2. Sovetskyi rayion, Ryazan, Russia, 8 June 2021 | perennial |
| Calamagrostis epigeios (L.) Roth | Severova s.n. (MW 1072496) | 1. as above, 9 July 2021 2. Deulino, Ryazan’ oblast, Russia, 5 July 2020 | perennial |
| Chloris barbata Sw. | Sokoloff et al. 87 (MW 0758380); OL752448 | Shivaji University, Botanical Garden, Kolhapur, India, 29 August 2019 | annual |
| Chloris virgata Sw. | Sokoloff et al. 93 (MW 0758383); OL752452 | as above, 29 August 2019 | annual |
| Dactylis glomerata L. | Severovas.n. (MW 1072497) | 1. Lomonosov Moscow State University, Moscow, Russia, 28 May 2021 2. Sovetskyi rayion, Ryazan, Russia, 9 June 2020 | perennial |
| Dactyloctenium aegyptium (L.) Willd. | Sokoloff et al. 88 (MW 0758384); OL752449 | Shivaji University, Botanical Garden, Kolhapur, India, 29 August 2019 | annual |
| Digitaria ciliaris (Retz.) Koeler | Sokoloff et al. 92 (MW 0758386); OL752451 | as above, 29 August 2019 | annual |
| Digitaria sanguinalis (L.) Scop. | Severova s.n. (MW 1072506) | Lomonosov Moscow State University, Moscow, Russia, 4 October 2021 | annual |
| Digitaria stricta Roth ex Roem. & Schult. | Sokoloff et al. 98 (MW 0758385); OL752455 ** | Shivaji University, Botanical Garden, Kolhapur, India, 29 August 2019 | annual |
| Echinochloa colona (L.) Link | Sokoloff et al. 86 (MW 0758390); OL752447 | as above, 29 August 2019 | annual |
| Echinochloa crus-galli (L.) P. Beauv. | Severova s.n. (MW 1072505) | Marfino, Moscow, Russia, 4 October 2021 | annual |
| Eleusine indica (L.) Gaertn. | Sokoloff et al. 97 (MW 0758388); OL752454 | Shivaji University, Botanical Garden, Kolhapur, India, 29 August 2019 | annual |
| Elymus repens (L.) Gould | Severova s.n. (MW 1072493) | 1. Lomonosov Moscow State University, Moscow, Russia, 28 May 2021 2. Deulino, Ryazan oblast, Russia, 10 June 2020 | perennial |
| Festuca pratensis Huds. | Severova s.n. (MW 1072495) | 1. as above, 28 May 2021 2. Sovetskyi rayion, Ryazan, Russia, 10 June 2020 | perennial |
| Isachne elegans Dalzell | Sokoloff et al. 156 (MW 0758391); OL752459 ** | Western Ghats, Maharashtra State, India, 2 September 2019 | perennial |
| Iseilema hackelii Shrestha et Gandhi | Sokoloff et al. 108 (MW 0758392); OL752456 * | Western Ghats, Maharashtra State, Satara District, India, 30 August 2019 | perennial |
| Lolium perenne L. | Severova s.n. (MW 1072494) | 1. Lomonosov Moscow State University, Moscow, Russia, 28 May 2021 2. Sovetskyi rayion, Ryazan, Russia, 12 June 2020 | perennial |
| Phleum pratense L. | Severova s.n. (MW 1072502) | 1. as above, 28 May 2021 2. as above, 12 June 2020 | perennial |
| Poa annua L. | Severova s.n. (MW 1072501) | 1. as above, 28 May 2021 2. as above, 6 June 2020 | annual or short-living perennial |
| Poa pratensis L. | Selezneva, Karaseva s.n. (MW 1072499) | Sovetskyi rayion, Ryazan, Russia, 10 June 2020 | perennial |
| Poa trivialis L. | Severova s.n. (MW) | 1. Lomonosov Moscow State University, Moscow, Russia, 28 May 2021 2. Ryazan, Russia, 10 June 2020 | perennial |
| Setaria intermedia Roem. & Schult. | Sokoloff et al. 95 (MW 0758381); OL752453 * | Shivaji University, Botanical Garden, Kolhapur, India, 29 August 2019 | annual |
| Setaria pumila (Poir.) Roem. & Schult. | Severova s.n. (MW 1072503) | Lomonosov Moscow State University, Moscow, Russia, 4 October 2021 | annual |
| Tripogon jacquemontii Stapf | Sokoloff et al. 111 (MW 0758389); OL752457 | Western Ghats, Maharashtra State, Satara District, India, 30 August 2019 | perennial |
| Urochloa panicoides P. Beauv. | Sokoloff et al. 85 (MW 0758379); OL752446 * | Shivaji University, Botanical Garden, Kolhapur, India, 29 August 2019 | annual |
| Urochloa ramosa (L.) T.Q. Nguyen | Sokoloff et al. 90 (MW 0758382); OL752450 | as above, 29 August 2019 | annual |
| Species | Pollen Production per Anther | Number of Flowers per Spikelet | Number of Spikelets per Inflorescence | Pollen Production per Inflorescence, ×1000 | P/O | Anther Length, mm | Diameter of Pollen Grain, μm |
|---|---|---|---|---|---|---|---|
| Alopecurus pratensis (M) | 4160 ± 393 (20.7) | 1 ± 0 (0) | 203 ± 17.4 (17.7) | 2539 ± 217 (27.5) | 12,480 ± 1179 (20.7) | 4.3 ± 0.11 (8.7) | 30.08 ± 0.77 (6.03) |
| Arrhenatherum elatius (M) | 5495 ± 318 (18.1) | 2 ± 0 (0) | 42 ± 7.3 (25.8) | 1397 ± 122 (31.9) | 32,970 ± 1908 (18.1) * | 4.1 ± 0.14 (7.5) | 35.44 ± 0.93 (6.79) |
| Arthraxon hispidus (I) | 230 ± 37 (22.4) | 1 ± 0 (0) | 101 ± 10 (15) | 46 ± 5 (27.2) | 460 ± 74 (22.4) | 0.79 ± 0.05 (8.6) | 26.9 ± 2 (14.5) |
| Bromus inermis (M) | 3287 ± 213 (14.2) | 5.2 ± 0.3 (17.6) | 42 ± 6.9 (35.2) | 2148 ± 203 (42.7) | 9861 ± 639 (14.2) | 5.67 ± 0.29 (12.3) | 38.84 ± 1.48 (8.35) |
| Bromus inermis (R) | 3082 ± 446 (30.9) | 6.6 ± 0.3 (17.4) | 35 ± 5.3 (32.9) | 2098 ± 189 (50.1) | 9246 ± 1338 (30.9) | 4.7 ± 0.3 (13.4) | 47.52 ± 1.44 (6.49) |
| Calamagrostis epigeios (M) | 1205 ± 85 (15.1) | 1 ± 0 (0) | 2638 ± 856 (42.2) | 9538 ± 1546 (45.3) | 3615 ± 255 (15.1) | 1.7 ± 0.12 (18.2) | 23.69 ± 1.06 (10.83) |
| Calamagrostis epigeios (R) | 1233 ± 77 (13.4) | 1 ± 0 (0) | 1648 ± 151.5 (19.6) | 6099 ± 452 (23.9) | 3699 ± 231 (13.4) | 1.36 ± 0.04 (7.6) | 23.36 ± 0.5 (5.17) |
| Chloris barbata (I) | 252 ± 20 (24.4) | 1 ± 0 (0) | 524 ± 75.4 (20.1) | 397 ± 33 (32) | 756 ± 60 (24.4) | 0.66 ± 0.04 (16.1) | 23.82 ± 1.1 (15.47) |
| Chloris virgata (I) | 126 ± 12 (18) | 1 ± 0 (0) | 365 ± 82.8 (31.8) | 138 ± 17 (37) | 378 ± 36 (18) | 0.45 ± 0.03 (13.5) | 24.11 ± 1.15 (16.64) |
| Dactylis glomerata (M) | 3505 ± 411 (25.8) | 2.6 ± 0.3 (23.5) | 180 ± 19.5 (23.1) | 4893 ± 533 (43.1) | 10,515 ± 1233 (25.8) | 2.55 ± 0.12 (11.3) | 30.56 ± 1.2 (8.45) |
| Dactylis glomerata (R) | 3040 ± 308 (21.7) | 3.5 ± 0.3 (27.8) | 211 ± 36.4 (36.9) | 6783 ± 706 (53.1) | 9120 ± 924 (21.7) | 2.53 ± 0.21 (17.9) | 38.6 ± 2.19 (12.45) |
| Dactyloctenium aegyptium (I) | 174 ± 37 (39.5) | 2.9 ± 0.2 (10.9) | 69 ± 7.5 (15.4) | 103 ± 15 (44.4) | 522 ± 111 (39.5) | 0.58 ± 0.03 (14.7) | 28.23 ± 0.96 (14.2) |
| Digitaria ciliaris (I) | 377 ± 43 (17) | 1 ± 0 (0) | 169 ± 44.5 (36.9) | 191 ± 28 (41.1) | 1131 ± 129 (17) | 1.12 ± 0.03 (4.1) | 28.44 ± 1.27 (12.39) |
| Digitaria sanguinalis (M) | 229 ± 28 (27.3) | 1 ± 0 (0) | 385 ± 69.8 (38.7) | 264 ± 39 (48.6) | 687 ± 84 (27.3) | 0.95 ± 0.04 (8.5) | 30.63 ± 1.65 (12.75) |
| Digitaria stricta (I) | 106 ± 7 (13.4) | 1 ± 0 (0) | 511 ± 141.7 (38.8) | 162 ± 22 (41.3) | 318 ± 21 (13.4) | 0.53 ± 0.01 (6.1) | 26.53 ± 0.81 (11.04) |
| Echinochloa colona (I) | 525 ± 36 (10.2) | 1 ± 0 (0) | 155 ± 26 (25.3) | 244 ± 27 (27.4) | 1575 ± 108 (10.2) | 0.95 ± 0.04 (5.3) | 35 ± 1.5 (7) |
| Echinochloa crus-galli (M) | 247 ± 24 (20.5) | 1 ± 0 (0) | 142 ± 23 (50.6) | 105 ± 15 (55.6) | 741 ± 72 (20.5) | 0.96 ± 0.03 (7.2) | 28.57 ± 1.57 (16.52) |
| Eleusine indica (I) | 362 ± 42 (16.3) | 5.2 ± 0.5 (12.2) | 109 ± 20.9 (26.7) | 618 ± 75 (34.1) | 1086 ± 126 (16.3) | 0.66 ± 0.04 (9.6) | 21.93 ± 0.87 (12.22) |
| Elymus repens (M) | 3980 ± 441 (24.3) | 4.3 ± 0.2 (22.4) | 19 ± 1.5 (16.9) | 991 ± 73 (37.9) | 11,940 ± 1323 (24.3) | 5.05 ± 0.3 (13.5) | 32.87 ± 0.84 (5.91) |
| Elymus repens (R) | 3815 ± 244 (13.6) | 4.7 ± 0.2 (16) | 16 ± 1.6 (20.3) | 879 ± 51 (29.6) | 11,445 ± 732 (13.6) | 4.08 ± 0.45 (25) | 33.23 ± 0.82 (5.98) |
| Festuca pratensis (M) | 4988 ± 596 (25.5) | 4.8 ± 0.4 (29.9) | 28 ± 3.9 (29.7) | 2024 ± 205 (51.3) | 14,964 ± 1788 (25.5) | 3.36 ± 0.2 (12.5) | 31.04 ± 1.19 (9.72) |
| Festuca pratensis (R) | 4412 ± 365 (17.7) | 7.3 ± 0.4 (19.7) | 34 ± 5.9 (37.3) | 3293 ± 305 (47) | 13,236 ± 1095 (17.7) | 3.04 ± 0.31 (22.5) | 31.21 ± 1.44 (9.83) |
| Isachne elegans (I) | 2877 ± 386 (18.7) | 2 ± 0 (0) | 108 ± 26.7 (34.6) | 1864 ± 266 (39.9) | 8631 ± 1158 (18.7) | 1.43 ± 0.12 (11.7) | 22.29 ± 1.37 (16.55) |
| Iseilema hackelii (I) | 1772 ± 211 (18.8) | 1 ± 0 (0) | 47 ± 17.6 (51.1) | 253 ± 41 (67.4) | 5316 ± 633 (18.8) ** | 2.39 ± 0.16 (11.3) | 23.79 ± 1.06 (13.32) |
| Lolium perenne (M) | 4949 ± 535 (22.4) | 6.5 ± 0.3 (20.7) | 17 ± 0.8 (13.9) | 1653 ± 103 (34.1) | 14,847 ± 1605 (22.4) | 4.27 ± 0.24 (12.6) | 34.01 ± 1.06 (6.68) |
| Lolium perenne (R) | 5329 ± 399 (16) | 6.5 ± 0.3 (19.3) | 17 ± 0.9 (11.2) | 1761 ± 96 (27.8) | 15,987 ± 1197 (16) | 3.11 ± 0.37 (25.4) | 42.63 ± 2.56 (12.84) |
| Phleum pratense (M) | 1797 ± 257 (26.9) | 1 ± 0 (0) | 855 ± 210.1 (52.5) | 4612 ± 913 (60.6) | 5391 ± 771 (26.9) | 2.24 ± 0.1 (11.1) | 37.46 ± 1.03 (8) |
| Phleum pratense (R) | 1737 ± 138 (17) | 1 ± 0 (0) | 1360 ± 360.4 (56.6) | 7086 ± 1315 (59.9) | 5211 ± 414 (17) | 1.67 ± 0.09 (14.8) | 31.72 ± 1.32 (9.17) |
| Poa annua (M) | 587 ± 68 (23.9) | 4.2 ± 0.3 (23.4) | 45 ± 6.5 (30.8) | 330 ± 31 (47) | 1761 ± 204 (23.9) | 0.9 ± 0.04 (11.2) | 25.23 ± 1.12 (10.97) |
| Poa annua (R) | 770 ± 63 (17.6) | 3.7 ± 0.3 (23.8) | 34 ± 7.8 (43.6) | 445 ± 51 (54.4) | 2310 ± 189 (17.6) | 0.96 ± 0.03 (7) | 27.5 ± 0.75 (6.06) |
| Poa pratensis (R) | 1525 ± 171 (24) | 4 ± 0.2 (16.7) | 187 ± 21.5 (24.6) | 3394 ± 260 (39.1) | 4575 ± 513 (24) | 1.57 ± 0.1 (13.8) | 32.25 ± 1.69 (10.87) |
| Poa trivialis (M) | 2126 ± 211 (21.8) | 2.1 ± 0.1 (20.1) | 257 ± 47 (34.4) | 3396 ± 355 (46.7) | 6378 ± 633 (21.8) | 1.56 ± 0.06 (9.6) | 23.6 ± 1.83 (18.72) |
| Poa trivialis (R) | 1891 ± 190 (21.4) | 2.5 ± 0.2 (32.5) | 189 ± 50.6 (57.1) | 2686 ± 385 (73) | 5673 ± 570 (21.4) | 1.7 ± 0.21 (27) | 23.24 ± 0.88 (9.23) |
| Setaria intermedia (I) | 354 ± 18 (12.4) | 1 ± 0 (0) | 197 ± 75.6 (53.6) | 209 ± 34 (55.4) | 1062 ± 54 (12.4) | 0.82 ± 0.02 (5.8) | 23.29 ± 0.62 (11.2) |
| Setaria pumila (M) | 363 ± 39 (22.9) | 1 ± 0 (0) | 45 ± 9.3 (44.5) | 49 ± 8 (51.1) | 1089 ± 117 (22.9) | 1.18 ± 0.05 (9.6) | 32.55 ± 0.91 (9.38) |
| Tripogon jacquemontii (I) | 1196 ± 74 (11.6) | 9 ± 2.3 (35.1) | 31 ± 2.7 (10.5) | 989 ± 129 (38.9) | 3588 ± 222 (11.6) | 1.45 ± 0.06 (7.8) | 22.98 ± 1.11 (17.26) |
| Urochloa panicoides (I) | 480 ± 33 (10.3) | 1 ± 0 (0) | 54 ± 5 (23.4) | 77 ± 6 (25.7) | 1440 ± 99 (10.3) | 0.96 ± 0.04 (7.3) | 24.5 ± 0.8 (5.6) |
| Urochloa ramosa (I) | 269 ± 37 (19.1) | 1 ± 0 (0) | 49 ± 11 (36.7) | 40 ± 7 (41.9) | 807 ± 111 (19.1) | 0.76 ± 0.05 (7.7) | 27.5 ± 3.1 (18.6) |
| This Study: I, India; M, Moscow; R, Ryazan | Prieto-Baena et al. [27] | Aboulaich et al. [28] | Kaybeleva, Yudakova [32] | Tormo-Molina et al. [42] | Bai, Reddy [33] | Smart et al. [40] | Subba Reddi, Reddi [31] | Cruden [30] | McKone [34] | |
|---|---|---|---|---|---|---|---|---|---|---|
| Bromus inermis | 9861 (M) 9246 (R) | 11,913 | ||||||||
| Chloris barbata | 756 (I) | 837 | 945 | |||||||
| Dactylis glomerata | 10,515 (M) 9120 (R) | 10,425 | 6429 | 9240 | 5431 | 3900 | ||||
| Dactyloctenium aegyptium | 522 (I) | 555 | ||||||||
| Digitaria ciliaris | 1131 (I) | 1125 | ||||||||
| Digitaria sanguinalis | 687 (I) | 1234 | ||||||||
| Echinochloa crus-gali | 741 (I) | 1267 | ||||||||
| Eleusine indica | 1086 (I) | 810 | 1111 | |||||||
| Elymus repens | 11,940 (M) 11,445 (R) | 16,230 | ||||||||
| Festuca pratensis | 14,964 (M) 13,236 (R) | 5151 | ||||||||
| Lolium perenne | 14,847 (M) 15,987 (R) | 16,200 | ||||||||
| Poa annua | 1761 (M) 2310 (R) | 1022 | 1216 | 5523 | ||||||
| Poa pratensis | 4575 (R) | 3735 | ||||||||
| Poa trivialis | 6378 (M) 5673 (R) | 3386 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Severova, E.; Kopylov-Guskov, Y.; Selezneva, Y.; Karaseva, V.; Yadav, S.R.; Sokoloff, D. Pollen Production of Selected Grass Species in Russia and India at the Levels of Anther, Flower and Inflorescence. Plants 2022, 11, 285. https://doi.org/10.3390/plants11030285
Severova E, Kopylov-Guskov Y, Selezneva Y, Karaseva V, Yadav SR, Sokoloff D. Pollen Production of Selected Grass Species in Russia and India at the Levels of Anther, Flower and Inflorescence. Plants. 2022; 11(3):285. https://doi.org/10.3390/plants11030285
Chicago/Turabian StyleSeverova, Elena, Yury Kopylov-Guskov, Yulia Selezneva, Vera Karaseva, Shrirang R. Yadav, and Dmitry Sokoloff. 2022. "Pollen Production of Selected Grass Species in Russia and India at the Levels of Anther, Flower and Inflorescence" Plants 11, no. 3: 285. https://doi.org/10.3390/plants11030285
APA StyleSeverova, E., Kopylov-Guskov, Y., Selezneva, Y., Karaseva, V., Yadav, S. R., & Sokoloff, D. (2022). Pollen Production of Selected Grass Species in Russia and India at the Levels of Anther, Flower and Inflorescence. Plants, 11(3), 285. https://doi.org/10.3390/plants11030285