
Degradation of Carbendazim by Molecular Hydrogen on Leaf Models



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
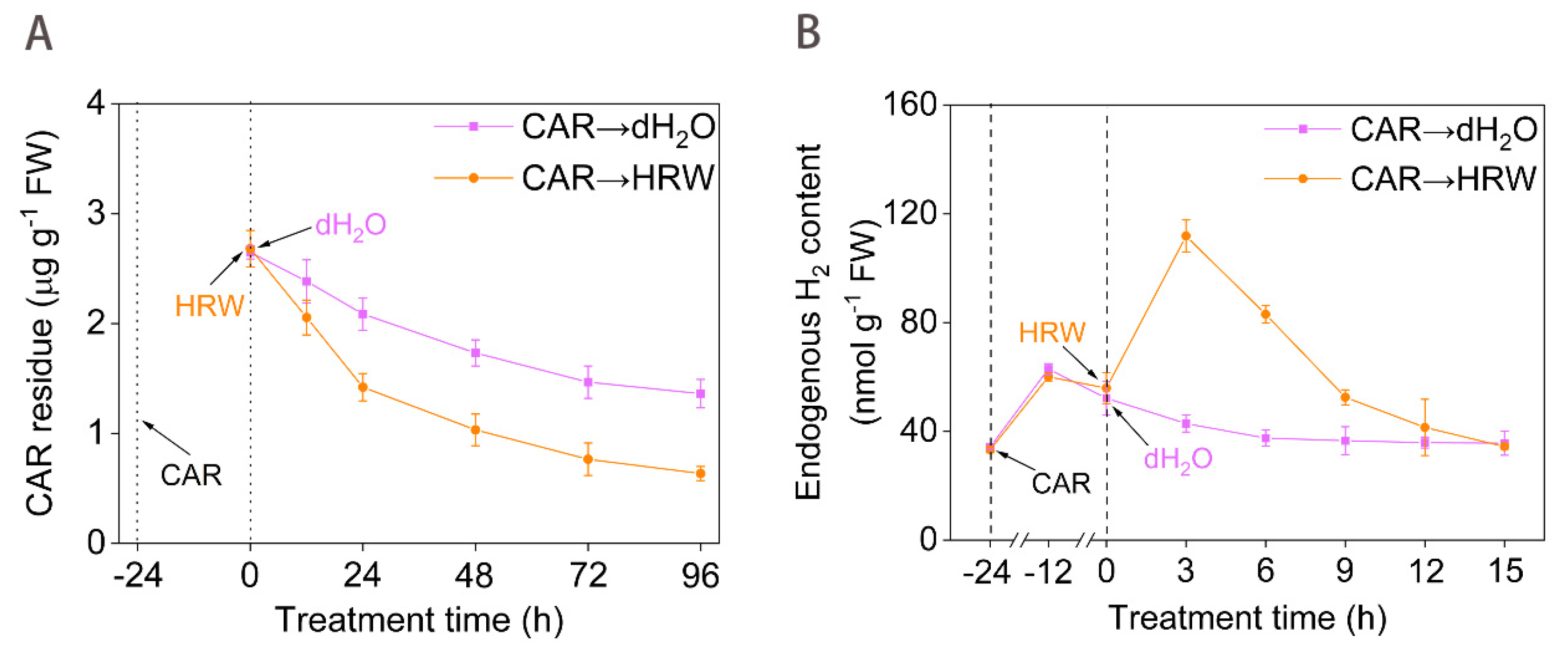
2.1. Exogenous H2 Control of CAR Degradation
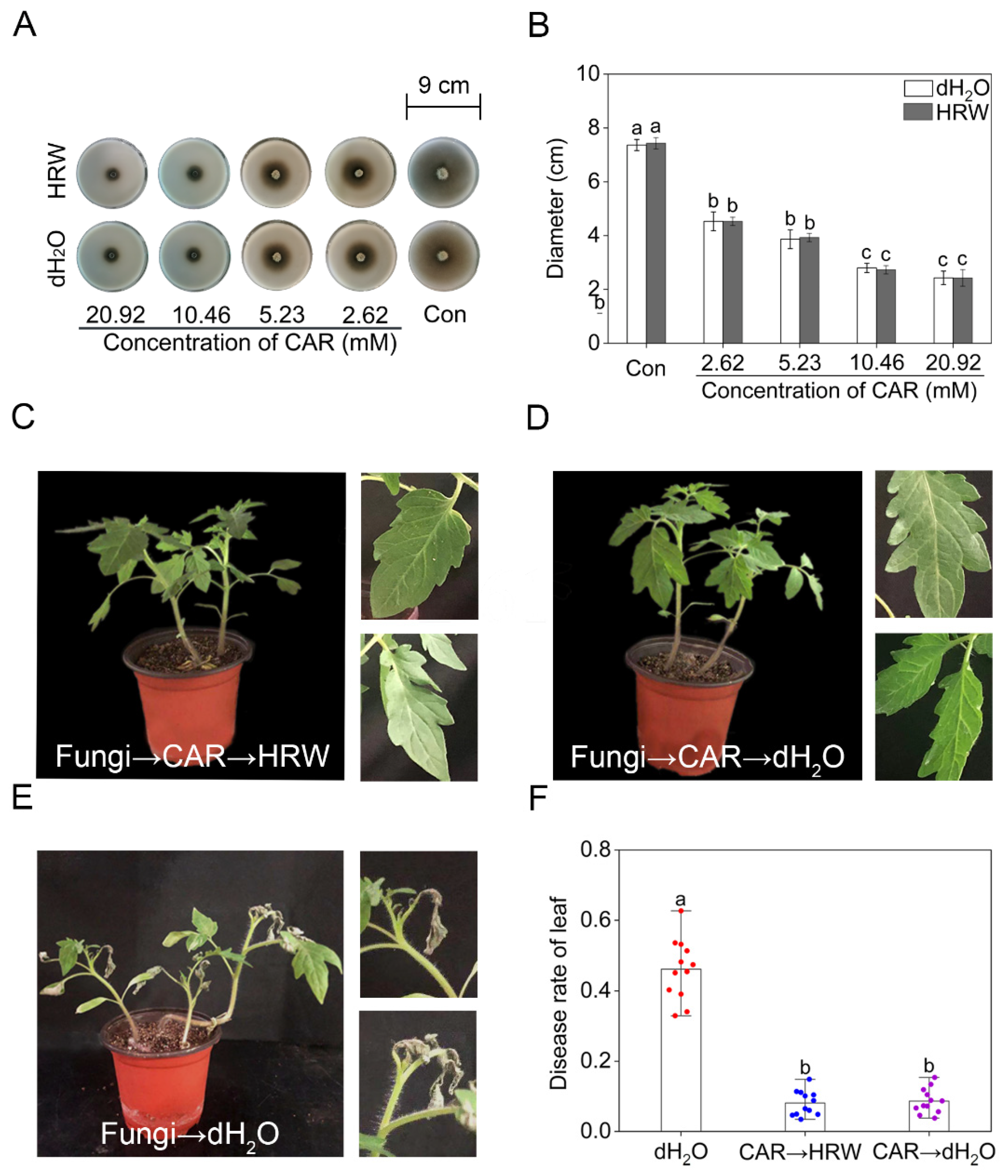
2.2. Exogenous H2 Did Not Alter the Antifungal Effect of CAR
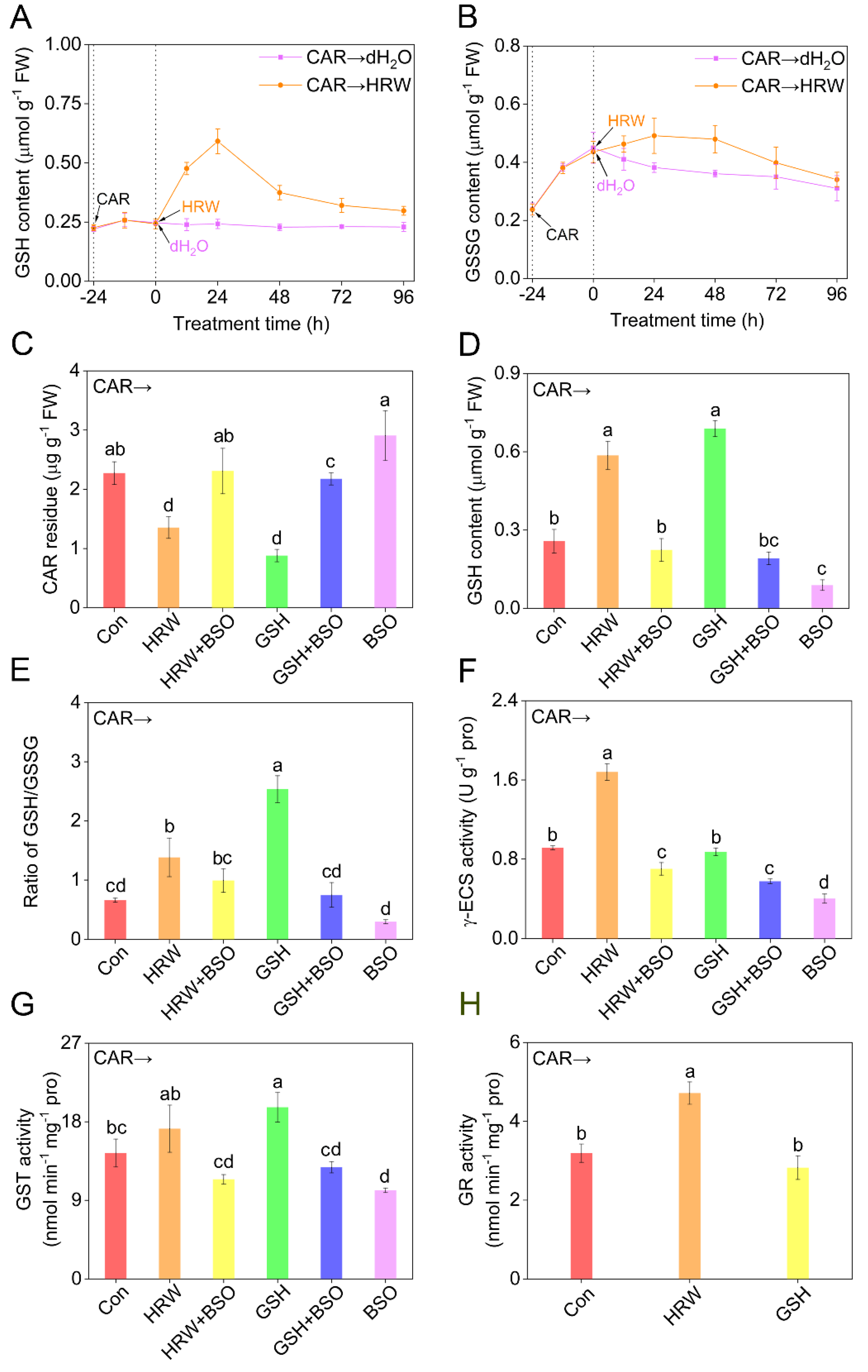
2.3. Glutathione Involvement in Molecular Hydrogen Control of CAR Degradation
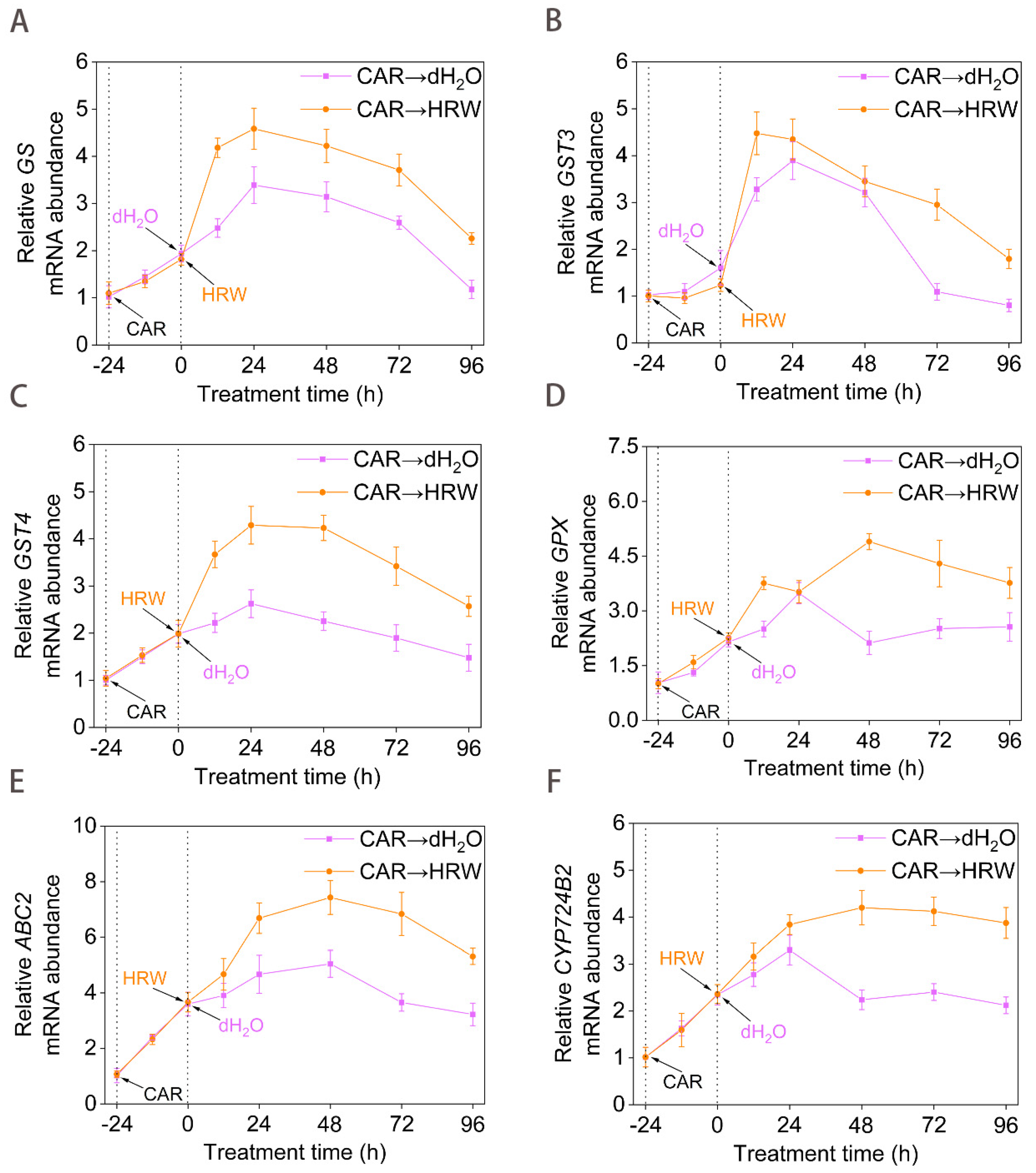
2.4. Transcriptional Regulation of GSH-Metabolism and Detoxification Genes
2.5. CAR-Triggered Redox Imbalance Was Abolished by HRW
2.6. Genetic Evidence Revealed That Endogenous Molecular Hydrogen Can Positively Influence Carbendazim Degradation via GSH
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material, Growth Conditions, and Treatments
3.3. Preparation of Hydrogen-Rich Water
3.4. Determination of H2 Content
3.5. Determination of GSH Content by UPLC Analysis
3.6. Determination of CAR Residues in Tomato Leaves
3.7. Indoor Toxicity Assessment and Experiment of Early Blight Resistance
3.8. Analysis of the GSH Cycle
3.9. RT-qPCR Determination of Transcript Levels of Genes
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Lee, S.J.; Mehler, L.; Beckman, J.; Diebolt-Brown, B.; Prado, J.; Lackovic, M.; Waltz, J.; Mulay, P.; Schwartz, A.; Mitchell, Y.; et al. Acute pesticide illnesses associated with off-target pesticide drift from agricultural applications: 11 states, 1998–2006. Environ. Health Perspect. 2011, 119, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA). Conclusion on the peer review of the pesticide risk assessment of the active substance carbendazim. EFSA J. 2010, 8, 1598. [Google Scholar] [CrossRef] [Green Version]
- Davidse, L.C. Benzimidazole fungicides: Mechanism of action and biological impact. Annu. Rev. Phytopathol. 1986, 24, 43–65. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, Y.; Harvey, P.R.; Li, H.; Ren, Y.; Li, J.; Wang, J.; Yang, H. Isolation and characterization of carbendazim-degrading Rhodococcus erythropolis djl-11. PLoS ONE 2013, 8, e74810. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, N.; Kumar, V.; Datta, S.; Wani, A.B.; Singh, D.; Singh, K.; Singh, J. Toxicity, monitoring and biodegradation of the fungicide carbendazim. Environ. Chem. Lett. 2016, 14, 317–329. [Google Scholar] [CrossRef]
- Besson-Barda, A.; Courtoisa, C.; Gauthiera, A.; Dahana, J.; Dobrowolskab, G.; Jeandrozc, S.; Pugin, A.; Wendehenne, D. Nitric oxide in plants: Production and cross-talk with Ca2+ signaling. Mol. Plant 2008, 1, 218–228. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Zhou, Y.; Zhou, Y.; Shi, K.; Zhou, J.; Yu, Y.; Yu, J.; Xia, X. Interplay between mitogen-activated protein kinase and nitric oxide in brassinosteroid-induced pesticide metabolism in Solanum lycopersicum. J. Hazard. Mater. 2016, 316, 221–231. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Ohta, S. Molecular hydrogen as a preventive and therapeutic medical gas: Initiation, development and potential of hydrogen medicine. Pharmacol. Ther. 2014, 144, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Sun, X.J. Hydrogen biology: It is just beginning. Chin. J. Biochem. Mol. Biol. 2019, 35, 1037–1050. [Google Scholar] [CrossRef]
- Gaffron, H. Reduction of carbon dioxide with molecular hydrogen in green algae. Nature 1939, 143, 204–205. [Google Scholar] [CrossRef]
- Renwick, G.M.; Giumarro, C.; Siegel, S.M. Hydrogen metabolism in higher plants. Plant Physiol. 1964, 39, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.; Layzell, D.B. Gas-exchange of legume nodules and the regulation of nitrogenase activity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 483–511. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Liu, Y.H.; Wang, S.; Du, H.M.; Shen, W. Hydrogen agronomy: Research progress and prospects. J. Zhejiang Univ. Sci. B 2020, 21, 841–855. [Google Scholar] [CrossRef]
- Li, L.; Lou, W.; Kong, L.; Shen, W. Hydrogen commonly applicable from medicine to agriculture: From molecular mechanisms to the field. Curr. Pharm. Des. 2021, 27, 747–759. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, T.; Wang, J.; Xu, S.; Shen, W. Regulation of chlorothalonil degradation by molecular hydrogen. J. Hazard. Mater. 2022, 424, 127291. [Google Scholar] [CrossRef]
- Wang, Z.W.; Huang, J.; Chen, J.Y.; Li, F.L. Time-dependent movement and distribution of chlorothalonil and chlorpyrifos in tomatoes. Ecotoxicol. Environ. Saf. 2013, 93, 107–111. [Google Scholar] [CrossRef]
- Su, J.; Yang, X.; Shao, Y.; Chen, Z.; Shen, W. Molecular hydrogen–induced salinity tolerance requires melatonin signaling in Arabidopsis thaliana. Plant Cell Environ. 2021, 44, 476–490. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, P.; Wang, Y.; Li, Y.; Su, J.; Chen, Z.; Yu, X.; Shen, W. Genetic elucidation of hydrogen signaling in plant osmotic tolerance and stomatal closure via hydrogen sulfide. Free Radic. Biol. Med. 2020, 161, 1–14. [Google Scholar] [CrossRef]
- Su, J.; Zhang, Y.; Nie, Y.; Cheng, D.; Wang, R.; Hu, H.; Chen, J.; Zhang, J.; Du, Y.; Shen, W. Hydrogen-induced osmotic tolerance isassociated with nitric oxide-mediated proline accumulation and reestablishment of redox balance in alfalfa seedlings. Environ. Exp. Bot. 2018, 147, 249–260. [Google Scholar] [CrossRef]
- Xie, Y.; Mao, Y.; Zhang, W.; Lai, D.; Wang, Q.; Shen, W. Reactive oxygen species-dependent nitric oxide production contributes to hydrogen-promoted stomatal closure in Arabidopsis. Plant Physiol. 2014, 165, 759–773. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Duan, X.; Yao, P.; Cui, W.; Cheng, D.; Zhang, J.; Jin, Q.; Chen, J.; Dai, T.; Shen, W. Hydrogen gas is involved in auxin-induced lateral root formation by modulating nitric oxide synthesis. Int. J. Mol. Sci. 2017, 18, 2084. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lou, W.; Wang, J.; Li, Q.; Shen, W. Glutathione produced by γ-glutamyl cysteine synthetase acts downstream of hydrogen to positively influence lateral root branching. Plant Physiol. Biochem. 2021, 167, 68–76. [Google Scholar] [CrossRef] [PubMed]
- De Wilde, T.; Spanoghe, P.; Ryckeboer, J.; Jaeken, P.; Springael, D. Sorption characteristics of pesticides on matrix substrates used in biopurification systems. Chemosphere 2009, 75, 100–108. [Google Scholar] [CrossRef]
- Yan, Y.; Sun, S.; Zhao, N.; Yang, W.; Shi, Q.; Gong, B. COMT1 overexpression resulting in increased melatonin biosynthesis contributes to the alleviation of carbendazim phytotoxicity and residues in tomato plants. Environ. Pollut. 2019, 252, 51–61. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, Y.; Cui, W.; Jin, Q.; Zhang, Y.; Bu, D.; Fu, J.; Wang, R.; Zhou, F.; Shen, W. Hydrogen enhances adaptation of rice seedlings to cold stress via the reestablishment of redox homeostasis mediated by miRNA expression. Plant Soil 2017, 414, 53–67. [Google Scholar] [CrossRef]
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of heme oxygenase-1 signalling system. Plant Cell Environ. 2013, 36, 956–969. [Google Scholar] [CrossRef]
- Zulfqar, F.; Russell, G.; Hancock, J.T. Molecular hydrogen in agriculture. Planta 2021, 254, 56. [Google Scholar] [CrossRef]
- Xu, X.; Chen, J.; Li, B.; Tang, L. Carbendazim residues in vegetables in China between 2014 and 2016 and a chronic carbendazim exposure risk assessment. Food Control 2018, 91, 20–25. [Google Scholar] [CrossRef]
- Chaerani, R.; Voorrips, R.E. Tomato early blight (Alternaria solani): The pathogen, genetics, and breeding for resistance. J. Gen. Plant Pathol. 2006, 72, 335–347. [Google Scholar] [CrossRef]
- Coleman, J.O.D.; Randall, R.; Blake-Kalff, M.M.A. Detoxification of xenobiotics in plant cells by glutathione conjugation and vacuolar compartmenttaiization: A fluorescent assay using monochiorobimane. Plant Cell Environ. 1997, 20, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.B.; Zhang, Y.; Ahammeda, G.J.; Xia, X.J.; Mao, W.H.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Glutathione biosynthesis and regeneration play an important role in the metabolism of chlorothalonil in tomato. Chemosphere 2013, 90, 2563–2570. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, Y.; Chen, S.; Xia, X.; Shi, K.; Zhou, Y.; Yu, Y.; Yu, J. The different responses of glutathione-dependent detoxification pathway to fungicide chlorothalonil and carbendazim in tomato leaves. Chemosphere 2010, 79, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, J.; Soldner, F.; Schulz, J.B.; Klockgether, T.; Kovar, K.A.; Wüllner, U. Differential effects of L-buthionine sulfoximine and ethacrynic acid on glutathione levels and mitochondrial function in PC12 cells. Neurosci. Lett. 1999, 264, 1–4. [Google Scholar] [CrossRef]
- Vernoux, T.; Wilson, R.C.; Seeley, K.A.; Reichheld, J.P.; Muroy, S.; Brown, S.; Maughan, S.C.; Cobbett, C.S.; Montagu, M.V.; Inzé, D.; et al. The ROOT MERISTEMLESS1/CADMIUM SENSITIVE2 Gene defines a glutathione-dependent pathway involved in initiation and maintenance of cell division during postembryonic root development. Plant Cell 2000, 12, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Gao, C.; Fang, P.; Lin, G.; Shen, W. Alleviation of cadmium toxicity in Medicago sativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Marrs, K.A. The functions and regulation of glutathione S-transferases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, Y.; Wu, J.; Wang, J.; Zhou, Y.; Shi, K.; Yu, Y.; Yu, J. Brassinosteroids promote metabolism of pesticides in cucumber. J. Agric. Food Chem. 2009, 57, 8406–8413. [Google Scholar] [CrossRef]
- Yu, G.; Wei, J.; Chen, X.; Li, X.; Liu, X.; Ye, X.; Zhang, N.; Sun, W. The effect of glutathione in the regulation of the degradation of residual fungicide in tomato. Int. J. Agric. Biol. 2018, 20, 1873–1879. [Google Scholar] [CrossRef]
- Cobbett, C.S.; May, M.J.; Howden, R.; Rolls, B. The glutathione-deficient, cadmium-sensitive mutant, cad2–1, of Arabidopsis thaliana is deficient in γ-glutamylcysteine synthetase. Plant J. 1998, 16, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Religia, P.; Nguyen, N.D.; Nong, Q.D.; Matsuura, T.; Kato, Y.; Watanabe, H. Mutation of the cytochrome P450 CYP360A8 gene increases sensitivity to paraquat in Daphnia magn. Environ. Toxicol. Chem. 2021, 40, 1279–1288. [Google Scholar] [CrossRef]
- Deen, M.; Vries, E.G.; Timens, W.; Scheper, R.J.; Timmer-Bosscha, H.; Postma, D.S. ATP-binding cassette (ABC) transporters in normal and pathological lung. Resp. Res. 2005, 6, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, R.A. Plant ATP-binding cassette transporters. Annu. Rev. Plant Biol. 2007, 58, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Ruan, Y.P.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Brassinosteroid alleviates polychlorinated biphenyls-induced oxidative stress by enhancing antioxidant enzymes activity in tomato. Chemosphere 2013, 90, 2645–2653. [Google Scholar] [CrossRef]
- Mei, Y.; Chen, H.; Shen, W.; Huang, L. Hydrogen peroxide is involved in hydrogen sulfide-induced lateral root formation in tomato seedlings. BMC Plant Biol. 2017, 17, 162. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, P.; Kong, L.; Shen, W.; He, Q. Nanomaterial-mediated sustainable hydrogen supply induces lateral root formation via nitrate reductase-dependent nitric oxide. Chem. Eng. J. 2021, 405, 126905. [Google Scholar] [CrossRef]
- Herschbach, C.; Pilch, B.; Tausz, M.; Rennenberg, H.; Grill, D. Metabolism of reduced and inorganic sulphur in pea cotyledons and distribution into developing seedlings. New Phytol. 2002, 153, 73–80. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, M.; Guo, D.; Zhai, Y.; Miao, D.; Yue, J.; Yuan, C.; Zhao, M.; An, D. Antifungal effect of magnolol and honokiol from Magnolia officinalis on Alternaria alternata causing tobacco brown spot. Molecules 2019, 24, 2140. [Google Scholar] [CrossRef] [Green Version]
- Chaerani, R.; Groenwold, R.; Stam, P.; Voorrips, R.E. Assessment of early blight (Alternaria solani) resistance in tomato using a droplet inoculation method. J. Gen. Plant Pathol. 2007, 73, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Xue, Q.; Chen, Y.; Li, S.; Chen, L.; Ding, G.; Guo, D.; Guo, J. Evaluation of the strains of Acinetobacter and Enterobacter as potential biocontrol agents against Ralstonia wilt of tomato. Biol. Control 2009, 48, 252–258. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Li, X.; Shen, Z. Adaptive copper tolerance in Elsholtzia haichowensis involves production of Cu-induced thiol peptides. Plant Growth Regul. 2005, 47, 65–73. [Google Scholar] [CrossRef]
- Gong, B.; Li, X.; Bloszies, S.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Sodic alkaline stress mitigation by interaction of nitric oxide and polyamines involves antioxidants and physiological strategies in Solanum lycopersicum. Free Radic. Biol. Med. 2014, 71, 36–48. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Su, J.; Nie, Y.; Zhao, G.; Cheng, D.; Wang, R.; Chen, J.; Zhang, S.; Shen, W. Endogenous hydrogen gas delays petal senescence and extends the vase life of lisianthus cut flowers. Postharvest Biol. Technol. 2019, 147, 148–155. [Google Scholar] [CrossRef]
- Gong, B.; Wen, D.; VandenLangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves. Sci. Hortic. 2013, 157, 1–12. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Wang, Y.; Zhao, Z.; Xu, S.; Shen, W. Degradation of Carbendazim by Molecular Hydrogen on Leaf Models. Plants 2022, 11, 621. https://doi.org/10.3390/plants11050621
Zhang T, Wang Y, Zhao Z, Xu S, Shen W. Degradation of Carbendazim by Molecular Hydrogen on Leaf Models. Plants. 2022; 11(5):621. https://doi.org/10.3390/plants11050621
Chicago/Turabian StyleZhang, Tong, Yueqiao Wang, Zhushan Zhao, Sheng Xu, and Wenbiao Shen. 2022. "Degradation of Carbendazim by Molecular Hydrogen on Leaf Models" Plants 11, no. 5: 621. https://doi.org/10.3390/plants11050621
APA StyleZhang, T., Wang, Y., Zhao, Z., Xu, S., & Shen, W. (2022). Degradation of Carbendazim by Molecular Hydrogen on Leaf Models. Plants, 11(5), 621. https://doi.org/10.3390/plants11050621