
The Role of Biostimulants as Alleviators of Biotic and Abiotic Stresses in Grapevine: A Review




Abstract
:1. Introduction
2. Impacts of Climate Change on Grapevine
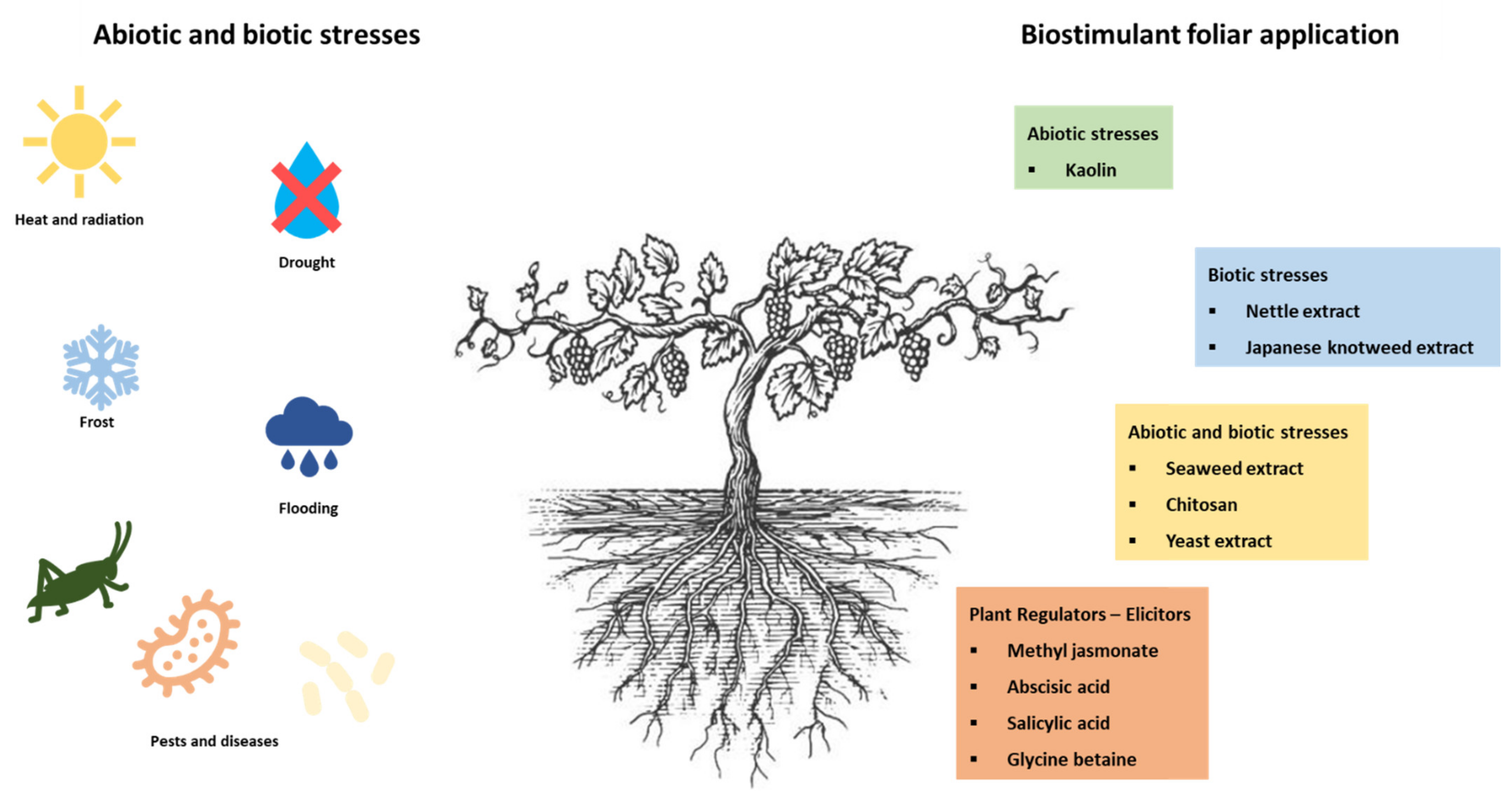
3. Use of Biostimulants as a Mitigation Strategy for Biotic and Abiotic Stresses in Grapevine
4. Biostimulants Used in Grapevine
4.1. Biostimulants as Abiotic Stress Alleviators
Kaolin
4.2. Biostimulants as Biotic Stress Alleviators
4.2.1. Nettle
4.2.2. Japanese Knotweed
4.3. Biostimulants as Abiotic and Biotic Stress Alleviators
4.3.1. Seaweed Extracts
4.3.2. Chitosan
4.3.3. Yeast Extracts
4.4. Biostimulants as Plant Regulators and Elicitors
4.4.1. Methyl Jasmonate
4.4.2. Abscisic Acid
4.4.3. Salicylic Acid
4.4.4. Glycine Betaine
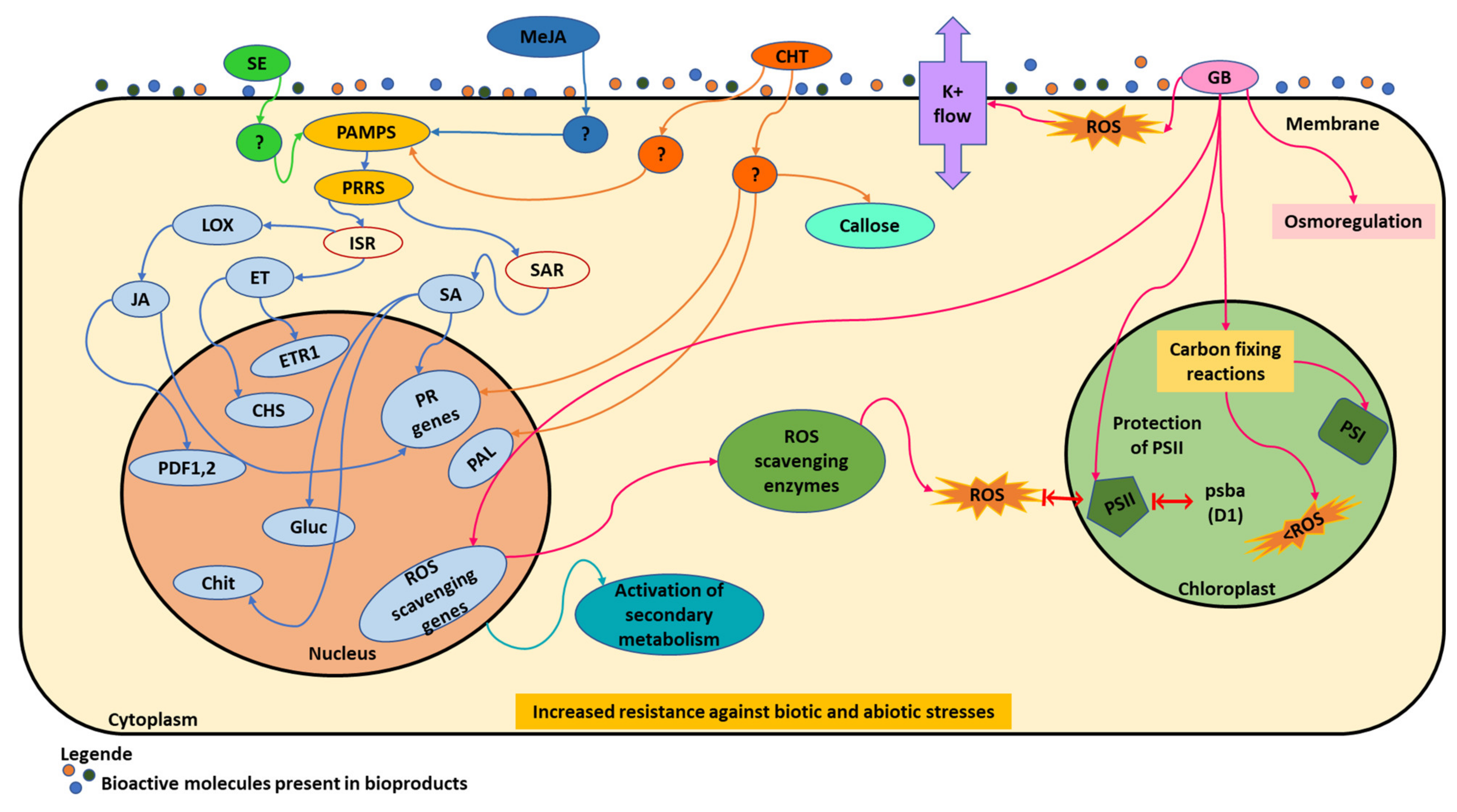
5. Grapevine Responses to Biostimulants
Differential Expression of Genes Involved in Metabolic Grapevine Pathways upon Biostimulants Application
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vivier, M.A.; Pretorius, I.S. Genetically tailored grapevines for the wine industry. Trends Biotechnol. 2002, 20, 472–478. [Google Scholar] [CrossRef]
- Conde, A.; Pimentel, D.; Neves, A.; Dinis, L.-T.; Bernardo, S.; Correia, C.M.; Gerós, H.; Moutinho-Pereira, J. Kaolin foliar application has a stimulatory effect on phenylpropanoid and flavonoid pathways in grape berries. Front. Plant Sci. 2016, 7, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, M.; Jain, S.; Bhardwaj, A.; Nagpal, R.; Puniya, M.; Tomar, R.; Singh, V.; Parkash, O.; Prasad, G.B.K.S.; Marotta, F.; et al. Biological and medicinal properties of grapes and their bioactive constituents: An update. J. Med. Food 2009, 12, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Schwaninger, H.R.; Baldo, A.M.; Labate, J.A.; Zhong, G.Y.; Simon, C.J. A phylogenetic analysis of the grape genus (vitis l.) reveals broad reticulation and concurrent diversification during neogene and quaternary climate change. BMC Evol. Biol. 2013, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Pilati, S.; Brazzale, D.; Guella, G.; Milli, A.; Ruberti, C.; Biasioli, F.; Zottini, M.; Moser, C. The onset of grapevine berry ripening is characterized by ROS accumulation and lipoxygenase-mediated membrane oeroxidation in the skin. BMC Plant Biol. 2014, 14, 87. [Google Scholar] [CrossRef] [Green Version]
- OIV State of the World Vitivinicultural Sector in 2020; International Organisation of Vine and Wine: Paris, France, 2021; Available online: https://www.oiv.int/public/medias/7909/oiv-state-of-the-world-vitivinicultural-sector-in-2020.pdf (accessed on 8 October 2021).
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum—Based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, N. Tratado de Viticultura—A Videira, a Vinha e o “terroir”; Fer, C., Ed.; Esfera Poética: Lisboa, Portugal, 2008; ISBN 978-972-8987-15-2. [Google Scholar]
- OIV Definition of Vitivinicultural “Terroir”; The International Organisation of Vine and Wine: Paris, France, 2010; Available online: https://www.oiv.int/en/technical-standards-and-documents/resolutions-of-the-oiv/viticulture-resolutions (accessed on 7 November 2021).
- Garrido, A.; Serôdio, J.; De Vos, R.; Conde, A.; Cunha, A. Influence of foliar kaolin application and irrigation on photosynthetic activity of grape berries. Agronomy 2019, 9, 685. [Google Scholar] [CrossRef] [Green Version]
- Jones, G. Uma Avaliação do Clima para a Região Demarcada do Douro: Uma Análise das Condições Climáticas do Passado, Presente e Futuro Para a Produção de Vinho; Alves, F., Moriondo, M., Ferrise, R., Santos, J., Malheiro, A., Eds.; ADVID-As: Vila Real, Portugal, 2013; ISBN 978-989-98368-0-8. [Google Scholar]
- Fraga, H.; De Cortázar Atauri, I.G.; Malheiro, A.C.; Moutinho-Pereira, J.; Santos, J.A. Viticulture in Portugal: A review of recent trends and climate change projections. OENO One 2017, 51, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Jamiołkowska, A. Natural compounds as elicitors of plant resistance against diseases and new biocontrol strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef] [Green Version]
- van Loon, L.C.; Bakker, P.A.H.M.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [Green Version]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Allakhverdiev, S.I.; Hurry, V.; Hüner, N.P.A. Stress-related hormones and glycine betaine interplay in protection of photosynthesis under abiotic stress conditions. Photosynth. Res. 2015, 126, 221–235. [Google Scholar] [CrossRef]
- Le Mire, G.; Nguyen, M.L.; Fassotte, B.; Du Jardin, P.; Verheggen, F.; Delaplace, P.; Haissam Jijakli, M. Implementing plant biostimulants and biocontrol strategies in the agroecological management of cultivated ecosystems. A Rev. Biotechnol. Agron. Soc. Environ. 2016, 20, 299–313. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 201, 213–221. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Romanazzi, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. A review of the use of biostimulants in the vineyard for improved grape and wine quality: Effects on prevention of grapevine diseases. J. Sci. Food Agric. 2019, 99, 1001–1009. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress-a review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Gamboa, G.; Moreno-Simunovic, Y. Seaweeds in viticulture: A review focused on grape quality. Cienc. E Tec. Vitivinic. 2021, 36, 9–21. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Darriet, P. The impact of climate change on viticulture and wine quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Change 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Santos, J.A. An overview of climate change impacts on European viticulture. Food Energy Secur. 2013, 1, 94–110. [Google Scholar] [CrossRef]
- Gambetta, J.M.; Holzapfel, B.P.; Stoll, M.; Friedel, M. Sunburn in grapes: A review. Front. Plant Sci. 2021, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Reis, S.; Fraga, H.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Rodrigues, P.; Santos, J.A. Grapevine phenology in four Portuguese wine regions: Modeling and predictions. Appl. Sci. 2020, 10, 3708. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.V. Climate change in the western united states grape growing regions. Acta Hortic. 2005, 689, 41–60. [Google Scholar] [CrossRef]
- Delaunois, B.; Farace, G.; Jeandet, P.; Clément, C.; Baillieul, F.; Dorey, S.; Cordelier, S. Elicitors as alternative strategy to pesticides in grapevine? Current knowledge on their mode of action from controlled conditions to vineyard. Environ. Sci. Pollut. Res. 2013, 21, 4837–4846. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
- Wong, F.P.; Wilcox, W.F. Distribution of baseline sensitivities to azoxystrobin among isolates of Plasmopara viticola. Plant Dis. 2000, 84, 275–281. [Google Scholar] [CrossRef]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant-Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- Belhadj, A.; Saigne, C.; Telef, N.; Cluzet, S.; Bouscaut, J.; Corio-Costet, M.F.; Mérillon, J.M. Methyl jasmonate induces defense responses in grapevine and triggers protection against Erysiphe necator. J. Agric. Food Chem. 2006, 54, 9119–9125. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; Mancini, V.; Carrasco-Quiroz, M.; Servili, A.; Gutiérrez-Gamboa, G.; Foglia, R.; Pérez-Álvarez, E.P.; Romanazzi, G. Chitosan and laminarin as alternatives to copper for Plasmopara viticola control: Effect on grape amino acid. J. Agric. Food Chem. 2017, 65, 7379–7386. [Google Scholar] [CrossRef]
- Rantsiou, K.; Giacosa, S.; Pugliese, M.; Englezos, V.; Ferrocino, I.; Río Segade, S.; Monchiero, M.; Gribaudo, I.; Gambino, G.; Gullino, M.L.; et al. Impact of chemical and alternative fungicides applied to grapevine cv Nebbiolo on microbial ecology and chemical-physical grape characteristics at harvest. Front. Plant Sci. 2020, 11, 700. [Google Scholar] [CrossRef]
- Colombo, M.; Masiero, S.; Rosa, S.; Caporali, E.; Toffolatti, S.L.; Mizzotti, C.; Tadini, L.; Rossi, F.; Pellegrino, S.; Musetti, R.; et al. NoPv1: A synthetic antimicrobial peptide aptamer targeting the causal agents of grapevine downy mildew and potato late blight. Sci. Rep. 2020, 10, 17574. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Petit, A.; Wojnarowiez, G.; Panon, M.; Baillieul, F.; Clément, C.; Fontaine, F.; Vaillant-gaveau, N. Botryticides affect grapevine leaf photosynthesis without inducing defense mechanisms. Planta 2009, 229, 497–506. [Google Scholar] [CrossRef]
- Jermini, M.; Blaise, P.; Gessler, C. Quantification of the influence of the downy mildew (Plasmopara viticola) epidemics on the compensatory capacities of Vitis vinifera “Merlot” to limit the qualitative yield damage. Vitis—J. Grapevine Res. 2010, 49, 153–160. [Google Scholar]
- Godard, S.; Slacanin, I.; Viret, O.; Gindro, K. Induction of defence mechanisms in grapevine leaves by emodin- and anthraquinone-rich plant extracts and their conferred resistance to downy mildew. Plant Physiol. Biochem. 2009, 47, 827–837. [Google Scholar] [CrossRef]
- Njogu, M.; Nyankanga, R.; Muthomi, J.; Muindi, E. Studies on the effects of stinging nettle extract, phosphoric acid and conventional fungicide combinations on the management of potato late blight and tuber yield in the highlands of Kenya. J. Agric. Food Sci. 2014, 2, 119–127. [Google Scholar]
- Rodino, S.; Butu, M.; Butu, A. Alternative antimicrobial formula for plant protection. Bull. USAMV Ser. Agric. 2018, 75, 32–37. [Google Scholar] [CrossRef]
- Hildebrandt, U.; Marsell, A.; Riederer, M. Direct Effects of physcion, chrysophanol, emodin, and pachybasin on germination and appressorium formation of the barley (Hordeum vulgare L.) powdery mildew fungus Blumeria graminis f. sp. hordei (DC.). Speer. J. Agric. Food Chem. 2018, 66, 3393–3401. [Google Scholar] [CrossRef] [PubMed]
- Ghazal, H.N.; Al-Shahwany, A.W.; Al-Dulaimy, F.T. Control of gray mold on tomato plants by spraying Piper nigrum and Urtica dioica extracts under greenhouse condition. Iraqi J. Sci. 2019, 60, 961–971. [Google Scholar] [CrossRef]
- Dinis, L.-T.; Ferreira, H.; Pinto, G.; Bernardo, S.; Correia, C.M.; Moutinho-Pereira, J. Kaolin-based, foliar reflective film protects photosystem II structure and function in grapevine leaves exposed to heat and high solar radiation. Photosynthetica 2016, 54, 47–55. [Google Scholar] [CrossRef]
- Dinis, L.T.; Bernardo, S.; Conde, A.; Pimentel, D.; Ferreira, H.; Félix, L.; Gerós, H.; Correia, C.M.; Moutinho-Pereira, J. Kaolin exogenous application boosts antioxidant capacity and phenolic content in berries and leaves of grapevine under summer stress. J. Plant Physiol. 2016, 191, 45–53. [Google Scholar] [CrossRef]
- Taskos, D.; Stamatiadis, S.; Yvin, J.C.; Jamois, F. Effects of an Ascophyllum nodosum (L.) Le Jol. extract on grapevine yield and berry composition of a Merlot vineyard. Sci. Hortic. 2019, 250, 27–32. [Google Scholar] [CrossRef]
- Salvi, L.; Brunetti, C.; Cataldo, E.; Niccolai, A.; Centritto, M.; Ferrini, F.; Mattii, G.B. Effects of Ascophyllum nodosum extract on Vitis vinifera: Consequences on plant physiology, grape quality and secondary metabolism. Plant Physiol. Biochem. 2019, 139, 21–32. [Google Scholar] [CrossRef]
- Frioni, T.; Tombesi, S.; Quaglia, M.; Calderini, O.; Moretti, C.; Poni, S.; Gatti, M.; Moncalvo, A.; Sabbatini, P.; Berrìos, J.G.; et al. Metabolic and transcriptional changes associated with the use of Ascophyllum nodosum extracts as tools to improve the quality of wine grapes (Vitis vinifera cv. Sangiovese) and their tolerance to biotic stress. J. Sci. Food Agric. 2019, 99, 6350–6363. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.T.; Dumas, B. An Ulva armoricana extract protects plants against three powdery mildew pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar] [CrossRef]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan oligomers and copper sulfate induce grapevine defense reactions and resistance to gray mold and downy mildew. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan stimulates defense reactions in grapevine leaves and inhibits development of Botrytis cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Romanazzi, G.; Landi, L.; Feliziani, E. Innovative strategies based on the use of biostimulants to manage plant diseases and minimize the application of synthetic fungicides in grapevine and stone fruits. Med. Jadertina 2019, 49, 34. [Google Scholar]
- Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.; Inês, A.; Oliveira, A.; Falco, V. Chitosan upregulates the genes of the ROS pathway and enhances the antioxidant potential of grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) yissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan application in vineyards (Vitis vinifera L. cv. Tinto Cão) induces accumulation of anthocyanins and other phenolics in berries, mediated by modifications in the transcription of secondary metabolism genes. Int. J. Mol. Sci. 2020, 21, 306. [Google Scholar] [CrossRef] [Green Version]
- Górnik, K.; Grzesik, M.; Romanowska-Duda, B. The Effect of chitosan on rooting of grapevine cuttings and on subsequent plant growth under drought and temperature stress. J. Fruit Ornam. Plant Res. 2008, 16, 333–343. [Google Scholar]
- Vitalini, S.; Ruggiero, A.; Rapparini, F.; Neri, L.; Tonni, M.; Iriti, M. The application of chitosan and benzothiadiazole in vineyard (Vitis vinifera L. cv Groppello Gentile) changes the aromatic profile and sensory attributes of wine. Food Chem. 2014, 162, 192–205. [Google Scholar] [CrossRef]
- Zamani, M.M.; Rabiei, V.; Nejatian, M.A.; Taheri, M. Effect of proline and glycine betaine application on some physiological characteristics in grapevine under drought stress. J. Crop. Improv. 2014, 26, 247–258. [Google Scholar] [CrossRef]
- Jalil, O.T.J.; Sabır, A. Changes in leaf and shoot water statutes of grapevines in response to contrasting water availability and glycine betaine pulverization. Int. J. Agric. Environ. Food Sci. 2017, 1, 20–26. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Fernández-Fernández, J.I.; Crespo-Villegas, O.; Garde-Cerdán, T. Elicitors used as a tool to increase stilbenes in grapes and wines. Food Res. Int. 2017, 98, 34–39. [Google Scholar] [CrossRef]
- Yamaguchi, I.; Cohen, J.D.; Culler, A.H.; Quint, M.; Slovin, J.P.; Nakajima, M.; Yamaguchi, S.; Sakakibara, H.; Kuroha, T.; Hirai, N.; et al. Plant hormones. In Comprehensive Natural Products II; Liu, H.-W., Mander, L.B.T.-C.N.P.I.I., Eds.; Elsevier: Oxford, UK, 2010; pp. 9–125. ISBN 978-0-08-045382-8. [Google Scholar]
- Gil-Muñoz, R.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Improving phenolic and chromatic characteristics of Monastrell, Merlot and Syrah wines by using methyl jasmonate and benzothiadiazole. OENO One 2017, 51, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.L.; Liu, M.; Zhao, H.; Meng, J.F.; Fang, Y.L. Effect of exogenous abscisic acid and methyl jasmonate on anthocyanin composition, fatty acids, and volatile compounds of Cabernet Sauvignon (Vitis vinifera L.) grape berries. Molecules 2016, 21, 1354. [Google Scholar] [CrossRef] [Green Version]
- Abdel Salam, M. Effect of foliar application of salicylic acid and micronutrients on the berries quality of “Bez El Naka” local grape cultivar. Middle East J. Appl. Sci. 2016, 6, 178–188. [Google Scholar]
- Blanch, G.P.; Gómez-Jiménez, M.C.; del Castillo, M.L.R. Exogenous salicylic acid improves phenolic content and antioxidant activity in table grapes. Plant Foods Hum. Nutr. 2020, 75, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Kraeva, E.; Andary, C.; Carbonneau, A.; Deloire, A. Salicylic acid treatment of grape berries retards ripening. Vitis 1998, 37, 143–144. [Google Scholar]
- Portu, J.; López, R.; Santamaría, P.; Garde-Cerdán, T. Elicitation with methyl jasmonate supported by precursor feeding with phenylalanine: Effect on Garnacha grape phenolic content. Food Chem. 2017, 237, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Lacroux, F.; Trégoat, O.; Van Leeuwen, C.; Pons, A.; Tominaga, T.; Lavigne-Cruège, V.; Dubourdieu, D. Effect of foliar nitrogen and sulphur application on aromatic expression of Vitis vinifera L. cv. Sauvignon Blanc. OENO One 2008, 42, 125. [Google Scholar] [CrossRef] [Green Version]
- Portu, J.; López-Alfaro, I.; Gómez-Alonso, S.; López, R.; Garde-Cerdán, T. Changes on grape phenolic composition induced by grapevine foliar applications of phenylalanine and urea. Food Chem. 2015, 180, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Dhouibi, R.; Affes, H.; Ben Salem, M.; Hammami, S.; Sahnoun, Z.; Zeghal, K.M.; Ksouda, K. Screening of pharmacological uses of Urtica dioica and others benefits. Prog. Biophys. Mol. Biol. 2019, 150, 67–77. [Google Scholar] [CrossRef]
- Bisht, S.; Bhandari, S.; Bisht, N.S. Urtica dioica (L): An undervalued, economically important plant. Agric. Sci. Res. 2012, 2, 250–252. [Google Scholar]
- Hadizadeh, I.; Peivastegan, B.; Kolahi, M. Antifungal activity of nettle (Urtica dioica L.), Colocynth (Citrullus colocynthis L. Schrad), Oleander (Nerium oleander L.) and Konar (Ziziphus spina-christi L.) extracts on plants pathogenic fungi. Pak. J. Biol. Sci. 2009, 12, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Guil-Guerrero, J.L.; Rebolloso-Fuentes, M.M.; Torija Isasa, M.E. Fatty acids and carotenoids from stinging nettle (Urtica dioica L.). J. Food Compos. Anal. 2003, 16, 111–119. [Google Scholar] [CrossRef]
- Reeve, J.R.; Carpenter-Boggs, L.; Reganold, J.P.; York, A.L.; McGourty, G.; McCloskey, L.P. Soil and winegrape quality in biodynamically and organically managed vineyards. Am. J. Enol. Vitic. 2005, 56, 367–376. [Google Scholar]
- Villanueva-Rey, P.; Vázquez-Rowe, I.; Moreira, M.T.; Feijoo, G. Comparative life cycle assessment in the wine sector: Biodynamic vs. conventional viticulture activities in NW Spain. J. Clean. Prod. 2014, 65, 330–341. [Google Scholar] [CrossRef]
- Döring, J.; Frisch, M.; Tittmann, S.; Stoll, M.; Kauer, R. Growth, yield and fruit quality of grapevines under organic and biodynamic management. PLoS ONE 2015, 10, e0138445. [Google Scholar] [CrossRef] [Green Version]
- Meissner, G.; Athmann, M.; Fritz, J.; Kauer, R.; Stoll, M.; Schultz, H.R. Conversion to organic and biodynamic viticultural practices: Impact on soil, grapevine development and grape quality. OENO One 2019, 53, 639–659. [Google Scholar] [CrossRef]
- Tapwal, A.; Nisha; Garg, S.; Gautam, N.; Kumar, R. In vitro antifungal potency of plant extracts against five phytopathogens. Braz. Arch. Biol. Technol. 2011, 54, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Anderson, H. Invasive Japanese Knotweed (Fallopia japonica (Houtt.)) Best Management Practices in Ontario; Ontario Invasive Plant Council: Peterborough, ON, USA, 2012. [Google Scholar]
- Marchante, H.; Morais, M.; Freitas, H.; Marchante, E. Guia Prático Para a Identificação de Plantas Invasoras em Portugal; Imprensa da Universidade de Coimbra: Coimbra, Portugal, 2014; ISBN 9789892607856. [Google Scholar]
- Mahmoud Zaki, E.-R.; Eid, S.Y.; Al-Amodi, H.S.; Wink, M. Fallopia japonica: Bioactive secondary metabolites and molecular mode of anticancer. J. Tradit. Med. Clin. Naturop. 2016, 5, 1–20. [Google Scholar] [CrossRef]
- Fouillaud, M.; Caro, Y.; Venkatachalam, M.; Grondin, I.; Fouillaud, M.; Caro, Y.; Venkatachalam, M.; Grondin, I.; An, L.D.; Nollet, L.M.L.; et al. Anthraquinones; CRC Press: Boca Raton, FL, USA, 2017; ISBN 9781498722964. [Google Scholar]
- Oleszek, M.; Kowalska, I.; Oleszek, W. Phytochemicals in Bioenergy Crops; Springer: Berlin/Heidelberg, Germany, 2019; Volume 18. [Google Scholar]
- Barney, J.N.; Tharayil, N.; DiTommaso, A.; Bhowmik, P.C. The Biology of invasive alien plants in Canada. XX. Polygonum cuspidatum Sieb. & Zucc. [=Fallopia japonica (Houtt.) Dcne.]. Can. J. Plant Sci. 2006, 86, 887–905. [Google Scholar] [CrossRef] [Green Version]
- Patočka, J.; Navrátilová, Z.; Ovando, M. Biologically active compounds of Knotweed (Reynoutria spp.). Mil. Med. Sci. Lett. 2017, 86, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-C.; Lim, M.Y.; Lee, H.S. Emodin isolated from Cassia obtusifolia (Leguminosae) seed shows larvicidal activity against three mosquito species. J. Agric. Food Chem. 2003, 51, 7629–7631. [Google Scholar] [CrossRef]
- Werner, S.J.; Tupper, S.K.; Pettit, S.E.; Ellis, J.W.; Carlson, J.C.; Goldade, D.A.; Hofmann, N.M.; Jeffrey Homan, H.; Linz, G.M. Application Strategies for an anthraquinone-based repellent to protect oilseed sunflower crops from pest blackbirds. Crop Prot. 2014, 59, 63–70. [Google Scholar] [CrossRef]
- Verkleij, F.N. Seaweed extracts in agriculture and horticulture: A review. Biol. Agric. Hortic. 1992, 8, 309–324. [Google Scholar] [CrossRef]
- Zodape, S.T.; Gupta, A.; Bhandari, S.C.; Rawat, U.S.; Chaudhary, D.R.; Eswaran, K.; Chikara, J. Foliar application of seaweed sap as biostimulant for enhancement of yield and quality of tomato (Lycopersicon esculentum Mill.). J. Sci. Ind. Res. 2011, 70, 215–219. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Jayaraman, J.; Norrie, J.; Punja, Z. Commercial Extract From the brown seaweed Ascophyllum nodosum reduces fungal diseases in greenhouse cucumber. J. Appl. Phycol. 2011, 23, 353–361. [Google Scholar] [CrossRef]
- Rayirath, P.; Benkel, B.; Mark Hodges, D.; Allan-Wojtas, P.; MacKinnon, S.; Critchley, A.T.; Prithiviraj, B. Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing tolerance in Arabidopsis thaliana. Planta 2009, 230, 135–147. [Google Scholar] [CrossRef]
- Alam, M.Z.; Braun, G.; Norrie, J.; Hodges, D.M. Effect of Ascophyllum extract application on plant growth, fruit yield and soil microbial communities of strawberry. Can. J. Plant Sci. 2012, 93, 23–36. [Google Scholar] [CrossRef]
- Correia, S.; Oliveira, I.; Queirós, F.; Ribeiro, C.; Ferreira, L.; Luzio, A.; Silva, A.P.; Gonçalves, B. Preharvest Application of seaweed based biostimulant reduced cherry (Prunus avium L.) cracking. Procedia Environ. Sci. 2015, 29, 251–252. [Google Scholar] [CrossRef] [Green Version]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K. Seaweed extract reduces foliar fungal diseases on carrot. Crop Prot. 2008, 27, 1360–1366. [Google Scholar] [CrossRef]
- Correia, S.; Santos, M.; Glińska, S.; Gapińska, M.; Matos, M.; Carnide, V.; Schouten, R.; Silva, A.P.; Gonçalves, B. Effects of exogenous compound sprays on cherry cracking: Skin properties and gene expression. J. Sci. Food Agric. 2020, 100, 2911–2921. [Google Scholar] [CrossRef]
- Cabo, S.; Morais, M.C.; Aires, A.; Carvalho, R.; Pascual-Seva, N.; Silva, A.P.; Gonçalves, B. Kaolin and seaweed-based extracts can be used as middle and long-term strategy to mitigate negative effects of climate change in physiological performance of hazelnut tree. J. Agron. Crop Sci. 2019, 206, 28–42. [Google Scholar] [CrossRef]
- Cabo, S.; Aires, A.; Carvalho, R.; Vilela, A.; Pascual-Seva, N.; Silva, A.P.; Gonçalves, B. Kaolin, Ascophyllum nodosum and salicylic acid mitigate effects of summer stress improving hazelnut quality. J. Sci. Food Agric. 2020, 101, 459–475. [Google Scholar] [CrossRef]
- Demain, A.L.; Phaff, H.J.; Kurtzman, C.P. The industrial and agricultural significance of yeasts. In The Yeasts; Cletus, P., Kurtzman, J.W.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 13–19. ISBN 9780444813121. [Google Scholar]
- Mukherjee, A.; Verma, J.P.; Gaurav, A.K.; Chouhan, G.K.; Patel, J.S.; Hesham, A.E. Yeast a potential bio-agent: Future for plant growth and postharvest disease management for sustainable agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 1497–1510. [Google Scholar] [CrossRef]
- Kapteyn, J.C.; Van Den, E.H.; Klis, F.M. The contribution of cell wall proteins to the organization of the yeast cell wall. Biochim. Biophys. Acta 1999, 1426, 373–383. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Portu, J.; López, R.; Santamaría, P.; Garde-Cerdán, T. Elicitor and nitrogen applications to Garnacha, Graciano and Tempranillo vines: Effect on grape amino acid composition. J. Sci. Food Agric. 2018, 98, 2341–2349. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Portu, J.; Santamaría, P.; López, R.; Garde-Cerdán, T. Effects on grape amino acid concentration through foliar application of three different elicitors. Food Res. Int. 2017, 99, 688–692. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Agudelo-Romero, P.; Erban, A.; Rego, C.; Carbonell-Bejerano, P.; Nascimento, T.; Sousa, L.; Martínez-Zapater, J.M.; Kopka, J.; Fortes, A.M. Transcriptome and metabolome reprogramming in Vitis vinifera cv. Trincadeira berries upon infection with Botrytis cinerea. J. Exp. Bot. 2015, 66, 1769–1785. [Google Scholar] [CrossRef] [Green Version]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Meijón, M.; Ferreira, H.; Pinto, G.; Moutinho-Pereira, J.; Correia, C. Salicylic acid modulates olive tree physiological and growth responses to drought and rewatering events in a dose dependent manner. J. Plant Physiol. 2018, 230, 21–32. [Google Scholar] [CrossRef]
- Trevisan, F.; Lima, C.; Pinto, V.; Bonome, L.; de Liz, K. Ácido salicílico no desenvolvimento de plantas e nas características físico-químicas de frutas de morango “Milsei-Tudla”. Rev. Iberoam. Tecnol. Postcosecha 2017, 18, 106–114. [Google Scholar]
- Dutta, T.; Neelapu, N.R.R.; Wani, S.H.; Challa, S. Compatible solute engineering of crop plants for improved tolerance toward abiotic stresses. In Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Wani, S.H., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 221–254. ISBN 9780128130667. [Google Scholar]
- Adak, N. Effects of glycine betaine concentrations on the agronomic characteristics of strawberry grown under deficit irrigation conditions. Appl. Ecol. Environ. Res. 2019, 17, 3753–3767. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Chapman, P.; Collier-Christian, L. Endogenous levels and exogenous application of glycine betaine to grapevines. Sci. Hortic. 2006, 111, 7–16. [Google Scholar] [CrossRef]
- Hussain Wani, S.; Brajendra Singh, N.; Haribhushan, A.; Iqbal Mir, J. Compatible solute engineering in plants for abiotic stress tolerance—Role of glycine betaine. Curr. Genom. 2013, 14, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Hayes, M.A.; Shor, A.C.; Jesse, A.; Miller, C.; Kennedy, J.P.; Feller, I. The role of glycine betaine in range expansions; protecting mangroves against extreme freeze events. J. Ecol. 2020, 108, 61–69. [Google Scholar] [CrossRef]
- Awad, M.A.; Al-Qurashi, A.D.; Mohamed, S.A. Postharvest trans-resveratrol and glycine betaine treatments affect quality, antioxidant capacity, antioxidant compounds and enzymes activities of ‘El-Bayadi’ table grapes after storage and shelf life. Sci. Hortic. 2015, 197, 350–356. [Google Scholar] [CrossRef]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Singh, H.P.; Cha-um, S. Foliar application of glycine betaine regulates soluble sugars and modulates physiological adaptations in sweet potato (Ipomoea batatas) under water deficit. Protoplasma 2020, 257, 197–211. [Google Scholar] [CrossRef]
- Khadouri, H.K.; Kandhan, K.; Salem, M.A. Effects of glycine betaine on plant growth and performance of Medicago sativa and Vigna unguiculata under water deficit conditions. J. Phytol. 2020, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Correia, S.; Queirós, F.; Ribeiro, C.; Vilela, A.; Aires, A.; Barros, A.I.; Schouten, R.; Silva, A.P.; Gonçalves, B. Effects of calcium and growth regulators on sweet cherry (Prunus avium L.) quality and sensory attributes at harvest. Sci. Hortic. 2019, 248, 231–240. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of combined applications of trichoderma virens and a biopolymer-based biostimulant on lettuce agronomical, physiological, and qualitative properties under variable n regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Nardi, S.; Carletti, P.; Pizzeghello, D.; Muscolo, A. Biological activities of humic substances. In Biophysico-Chemical Processes Involving Natural Nonliving Organic Matter in Environmental Systems; Senesi, N., Xing, B., Huang, P., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 305–339. [Google Scholar] [CrossRef]
- Koyama, R.; Roberto, S.R.; de Souza, R.T.; Borges, W.F.S.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous abscisic acid promotes anthocyanin biosynthesis and increased expression of flavonoid synthesis genes in Vitis vinifera × Vitis labrusca table grapes in a subtropical region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef] [Green Version]
- Dong, T.; Zheng, T.; Fu, W.; Guan, L.; Jia, H.; Fang, J. The effect of ethylene on the color change and resistance to Botrytis cinerea infection in “Kyoho” grape fruits. Foods 2020, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- Legay, G.; Marouf, E.; Berger, D.; Neuhaus, J.M.; Mauch-Mani, B.; Slaughter, A. Identification of genes expressed during the compatible interaction of grapevine with Plasmopara viticola through suppression subtractive hybridization (SSH). Eur. J. Plant Pathol. 2011, 129, 281–301. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.A.; Kumari, P.H.; Kumar, G.S.; Mohanalatha, C.; Kishor, P.B.K. Osmotin: A plant sentinel and a possible agonist of mammalian adiponectin. Front. Plant Sci. 2015, 6, 163. [Google Scholar] [CrossRef] [Green Version]
- Dufour, M.C.; Magnin, N.; Dumas, B.; Vergnes, S.; Corio-Costet, M.F. High-throughput gene-expression quantification of grapevine defense responses in the field using microfluidic dynamic arrays. BMC Genomics 2016, 17, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesler, A.; Perazzolli, M.; Puopolo, G.; Giovannini, O.; Elad, Y.; Pertot, I. A complex protein derivative acts as biogenic elicitor of grapevine resistance against powdery mildew under field conditions. Front. Plant Sci. 2015, 6, 715. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Treatment | Matrix | Effect | References |
|---|---|---|---|
| Seaweed Extract | Grape/wine | A. nodosum extracts, improved quality, increased plant defenses by the regulation of molecular, physiological and biochemical processes. | [49,50,51] |
| Grape | Laminaria digitata extracts, reduced the infection caused by P. viticola and B. cinerea. Ulva armoricana extracts protect against E. necator. | [32,34,52] | |
| Chitosan | Grape | Stimulated defense responses against P. viticola and B. cinerea. Increased levels of polyphenols, anthocyanins, and tannins in cv. Tinto Cão berries, and polyphenols and tannins in cv. Touriga Franca. Accumulation of phenolics in berries, namely anthocyanins in cv. Tinto Cão. | [53,54,55,56,57] |
| Grapevine | Improved rooting of the cuttings, increased the number of new shoots, their length and number of internodes, as well as the chlorophyll content in the leaves. | [58] | |
| Grape/wine | The volatile profile (raising the levels of total acetals and alcohols) in cv. Groppello Gentile, responsible for the wine flavor and taste. No substantial effect on phenolic content including anthocyanins, stilbenes, and flavonols, on either grape or wine of cv. Tempranillo. | [17,59] | |
| Yeast Extract | Grape/wine | Saccharomyces cerevisiae extracts enhanced anthocyanin and stilbene contents in grape and wine of cv. Tempranillo. | [17] |
| Grape | A mixture of Laminaria digitata and Saccharomyces spp. Extracts used to control P. viticola in the cv. Montepulciano did not affect grape quality (amino acid content). | [34] | |
| Japanese Knotweed (anthraquinone) | Grape | Protect against P. viticola in cv. Chasselas. | [42] |
| Kaolin | Grape | Enhanced photosynthetic activity, improved antioxidant capacity, increased concentration of phenolics including anthocyanins, vitamin C and sugars, in addition to protecting the leaves and berries of cv. Touriga Nacional from solar radiation. Protected the berry exocarp from light stress in cv. Alvarinho. | [2,10,47,48] |
| Glycine Betaine | Grapevine | Increased the resistance to water stress. Significantly affected leaf relative water content (RWC), canopy temperature, chlorophyll content, soluble carbohydrate contents, and leaf area. Under drought stress, seemed to influence the leaf water status, slightly alleviating the leaf water loss and increasing leaves fresh weight in cv. Alphonse Lavallée. | [60,61] |
| Methyl Jasmonate | Grapevine | Enhanced tolerance of grapevine foliar cuttings and vineyard against the pathogen E. necator in cv. Cabernet Sauvignon. | [33] |
| Grape/wine | Increased the phenolic compounds content, depending on the variety and vintage. | [17,62,63,64] | |
| Abscisic Acid | Grape | Increased °Brix, total phenolic and anthocyanin contents and improved the production of volatile aromas in cv. Cabernet Sauvignon berries. | [65] |
| Salicylic Acid | Grape | Increased total phenolic content and antioxidant activity in cv. Syrah. Delayed berries ripening in the cvs. Bez El Naka and Shiraz | [66,67,68] |
| Others | Grape | Urea: Increased yeast assimilable nitrogen, berry amino acid, flavonoid, stilbene and anthocyanin concentrations. Foliar N and foliar N + S: Improved vine nitrogen status and enhanced aroma expression in Sauvignon blanc wines without the negative impact on vigour and Botrytis susceptibility. Phenylalanine: Small improvement in some phenolic compounds. | [69,70,71] |
| Protein (Gene) | Function | References |
|---|---|---|
| Amine oxidase (AO) | ROS pathway gene. | [56] |
| Anthocyanin transporters (ABCC1) | Anthocyanin transporter gene. | [57] |
| Anthocyanidin reductase (ANR) | Gene of the synthesis pathway of proanthocyanidins. | [57] |
| Callose synthase 2 (VvCaS2) | Involved in the synthesis of the β-1,3-glucan callose, a plant defense response common to different resistance pathways. | [51] |
| Catalase (CAT) | ROS pathway gene. | [56] |
| Cu/Zn-superoxide dismutase (Cu/Zn-SOD) | ROS pathway gene. | [56] |
| Dihydroflavonol reductase (DFR) | Key gene in the synthesis pathway of anthocyanins. | [51,122] |
| Fe superoxide dismutase (Fe-SOD) | ROS pathway gene. | [56] |
| Glutathione reductase (GR) | ROS pathway gene. | [56] |
| Glutathion-S-transferase (GST) | Essencial in the detoxification of elicitor-generated oxidants. | [32] |
| β-1,3-Glucanase (Glu1) | Involved in plant defense. Gene encoding a basic β-1,3-glucanase thought to hydrolyse the structural β-1,3-glucan present in some fungal cell walls. | [32,33] |
| Leucoanthocyanin Dioxygenase (LDOX) | Key gene in the synthesis pathway of polyphenols, including anthocyanins. | [2] |
| 9-Lipoxygenase (LOX) | Essential for the resistance to fungal infection. | [32] |
| MATE1 | Anthocyanin transporter gene. | [57] |
| Myeloblastosis A1 and A2 (MYBA1 and MYBA2) | Key transcription factors of the synthesis pathway of polyphenols, including anthocyanins. | [106,107,123] |
| Phenylalanine Ammonia Lyase (PAL) | Catalyses the first step in the phenylpropanoid pathway. | [2,17,32,106] |
| Pathogenesis-Related (PR) proteins | Responsible for plant defense by hydrolysing fungal cell wall components. PR1 is usually reported to be a typical marker of SAR. PR3/PR4/PR8/PR11 are chitinase proteins, that are strongly induced in the host plant, after the pathogen infection. PR5 is a thaumatin-like/osmotin, and is a responsive antifungal protein that confers tolerance to both biotic and abiotic stresses in plants. PR10 may play an important role in the defense of grapevine against P. viticola. PR17 is related to defense responses. An accumulation of PR17 after P. viticola inoculation has been verified. PIN codifies for a serine-proteinase inhibitor (PIN), a class of antifungal PR-6 proteins, which have potent activity against pathogens. | [32,124,125,126,127] |
| Polygalacturonase-inhibiting protein (PGIP) | The PGIP gene product, a polygalacturase-inhibiting protein, interacts with extracellular endo-α-1,4-polygalacturonases (PGs), secreted by phytopathogenic fungi, to inhibit their activity. | [32] |
| Resveratrol Synthase (STS1) | Involved in resveratrol and ε-viniferin production, the two major phytoalexinsin the protection against pathogens. | [32] |
| UDP-glucose:flavonoid 3-O-glucosyltransferase (UFGT) | Key gene of the flavonoid biosynthetic pathway. | [2,106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, E.; Gonçalves, B.; Cortez, I.; Castro, I. The Role of Biostimulants as Alleviators of Biotic and Abiotic Stresses in Grapevine: A Review. Plants 2022, 11, 396. https://doi.org/10.3390/plants11030396
Monteiro E, Gonçalves B, Cortez I, Castro I. The Role of Biostimulants as Alleviators of Biotic and Abiotic Stresses in Grapevine: A Review. Plants. 2022; 11(3):396. https://doi.org/10.3390/plants11030396
Chicago/Turabian StyleMonteiro, Eliana, Berta Gonçalves, Isabel Cortez, and Isaura Castro. 2022. "The Role of Biostimulants as Alleviators of Biotic and Abiotic Stresses in Grapevine: A Review" Plants 11, no. 3: 396. https://doi.org/10.3390/plants11030396
APA StyleMonteiro, E., Gonçalves, B., Cortez, I., & Castro, I. (2022). The Role of Biostimulants as Alleviators of Biotic and Abiotic Stresses in Grapevine: A Review. Plants, 11(3), 396. https://doi.org/10.3390/plants11030396