
Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate


Abstract
:1. Introduction
2. Origin and Distribution
3. Genetic Diversity and Morphological Variation
4. C. sativa Yield Potential under Hot and Dry Conditions

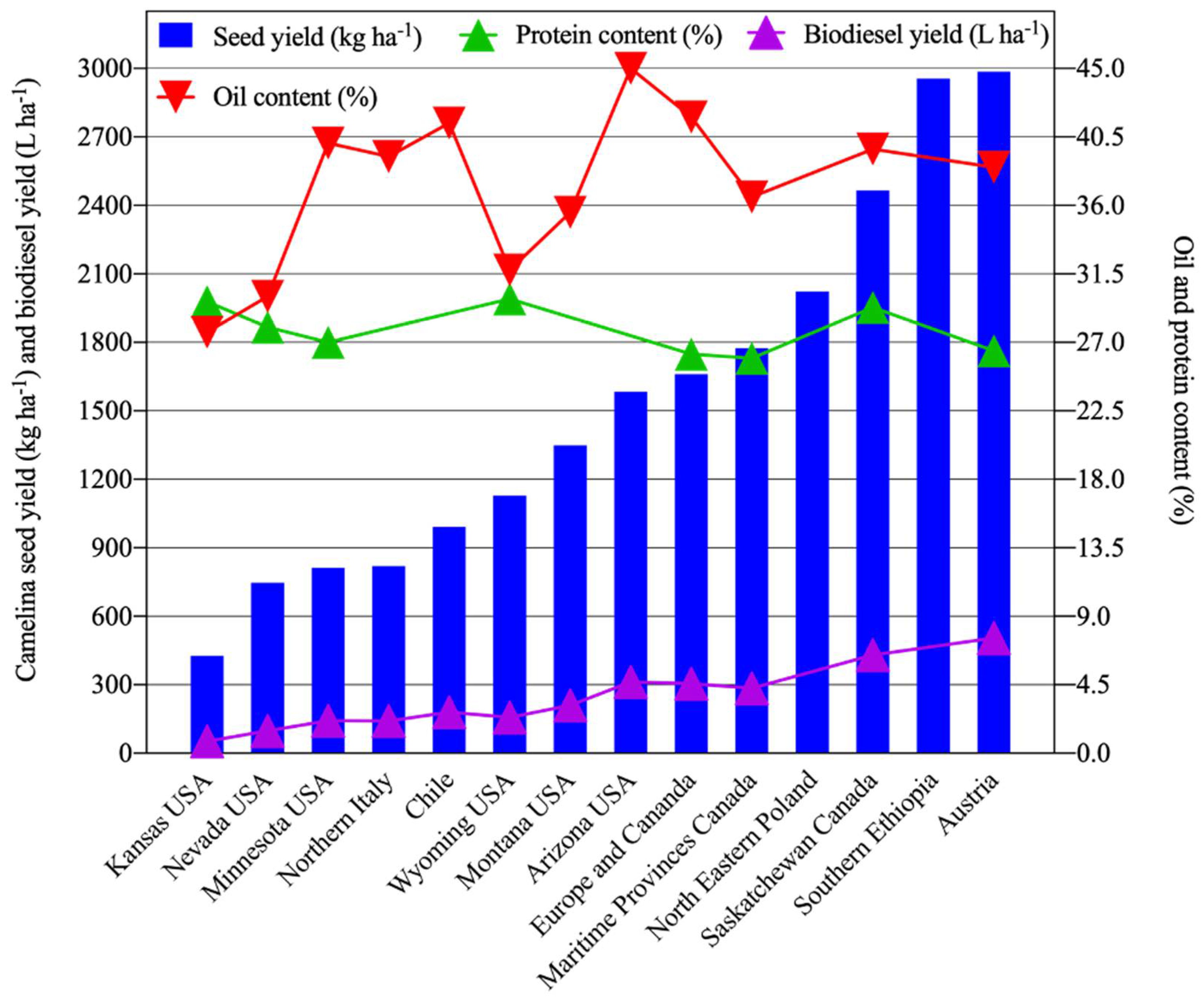{kind=link}
{kind=link}
| Locations | Seed Yield (kg ha−1) | Seed Oil Content (%) 1 | Seed Meal Protein Content (%) 2 | Biodiesel Yield (L ha−1) 3 | Major Sources of Variation | Reference | |
|---|---|---|---|---|---|---|---|
| Mean | Range | Range | Range | Mean | |||
| Austria | 2986 | 2419–3625 | 37.0–40.0 | 25.0–27.9 | 505 | Nitrogen and sulfur rates | [73] |
| Southern Ethiopia | 2956 | 2795–3200 | - | - | - | Seeding rates and nitrogen fertilizer | [74] |
| Saskatchewan, Canada | 2466 | 2184–2747 | 38 to 43 (39.7) | 27 to 32 (29.3) | 430 | Genotypes and environment | [39] |
| North–Eastern Poland | 2023 | 1700–2210 | - | - | - | Genotypes | [72] |
| Maritime Provinces of Eastern Canada | 1775 | 1638–1911 | 33.8–39.0 (36.6) | 25.0–26.8 (26.0) | 285 | Breeding lines, nitrogen and sulfur rates | [87] |
| Europe and Canada | 1660 | 1100–2700 | (41.8) | (26.2) | 305 | Genotypes and environment | [77] |
| Arizona, USA | 1583 | 1527–1638 | (45) | - | 313 | Nitrogen rates, water use and irrigation scheduling | [84] |
| Montana USA | 1349 | 546–2942 | 33.5–37.6 (36.0) | - | 211 | Cultivars and locations | [78] |
| Wyoming, USA | 1129 | 832–1643 | 31.1–32.4 | 29.3–30.4 | 157 | Nitrogen and sulfur rates | [91] |
| Chile | 991 | 387–2314 | 39.8–45.7 (41.41) | - | 180 | Cultivars, planting dates, and locations | [61] |
| Northern Italy | 820 | 600–940 | (39.2) | 141 | Low input and growing seasons | [71] | |
| Nevada, USA | 899 | 770–1013 | 31.8–33.3 (32.4) | 27.1–28.1 (27.5) | 128 | Cultivars and irrigation rates | [36] |
| 784 | 534–1010 | 31.8–32.6 (32.2) | 26.2–30.4 (28.4) | 111 | Cultivars, nitrogen sources and rates | [59] | |
| 735 | 34–1921 | 26.6–30.8 | - | 69.7 | Cultivars and year | [37] | |
| 570 | 130–921 | 28.4–29.5 | - | 72 | cultivars, sowing date, and methods | [62] | |
| Minnesota, USA | 812 | 650–944 | 39.4–40.7 (40.1) | 26.2–27.9 (27.0) | 143 | Genotypes and seeding rate | [3] |
| Kansas, USA | 427 | 317–503 | 27–29 (27.7) | 29–30 (29.7) | 52 | Cultivar and planting date | [75] |
| Overall mean | 1410 | 36.0 | 27.8 | 208.4 | |||
5. Biotic Production Constraints of C. sativa
5.1. Weed Control
5.2. Insect Pests
5.3. Fungal, Bacterial, and Viral Pathogens
6. Ecosystem Services Provided by C. sativa
7. Fatty Acid Synthesis and Seed Oil Profiling in C. sativa
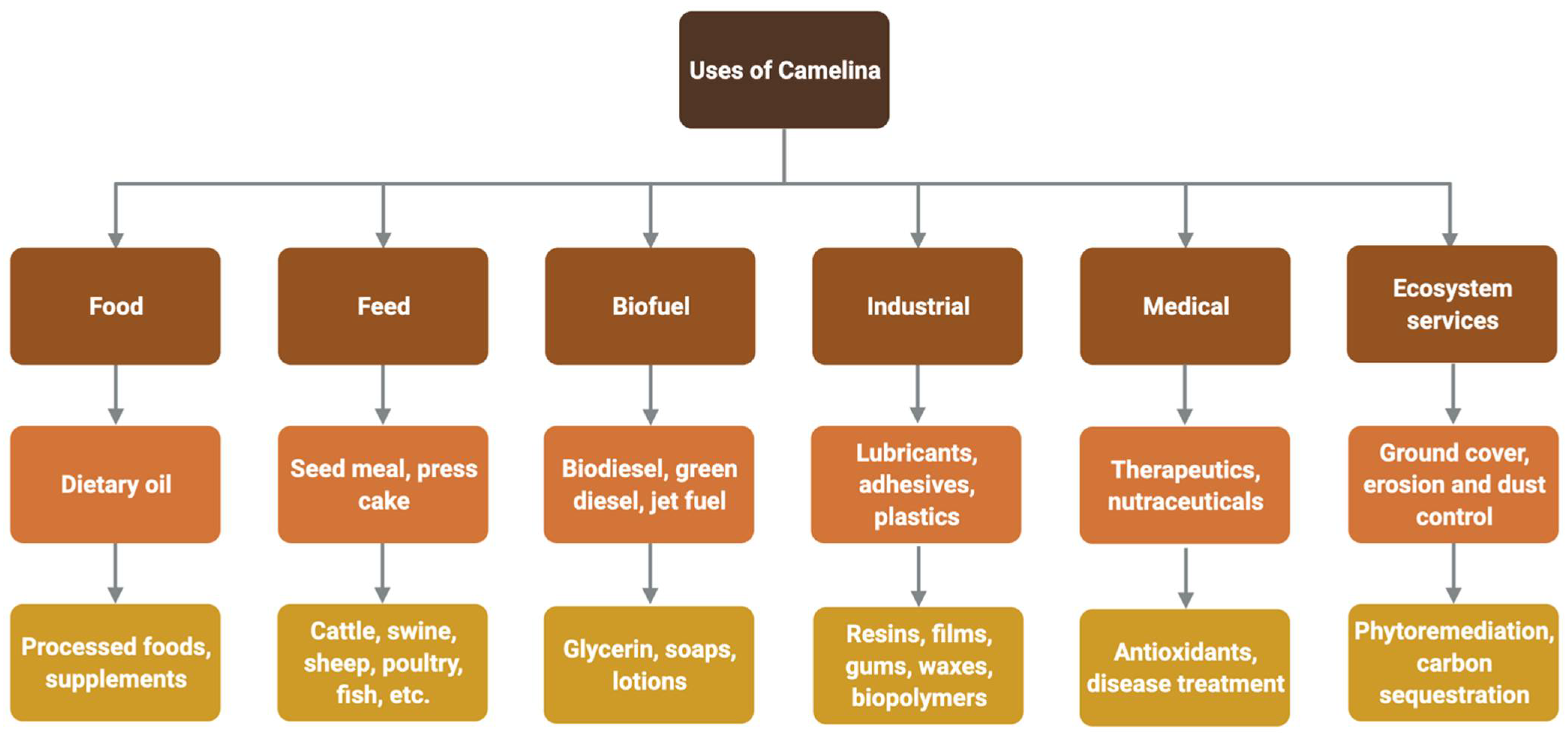
8. Uses of C. sativa
8.1. Food and Food Products for Humans
8.2. Feed for Animal Nutrition
8.2.1. Beef Cattle
8.2.2. Dairy Cows
8.2.3. Sheep and Goat
8.2.4. Swine
8.2.5. Poultry
8.2.6. Fish
8.3. Biodiesel/Renewable Diesel Fuel
8.4. Hydro-Processed Renewable Jet (HRJ) Fuel
8.5. Fast Pyrolysis Bio-Oil
8.6. Industrial Applications
8.7. Biomedical Uses
9. Strategic Innovations for Climate-Resilient C. sativa
9.1. Molecular Breeding Approaches
9.2. Improving Productivity Using Rhizosphere Microbes
9.3. Improving Quality Traits
9.4. Improving Stress Tolerance Traits
9.5. Genome-Editing Approaches
10. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bansal, S.; Durrett, T.P. Camelina sativa: An ideal platform for the metabolic engineering and field production of industrial lipids. Biochimie 2016, 120, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crops Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Gesch, R.W.; Matthees, H.L.; Alvarez, A.L.; Gardner, R.D. Winter Camelina: Crop growth, seed yield, and quality response to cultivar and seeding rate. Crop Sci. 2018, 58, 2089–2098. [Google Scholar] [CrossRef]
- Putnam, D.H.; Budin, J.T.; Field, L.A.; Breene, W.M. Camelina: A promising low-input oilseed. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 314–322. [Google Scholar]
- Schillinger, W.F. Camelina: Long-term cropping systems research in a dry Mediterranean climate. Field Crops Res. 2019, 235, 87–94. [Google Scholar] [CrossRef]
- Vollmann, J.; Eynck, C. Camelina as a sustainable oilseed crop: Contributions of plant breeding and genetic engineering. Biotechnol. J. 2015, 10, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Masella, P.; Martinelli, T.; Galasso, I. Agronomic evaluation and phenotypic plasticity of Camelina sativa growing in Lombardia, Italy. Crop Pasture Sci. 2014, 65, 453–460. [Google Scholar] [CrossRef]
- Iskandarov, U.; Kim, H.J.; Cahoon, E.B. Camelina: An emerging oilseed platform for advanced biofuels and bio-based materials. In Plants and Bioenergy; Advances in Plant Biology; McCann, M., Buckeridge, M., Carpita, N., Eds.; Springer: New York, NY, USA, 2014; Volume 4, pp. 131–140. [Google Scholar]
- Liu, X.; Brost, J.; Hutcheon, C.; Guilfoil, R.; Wilson, A.K.; Leung, S.; Shewmaker, C.K.; Rooke, S.; Nguyen, T.; Kiser, J. Transformation of the oilseed crop Camelina sativa by Agrobacterium-mediated floral dip and simple large-scale screening of transformants. Vitr. Cell. Dev. Biol.-Plant 2012, 48, 462–468. [Google Scholar] [CrossRef]
- Lu, C.; Kang, J. Generation of transgenic plants of a potential oilseed crop Camelina sativa by Agrobacterium-mediated transformation. Plant Cell Rep. 2008, 27, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Sainger, M.; Jaiwal, A.; Sainger, P.A.; Chaudhary, D.; Jaiwal, R.; Jaiwal, P.K. Advances in genetic improvement of Camelina sativa for biofuel and industrial bio-products. Renew. Sustain. Energy Rev. 2017, 68, 623–637. [Google Scholar] [CrossRef]
- Sitther, V.; Tabatabai, B.; Enitan, O.; Dhekney, S. Agrobacterium-mediated transformation of Camelina sativa for production of transgenic plants. J. Biol. Methods 2018, e83. [Google Scholar] [CrossRef] [Green Version]
- Yemets, A.I.; Boychuk, Y.N.; Shysha, E.N.; Rakhmetov, D.B.; Blume, Y.B. Establishment of in vitro culture, plant regeneration, and genetic transformation of Camelina sativa. Cytol. Genet. 2013, 47, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Ozseyhan, M.E.; Kang, J.; Mu, X.; Lu, C. Mutagenesis of the FAE1 genes significantly changes fatty acid composition in seeds of Camelina sativa. Plant Physiol. Biochem. 2018, 123, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Morineau, C.; Bellec, Y.; Tellier, F.; Gissot, L.; Kelemen, Z.; Nogué, F.; Faure, J.D. Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol. J. 2017, 15, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, S.; Bhattacharya, A.; Khare, S.K.; Kaushik, G. Camelina sativa: An Emerging Biofuel Crop; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–38. [Google Scholar]
- Dharavath, R.N.; Singh, S.; Chaturvedi, S.; Luqman, S. Camelina sativa (L.) Crantz A mercantile crop with speckled pharmacological activities. Ann. Phytomed. 2016, 5, 6–26. [Google Scholar] [CrossRef]
- Malik, M.R.; Tang, J.; Sharma, N.; Burkitt, C.; Ji, Y.; Mykytyshyn, M.; Bohmert-Tatarev, K.; Peoples, O.; Snell, K.D. Camelina sativa, an oilseed at the nexus between model system and commercial crop. Plant Cell Rep. 2018, 37, 1367–1381. [Google Scholar] [CrossRef]
- Moser, B.R. Camelina (Camelina sativa L.) oil as a biofuels feedstock: Golden opportunity or false hope? Lipid Technol. 2010, 22, 270–273. [Google Scholar] [CrossRef]
- Murphy, E.J. Camelina (Camelina sativa). In Industrial Oil Crops; McKeon, T., Hayes, D., Hildebrand, D., Weselake, R., Eds.; Elevier: London, UK, 2016; pp. 207–230. [Google Scholar]
- Obour, K.A.; Sintim, H.Y.; O’bend, E.; Jeliazkov, V.D. Oilseed Camelina (Camelina sativa L. Crantz): Production systems, prospects and challenges in the USA Great Plains. Adv. Plants Agric. Res. 2015, 2, 00043. [Google Scholar] [CrossRef] [Green Version]
- Waraich, E.A.; Ahmed, Z.; Ahmad, R.; Ashraf, M.Y.; Naeem, M.S.; Rengel, Z. ‘Camelina sativa’, a climate proof crop, has high nutritive value and multiple-uses: A review. Aust. J. Crop Sci. 2013, 7, 1551–1559. [Google Scholar]
- Zubr, J. Oil-seed crop: Camelina sativa. Ind. Crops Prod. 1997, 6, 113–119. [Google Scholar] [CrossRef]
- Fröhlich, A.; Rice, B. Evaluation of Camelina sativa oil as a feedstock for biodiesel production. Ind. Crops Prod. 2005, 21, 25–31. [Google Scholar] [CrossRef]
- Ciubota-Rosie, C.; Ruiz, J.R.; Ramos, M.J.; Pérez, Á. Biodiesel from Camelina sativa: A comprehensive characterisation. Fuel 2013, 105, 572–577. [Google Scholar] [CrossRef]
- Zaleckas, E.; Makarevičienė, V.; Sendžikienė, E. Possibilities of using Camelina sativa oil for producing biodiesel fuel. Transport 2012, 27, 60–66. [Google Scholar] [CrossRef]
- Agarwal, A.; Prakash, O.; Bala, M. Camelina sativa, a short gestation oilseed crop with biofuel potential: Opportunities for Indian scenario. Oil Crop Sci. 2021, 6, 114–121. [Google Scholar] [CrossRef]
- Domestication of Plants in the Old World, 4th ed.; Oxford University Press: Oxford, UK, 2012.
- Larsson, M. Cultivation and processing of Linum usitatissimum and Camelina sativa in southern Scandinavia during the Roman Iron Age. Veg. Hist. Archaeobotany 2013, 22, 509–520. [Google Scholar] [CrossRef]
- Dönmez, E.O.; Belli, O. Urartian plant cultivation at Yoncatepe (Van), eastern Turkey. Econ. Bot. 2007, 61, 290–298. [Google Scholar] [CrossRef]
- Carciumaru, M. Cultivarea plantelor in Dacia (Plants cultivation in Dacia). Thraco-Dacica 2007, 8, 1–6. [Google Scholar]
- Knorzer, K.H. Evolution and spread of Gold of Pleasure (Camelina sativa S.L.). Ber. Dtsch. Bot. Ges. 1978, 91, 187–195. [Google Scholar]
- Hutcheon, C.; Ditt, R.F.; Beilstein, M.; Comai, L.; Schroeder, J.; Goldstein, E.; Shewmaker, C.K.; Nguyen, T.; De Rocher, J.; Kiser, J. Polyploid genome of Camelina sativa revealed by isolation of fatty acid synthesis genes. BMC Plant Biol. 2010, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Gesch, R.W. Influence of genotype and sowing date on camelina growth and yield in the north central U.S. Ind. Crops Prod. 2014, 54, 209–215. [Google Scholar] [CrossRef]
- Neupane, D.; Solomon, J.K.Q.; McLennon, E.; Davison, J.; Lawry, T. Camelina production parameters response to different irrigation regimes. Ind. Crops Prod. 2020, 148, 112286. [Google Scholar] [CrossRef]
- Lohaus, R.H.; Neupane, D.; Mengistu, M.A.; Solomon, J.K.Q.; Cushman, J.C. Five-year field trial of eight Camelina sativa cultivars for biomass to be used in biofuel under irrigated conditions in a semi-arid climate. Agronomy 2020, 10, 562. [Google Scholar] [CrossRef]
- Blackshaw, R.; Johnson, E.; Gan, Y.; May, W.; McAndrew, D.; Barthet, V.; McDonald, T.; Wispinski, D. Alternative oilseed crops for biodiesel feedstock on the Canadian prairies. Can. J. Plant Sci. 2011, 91, 889–896. [Google Scholar] [CrossRef]
- Gugel, R.K.; Falk, K.C. Agronomic and seed quality evaluation of Camelina sativa in western Canada. Can. J. Plant Sci. 2006, 86, 1047–1058. [Google Scholar] [CrossRef]
- Urbaniak, S.D.; Caldwell, C.D.; Zheljazkov, V.D.; Lada, R.; Luan, L. The effect of seeding rate, seeding date and seeder type on the performance of Camelina sativa L. in the Maritime Provinces of Canada. Can. J. Plant Sci. 2008, 88, 501–508. [Google Scholar] [CrossRef]
- Beilstein, M.A.; Al-Shehbaz, I.A.; Kellogg, E.A. Brassicaceae phylogeny and trichome evolution. Am. J. Bot. 2006, 93, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Beilstein, M.A.; Al-Shehbaz, I.A.; Mathews, S.; Kellogg, E.A. Brassicaceae phylogeny inferred from phytochrome A and ndhF sequence data: Tribes and trichomes revisited. Am. J. Bot. 2008, 95, 1307–1327. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.R.; Donmez, A.A.; Beilstein, M.A.; Olsen, K.M. Phylogenetics of Camelina Crantz. (Brassicaceae) and insights on the origin of gold-of-pleasure (Camelina sativa). Mol. Phylogenetics Evol. 2018, 127, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.R.; Mandakova, T.; Lysak, M.A.; Al-Shehbaz, I.A. Camelina neglecta (Brassicaceae, Camelineae), a new diploid species from Europe. PhytoKeys 2019, 115, 51–57. [Google Scholar] [CrossRef]
- Ghamkhar, K.; Croser, J.; Aryamanesh, N.; Campbell, M.; Konkova, N.; Francis, C. Camelina (Camelina sativa (L). Crantz) as an alternative oilseed: Molecular and ecogeographic analyses. Genome 2010, 53, 558–567. [Google Scholar] [CrossRef]
- Kagale, S.; Koh, C.; Nixon, J.; Bollina, V.; Clarke, W.E.; Tuteja, R.; Spillane, C.; Robinson, S.J.; Links, M.G.; Clarke, C.; et al. The emerging biofuel crop Camelina sativa retains a highly undifferentiated hexaploid genome structure. Nat. Commun. 2014, 5, 3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galasso, I.A.; Braglia, M.L.; Ponzoni, E.; Breviario, D. Genomic fingerprinting of Camelina species using cTBP as molecular marker. Am. J. Plant Sci. 2015, 6, 1184–1200. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.L.; Lujan-Toro, B.E.; Sauder, C.A.; James, T.; Ohadi, S.; Hall, L.M. An update to the Canadian range, abundance, and ploidy of Camelina spp. (Brassicaceae) east of the Rocky Mountains. Botany 2017, 95, 405–417. [Google Scholar] [CrossRef]
- Francis, A.; Warwick, S.I. The biology of Canadian weeds. 142. Camelina alyssum (Mill.) thell. C. microcarpa andrz. ex DC.; C. sativa (L.) crantz. Can. J. Plant Sci. 2009, 89, 791–810. [Google Scholar] [CrossRef]
- Gehringer, A.; Friedt, W.; Luhs, W.; Snowdon, R.J. Genetic mapping of agronomic traits in false flax (Camelina sativa subsp. sativa). Genome 2006, 49, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Al-Shehbaz, I.A.; Beilstein, M.A. Flora of North America North of Mexico. N. Y. Oxf. 2010, 7, 451–453. [Google Scholar]
- Koch, M.A.; Kiefer, M.; German, D.A.; Al-Shehbaz, I.A.; Franzke, A.; Mummenhoff, K.; Schmickl, R. BrassiBase: Tools and biological resources to study characters and traits in the Brassicaceae version 1.1. Taxon 2012, 61, 1001–1009. [Google Scholar] [CrossRef]
- Chaudhary, R.; Koh, C.S.; Kagale, S.; Tang, L.; Wu, S.W.; Lv, Z.; Mason, A.S.; Sharpe, A.G.; Diederichsen, A.; Parkin, I.A.P. Assessing diversity in the Camelina genus provides insights into the genome structure of Camelina sativa. G3 Genes Genomes Genet. 2020, 10, 1297–1308. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.R.; Mandáková, T.; McKain, M.; Lysak, M.A.; Olsen, K.M. Chloroplast phylogenomics in Camelina (Brassicaceae) reveals multiple origins of polyploid species and the maternal lineage of C. sativa. Hortic. Res. 2022, 9, uhab050. [Google Scholar] [CrossRef] [PubMed]
- Mirek, Z. Genus Camelina in Poland—Taxonomy, Distribution and Habitats. Fragm. Florist. Geobot. 1981, 27, 445–503. [Google Scholar]
- Gesch, R.W.; Archer, D.W. Double-cropping with winter camelina in the northern corn belt to produce fuel and food. Ind. Crops Prod. 2013, 44, 718–725. [Google Scholar] [CrossRef]
- Gesch, R.W.; Cermak, S. Sowing date and tillage effects on fall-seeded camelina in the northern Corn Belt. Agron. J. 2011, 103, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.A.; Wells, M.S.; Anderson, K.; Gesch, R.W.; Forcella, F.; Wyse, D.L. Yield tradeoffs and nitrogen between pennycress, camelina, and soybean in relay- and double-crop systems. Agron. J. 2017, 109, 2128–2135. [Google Scholar] [CrossRef]
- Neupane, D.; Solomon, J.K.Q.; Davison, J.; Lawry, T. Nitrogen source and rate effects on grain and potential biodiesel production of camelina in the semiarid environment of northern Nevada. Glob. Change Biol. Bioenergy 2018, 10, 861–876. [Google Scholar] [CrossRef] [Green Version]
- Walia, M.K.; Wells, M.S.; Cubins, J.; Wyse, D.; Gardner, R.D.; Forcella, F.; Gesch, R. Winter camelina seed yield and quality responses to harvest time. Ind. Crops Prod. 2018, 124, 765–775. [Google Scholar] [CrossRef]
- Berti, M.; Wilckens, R.; Fischer, S.; Solis, A.; Johnson, B. Seeding date influence on camelina seed yield, yield components, and oil content in Chile. Ind. Crops Prod. 2011, 34, 1358–1365. [Google Scholar] [CrossRef]
- Neupane, D.; Solomon, J.K.Q.; McLennon, E.; Davison, J.; Lawry, T. Sowing date and sowing method influence on camelina cultivars grain yield, oil concentration, and biodiesel production. Food Energy Secur. 2019, 8, e00166. [Google Scholar] [CrossRef] [Green Version]
- Angelini, L.G.; Moscheni, E.; Colonna, G.; Belloni, P.; Bonari, E. Variation in agronomic characteristics and seed oil composition of new oilseed crops in central Italy. Ind. Crops Prod. 1997, 6, 313–323. [Google Scholar] [CrossRef]
- Vollmann, J.; Moritz, T.; Kargl, C.; Baumgartner, S.; Wagentristl, H. Agronomic evaluation of camelina genotypes selected for seed quality characteristics. Ind. Crops Prod. 2007, 26, 270–277. [Google Scholar] [CrossRef]
- Dai, A.; Zhao, T.; Chen, J. Climate Change and Drought: A Precipitation and Evaporation Perspective. Curr. Clim. Change Rep. 2018, 4, 301–312. [Google Scholar] [CrossRef]
- Naumann, G.; Alfieri, L.; Wyser, K.; Mentaschi, L.; Betts, R.A.; Carrao, H.; Spinoni, J.; Vogt, J.; Feyen, L. Global changes in drought conditions under different levels of warming. Geophys. Res. Lett. 2018, 45, 3285–3296. [Google Scholar] [CrossRef]
- McKenzie, B.A.; Smallfield, B.M.; Fasi, V.; Martin, R.J. Possible species for the production of biodiesel on marginal land. Agron. N. Z. 2011, 41, 97–108. [Google Scholar]
- Pavlista, A.D.; Isbell, T.A.; Baltensperger, D.D.; Hergert, G.W. Planting date and development of spring-seeded irrigated canola, brown mustard and camelina. Ind. Crops Prod. 2011, 33, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Hergert, G.W.; Margheim, J.F.; Pavlista, A.D.; Martin, D.L.; Isbell, T.A.; Supalla, R.J. Irrigation response and water productivity of deficit to fully irrigated spring camelina. Agric. Water Manag. 2016, 177, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Hergert, G.W.; Margheim, J.F.; Pavlista, A.D.; Martin, D.L.; Supalla, R.J.; Isbell, T.A. Yield, irrigation response, and water productivity of deficit to fully irrigated spring canola. Agric. Water Manag. 2016, 168, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Matteo, R.; D’Avino, L.; Ramirez-Cando, L.J.; Pagnotta, E.; Angelini, L.G.; Spugnoli, P.; Tavarini, S.; Ugolini, L.; Foschi, L.; Lazzeri, L. Camelina (Camelina sativa L. Crantz) under low-input management systems in northern Italy: Yields, chemical characterization and environmental sustainability. Ital. J. Agron. 2020, 15, 132–143. [Google Scholar] [CrossRef]
- Załuski, D.; Tworkowski, J.; Krzyżaniak, M.; Stolarski, M.J.; Kwiatkowski, J. The characterization of 10 spring Camelina genotypes grown in environmental conditions in North-eastern Poland. Agronomy 2020, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Lošák, T.; Hlusek, J.; Martinec, J.; Vollmann, J.; Peterka, J.; Filipcik, R.; Varga, L.; Ducsay, L.; Martensson, A. Effect of combined nitrogen and sulphur fertilization on yield and qualitative parameters of Camelina sativa [L.] Crtz. (false flax). Acta Agric. Scand. Sect. B-Soil Plant Sci. 2011, 61, 313–321. [Google Scholar] [CrossRef]
- Manore, D.; Yohanns, A. Evaluating growth, seed yield and yield attributes of Camelina (Camelina sativa L) in response to seeding rate and nitrogen fertilizer levels under irrigation condition, southern Ethiopia. Agric. For. Fish. 2019, 8, 31–35. [Google Scholar] [CrossRef]
- Obeng, E.; Obour, A.K.; Nelson, N.O.; Moreno, J.A.; Ciampitti, I.A.; Wang, D.; Durrett, T.P. Seed yield and oil quality as affected by Camelina cultivar and planting date. J. Crop Improv. 2019, 33, 202–222. [Google Scholar] [CrossRef]
- Amiri-Darban, N.; Nourmohammadi, G.; Rad, A.H.S.; Mirhadi, S.M.J.; Heravan, I.M. Potassium sulfate and ammonium sulfate affect quality and quantity of camelina oil grown with different irrigation regimes. Ind. Crops Prod. 2020, 148, 112308. [Google Scholar] [CrossRef]
- Zanetti, F.; Eynck, C.; Christou, M.; Krzyżaniak, M.; Righini, D.; Alexopoulou, E.; Stolarski, M.J.; Van Loo, E.N.; Puttick, D.; Monti, A. Agronomic performance and seed quality attributes of Camelina (Camelina sativa L. crantz) in multi-environment trials across Europe and Canada. Ind. Crops Prod. 2017, 107, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, Y.A.; Chen, C.; Lamb, P.; Afshar, R.K. Agronomic Evaluation of Camelina (Camelina sativa L. Crantz) Cultivars for Biodiesel Feedstock. BioEnergy Res. 2017, 10, 792–799. [Google Scholar] [CrossRef]
- Obour, A.K.; Obeng, E.; Mohammed, Y.A.; Ciampitti, I.A.; Durrett, T.P.; Aznar-Moreno, J.A.; Chen, C.C. Camelina seed yield and fatty acids as influenced by genotype and wnvironment. Agron. J. 2017, 109, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Obour, A.K.; Chen, C.C.; Sintim, H.Y.; McVay, K.; Lamb, P.; Obeng, E.; Mohammed, Y.A.; Khan, Q.; Afshar, R.K.; Zheljazkov, V.D. Camelina sativa as a fallow replacement crop in wheat-based crop production systems in the US Great Plains. Ind. Crops Prod. 2018, 111, 22–29. [Google Scholar] [CrossRef]
- Pavlista, A.D.; Hergert, G.W.; Margheim, J.M.; Isbell, T.A. Growth of spring camelina (Camelina sativa) under deficit irrigation in western Nebraska. Ind. Crops Prod. 2016, 83, 118–123. [Google Scholar] [CrossRef] [Green Version]
- French, A.N.; Hunsaker, D.; Thorp, K.; Clarke, T. Evapotranspiration over a camelina crop at Maricopa, Arizona. Ind. Crops Prod. 2009, 29, 289–300. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; French, A.N.; Clarke, T.R.; El-Shikha, D.M. Water use, crop coefficients, and irrigation management criteria for camelina production in arid regions. Irrig. Sci. 2011, 29, 27–43. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; French, A.N.; Thorp, K.R. Camelina water use and seed yield response to irrigation scheduling in an arid environment. Irrig. Sci. 2012, 31, 911–929. [Google Scholar] [CrossRef] [Green Version]
- George, N.; Thompson, S.E.; Hollingsworth, J.; Orloff, S.; Kaffka, S. Measurement and simulation of water-use by canola and camelina under cool-season conditions in California. Agric. Water Manag. 2018, 196, 15–23. [Google Scholar] [CrossRef]
- Bronson, K.F.; Hunsaker, D.J.; Thorp, K.R. Nitrogen fertilizer and irrigation effects on seed yield and oil in Camelina. Agron. J. 2019, 111, 1712–1719. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Caldwell, C.D.; Falk, K.C.; Lada, R.R.; MacDonald, D. Camelina yield and quality response to combined nitrogen and sulfur. Agron. J. 2013, 105, 1847–1852. [Google Scholar] [CrossRef]
- Malhi, S.S.; Johnson, E.N.; Hall, L.M.; May, W.E.; Phelps, S.; Nybo, B. Effect of nitrogen fertilizer application on seed yield, N uptake, and seed quality of Camelina sativa. Can. J. Soil Sci. 2014, 94, 35–47. [Google Scholar] [CrossRef]
- Martinez, S.; Alvarez, S.; Capuano, A.; del Mar Delgado, M. Environmental performance of animal feed production from Camelina sativa (L.) Crantz: Influence of crop management practices under Mediterranean conditions. Agric. Syst. 2020, 177, 102717. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J. Life cycle assessment of camelina and crambe production for biorefinery and energy purposes. J. Clean. Prod. 2019, 237, 117755. [Google Scholar] [CrossRef]
- Sintim, H.Y.; Zheljazkov, V.D.; Obour, A.K.; Garcia y Garcia, A.; Foulke, T.K. Influence of nitrogen and sulfur application on camelina performance under dryland conditions. Ind. Crops Prod. 2015, 70, 253–259. [Google Scholar] [CrossRef]
- Kemp, W.H. Biodiesel: Basics and Beyond, a Comprehensive Guide to Production and Use for the Home and Farm; Aztext Press: Tamworth, ON, Canada, 2006. [Google Scholar]
- Bobrecka-Jamro, M.C. The effects of varied plant density and nitrogen fertilization on quantity and quality yield of Camelina sativa L. Emir. J. Food Agric. 2018, 29, 988–993. [Google Scholar] [CrossRef]
- Keshavarz-Afshar, R.; Mohammed, Y.A.; Chen, C. Energy balance and greenhouse gas emissions of dryland camelina as influenced by tillage and nitrogen. Energy 2015, 91, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Rathke, G.-W.; Christen, O.; Diepenbrock, W. Effects of N source and rate on productivity and quality of winter oilseed rape (Brassica napus L.) grown in different crop rotations. Field Crops Res. 2005, 94, 103–113. [Google Scholar] [CrossRef]
- Leclère, M.; Jeuffroy, M.-H.; Butier, A.; Chatain, C.; Loyce, C. Controlling weeds in camelina with innovative herbicide-free crop management routes across various environments. Ind. Crops Prod. 2019, 140, 111605. [Google Scholar] [CrossRef]
- Lovett, J.V.; Jackson, H.F. Allelopathic activity of Camelina sativa (L.) Crantz inrelation to its phyllosphere bacteria. New Phytol. 1980, 86, 273–277. [Google Scholar] [CrossRef]
- Walsh, K.D.; Sanderson, D.; Hall, L.M.; Mugo, S.; Hills, M.J. Allelopathic effects of camelina (Camelina sativa) and canola (Brassica napus) on wild oat, flax and radish. Allelopath. J. 2014, 33, 83–96. [Google Scholar]
- Abbas, A.; Huang, P.; Hussain, S.; Saqib, M.; He, L.; Shen, F.; Du, D. Application of allelopathic phenomemna to enhance growth and production of camelina (Camelina sativa (L.). Appl. Ecol. Environ. Res. 2021, 19, 453–469. [Google Scholar] [CrossRef]
- Davis, P.B.; Maxwell, B.; Menalled, F.D. Impact of growing conditions on the competitive ability of Camelina sativa (L.) Crantz (Camelina). Can. J. Plant Sci. 2013, 93, 243–247. [Google Scholar] [CrossRef]
- Lenssen, A.W.; Iversen, W.M.; Sainju, U.M.; Caesar-TonThat, T.C.; Blodgett, S.L.; Allen, B.L.; Evans, R.G. Yield, pests, and water use of durum and selected crucifer oilseeds in two-year rotations. Agron. J. 2012, 104, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Julié-Galau, S.; Bellec, Y.; Faure, J.-D.; Tepfer, M. Evaluation of the potential for interspecific hybridization between Camelina sativa and related wild Brassicaceae in anticipation of field trials of GM camelina. Transgenic Res. 2014, 23, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-J.; Auer, C. Hybridization between Camelina sativa (L.) Crantz and common Brassica weeds. Ind. Crops Prod. 2020, 147, 112240. [Google Scholar] [CrossRef]
- Davis, P.B. The Invasion Potential and Competitive Ability of Camelina sativa (L.) Crantz (Camelina) in Rangeland Ecosystems; Montana State University-Bozeman, College of Agriculture: Bozeman, MT, USA, 2010. [Google Scholar]
- Enjalbert, J.; Johnson, J. Guide for Producing Dryland Camelina in Eastern Colorado; Colorado State University Extension: Fort Collins, CO, USA, 2011. [Google Scholar]
- WAPMC. Biomass Crop Assistance Program: Camelina Agronomy Sheet; NRCS: Washington, DC, USA, 2012. [Google Scholar]
- Sobiech, Ł.; Grzanka, M.; Kurasiak-Popowska, D.; Radzikowska, D. Phytotoxic effect of herbicides on various camelina [Camelina sativa (L.) Crantz] genotypes and plant chlorophyll fluorescence. Agriculture 2020, 10, 185. [Google Scholar] [CrossRef]
- Scheliga, M.; Petersen, J. Selectivity of herbicides in camelina (Camelina sativa (L.) Crtz.). Jul.-Kühn-Arch. 2016, 452, 154–162. [Google Scholar] [CrossRef]
- Jha, P.; Stougaard, R.N. Camelina (Camelina sativa) tolerance to selected preemergence herbicides. Weed Technol. 2013, 27, 712–717. [Google Scholar] [CrossRef]
- Walsh, D.T.; Babiker, E.M.; Burke, I.C.; Hulbert, S.H. Camelina mutants resistant to acetolactate synthase inhibitor herbicides. Mol. Breed. 2012, 30, 1053–1063. [Google Scholar] [CrossRef]
- Schillinger, W.F.; Wysocki, D.J.; Chastain, T.G.; Guy, S.O.; Karow, R.S. Camelina: Planting date and method effects on stand establishment and seed yield. Field Crops Res. 2012, 130, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Saucke, H.; Ackermann, K. Weed suppression in mixed cropped grain peas and false flax (Camelina sativa). Weed Res. 2006, 46, 453–461. [Google Scholar] [CrossRef]
- Almoussawi, A.; Lenoir, J.; Spicher, F.; Dupont, F.; Chabrerie, O.; Closset-Kopp, D.; Brasseur, B.; Kobaissi, A.; Dubois, F.; Decocq, G. Direct seeding associated with a mixture of winter cover crops decreases weed abundance while increasing cash-crop yields. Soil Tillage Res. 2020, 200, 104622. [Google Scholar] [CrossRef]
- Royo-Esnal, A.; Valencia-Gredilla, F. Camelina as a rotation crop for weed control in organic farming in a semiarid Mediterranean climate. Agriculture 2018, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Berti, M.; Gesch, R.; Johnson, B.; Ji, Y.; Seames, W.; Aponte, A. Double-and relay-cropping of energy crops in the northern Great Plains, USA. Ind. Crops Prod. 2015, 75, 26–34. [Google Scholar] [CrossRef]
- Berti, M.; Samarappuli, D.; Johnson, B.L.; Gesch, R.W. Integrating winter camelina into maize and soybean cropping systems. Ind. Crops Prod. 2017, 107, 595–601. [Google Scholar] [CrossRef]
- Chandra, K.; Sharma, D.K.; Meher, L.C.; Kulkarni, A.V.; Nasim, M. Studies of feasibility of intercropping of Camelina sativa in jatropha plantation in semi–arid climate in Andhra Pradesh, India. Res. J. Agric. For. Sci. 2014, 2, 23–26. [Google Scholar]
- Robinson, R.G. Camelina: A Useful Research Crop and a Potential Oilseed Crop. 1987, 579. Available online: https://conservancy.umn.edu/bitstream/handle/11299/141546/SB579.pdf?sequence=1 (accessed on 10 February 2022).
- Soroka, J.J.; Olivier, C.; Wist, T.J.; Grenkow, L. Present and potential impacts of insects on Camelina and Crambe. In Integrated Management of Insect Pests on Canola and Other Brassica Oilseed Crops; CABI: Boston, MA, USA, 2017; pp. 316–340. [Google Scholar]
- Nielsen, J.K.; Hansen, M.L.; Agerbirk, N.; Petersen, B.L.; Halkier, B.A. Responses of the flea beetles Phyllotreta nemorum and P. cruciferae to metabolically engineered Arabidopsis thaliana with an altered glucosinolate profile. Chemoecology 2001, 11, 75–83. [Google Scholar] [CrossRef]
- Pachagounder, P.; Lamb, R.J.; Bodnaryk, R.P. Resistance to the flea beetle Phyllotreta cruciferae (Coleoptera: Chrysomelidae) in false flax, Camelina sativa (Brassicaceae). Can. Entomol. 1998, 130, 235–240. [Google Scholar] [CrossRef]
- Henderson, A.E.; Hallett, R.H.; Soroka, J.J. Prefeeding behavior of the crucifer flea beetle, Phyllotreta cruciferae, on host and nonhost crucifers. J. Insect Behav. 2004, 17, 17–39. [Google Scholar] [CrossRef]
- Onyilagha, J.C.; Gruber, M.Y.; Hallett, R.H.; Holowachuk, J.; Buckner, A.; Soroka, J.J. Constitutive flavonoids deter flea beetle insect feeding in Camelina sativa L. Biochem. Syst. Ecol. 2012, 42, 128–133. [Google Scholar] [CrossRef]
- Soroka, J.; Olivier, C.; Grenkow, L.; Séguin-Swartz, G. Interactions between Camelina sativa (Brassicaceae) and insect pests of canola. Can. Entomol. 2015, 147, 193–214. [Google Scholar] [CrossRef]
- Cárcamo, H.; Olfert, O.; Dosdall, L.; Herle, C.; Beres, B.; Soroka, J. Resistance to cabbage seedpod weevil among selected Brassicaceae germplasm. Can. Entomol. 2007, 139, 658–669. [Google Scholar] [CrossRef]
- Jewett, F.G. Camelina sativa: For biofuels and bioproducts. Ind. Crops 2015, 9, 157–170. [Google Scholar] [CrossRef]
- Chesnais, Q.; Verzeaux, J.; Couty, A.; Le Roux, V.; Ameline, A. Is the oilseedcrop Camelina sativa a potential host for aphid pests? BioEnergy Res. 2015, 8, 91–99. [Google Scholar] [CrossRef]
- Séguin-Swartz, G.; Eynck, C.; Gugel, R.K.; Strelkov, S.E.; Olivier, C.Y.; Li, J.L.; Klein-Gebbinck, H.; Borhan, H.; Caldwell, C.D.; Falk, K.C. Diseases of Camelina sativa (false flax). Can. J. Plant Pathol. 2009, 31, 375–386. [Google Scholar] [CrossRef]
- Conn, K.L.; Tewari, J.P.; Dahiya, J.S. Resistance to Alternaria brassicae and phytoalexin-elicitation in rapeseed and other crucifers. Plant Sci. 1988, 56, 21–25. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar, V.D.; Haque, A.; Bhat, S.R.; Prakash, S.; Chopra, V.L. Brassica coenospecies: A rich reservoir for genetic resistance to leaf spot caused by Alternaria brassicae. Euphytica 2002, 125, 411–417. [Google Scholar] [CrossRef]
- Jejelowo, O.A.; Conn, K.L.; Tewari, J.P. Relationship between conidial concentration, germling growth, and phytoalexin production by Camelina sativa leaves inoculated with Alternaria brassicae. Mycol. Res. 1991, 95, 928–934. [Google Scholar] [CrossRef]
- Dixit, S.; Jangid, V.K.; Grover. Evaluation of physiological and molecular effect of variable virulence of Alternaria brassicae isolates in Brassica juncea, Sinapis alba and Camelina sativa. Plant Physiol. Biochem. 2020, 155, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Westman, A.L.; Kresovich, S.; Dickson, M.H. Regional variation in Brassica nigra and other weedy crucifers for disease reaction to Alternaria brassicicola and Xanthomonas campestris pv. campestris. Euphytica 1999, 106, 253–259. [Google Scholar] [CrossRef]
- Sigareva, M.A.; Earle, E.D. Camalexin induction in intertribal somatic hybrids between Camelina sativa and rapid-cycling Brassica oleracea. Theor. Appl. Genet. 1999, 98, 164–170. [Google Scholar] [CrossRef]
- Hansen, L.N. Intertribal somatic hybridization between rapid cycling Brassica oleracea L. and Camelina sativa (L.) Crantz. Euphytica 1998, 104, 173–179. [Google Scholar] [CrossRef]
- Browne, L.M.; Conn, K.L.; Ayer, W.A.; Tewari, J.P. The camalexins: New phytoalexins produced in the leaves of Camelina sativa (Cruciferae). Tetrahedron 1991, 47, 3909–3914. [Google Scholar] [CrossRef]
- Mir, Z.A.; Ali, S.; Shivaraj, S.M.; Bhat, J.A.; Singh, A.; Yadav, P.; Rawat, S.; Paplao, P.K.; Grover, A. Genome-wide identification and characterization of chitinase gene family in Brassica juncea and Camelina sativa in response to Alternaria brassicae. Genomics 2020, 112, 749–763. [Google Scholar] [CrossRef]
- Li, H.; Barbetti, M.J.; Sivasithamparam, K. Hazard from reliance on cruciferous hosts as sources of major gene-based resistance for managing blackleg (Leptosphaeria maculans) disease. Field Crops Res. 2005, 91, 185–198. [Google Scholar] [CrossRef]
- Paul, V.H.; Tewari, J.P.; Tewari, I.; Foller, I.; Henneken, M. Joint field experiment on cultivation, yield, and diseases of false flax (Camelina sativa) in Germany and Canada. IOBC WPRS BULLETIN 2000, 23, 205–208. [Google Scholar]
- Föller, I.; Henneken, M.; Paul, V.H. Occurrence of false flax diseases (Camelina sativa (L.) CRTZ.) in field trials in Germany in 1995 and 1996. Bull. OILB/SROP 1998, 21, 65–76. [Google Scholar]
- Putnam, M.L.; Serdani, M.; Ehrensing, D.; Curtis, M. Camelina infected by downy mildew (Hyaloperonospora camelinae) in the western United States: A first report. Plant Health Prog. 2009, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Purnamasari, M.; Cawthray, G.R.; Barbetti, M.J.; Erskine, W.; Croser, J.S. Camalexin production in Camelina sativa is independent of cotyledon resistance to Sclerotinia sclerotiorum. Plant Dis. 2015, 99, 1544–1549. [Google Scholar] [CrossRef] [Green Version]
- Eynck, C.; Séguin-Swartz, G.; Clarke, W.E.; Parkin, I.A.P. Monolignol biosynthesis is associated with resistance to Sclerotinia sclerotiorum in Camelina sativa. Mol. Plant Pathol. 2012, 13, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi-Leonel, R.; Mueller, D.; Harbach, C.; Tylka, G.; Leandro, L. Susceptibility of cover crop plants to Fusarium virguliforme, causal agent of soybean sudden death syndrome, and Heterodera glycines, the soybean cyst nematode. J. Soil Water Conserv. 2017, 72, 575–583. [Google Scholar] [CrossRef]
- Conn, K.L.; Browne, L.M.; Tewari, J.P.; Ayer, W.A. Resistance to Rhizoctonia solani and presence of antimicrobial compounds in Camelina sativa roots. J. Plant Biochem. Biotechnol. 1994, 3, 125–130. [Google Scholar] [CrossRef]
- Paul, V.H.; Badent, S.; Henneken, M.; Foller, I. Results on the occurrence and seed-borne nature of the new disease bacterial blight (Pseudomonas syringae pv. spec) on false flax (Camelina sativa). IOBC WPRS BULLETIN 2000, 23, 43–50. [Google Scholar]
- Broadbent, L.; Heathcote, G.D. Properties and host range of turnip crinkle, rosette and yellow mosaic viruses. Ann. Appl. Biol. 1958, 46, 585–592. [Google Scholar] [CrossRef]
- Hein, A. Transmission of turnip yellow mosaic virus through seed of Camelina sativa (gold of pleasure). Z. Pflanzenkrankh. Pflanzenschutz/J. Plant Dis. Prot. 1984, 91, 549–551. [Google Scholar]
- Graichen, K. Zur Bedeutung von Virusbefall fuer denAnbau von Winterraps und Leindotter. Mitt. Biol. Bundesanst. Land-Forstwirtsch 1995, 310, 102–108. [Google Scholar]
- Chesnais, Q.; Mauck, K.E.; Bogaert, F.; Bamière, A.; Catterou, M.; Spicher, F.; Brault, V.; Tepfer, M.; Ameline, A. Virus effects on plant quality and vector behavior are species specific and do not depend on host physiological phenotype. J. Pest Sci. 2019, 92, 791–804. [Google Scholar] [CrossRef] [Green Version]
- Eberle, C.A.; Thom, M.D.; Nemec, K.T.; Forcella, F.; Lundgren, J.G.; Gesch, R.W.; Riedell, W.E.; Papiernik, S.K.; Wagner, A.; Peterson, D.H. Using pennycress, camelina, and canola cash cover crops to provision pollinators. Ind. Crops Prod. 2015, 75, 20–25. [Google Scholar] [CrossRef]
- Thom, M.D.; Eberle, C.A.; Forcella, F.; Gesch, R.; Weyers, S.; Lundgren, J.G. Nectar production in oilseeds: Food for pollinators in an agricultural landscape. Crop Sci. 2016, 56, 727–739. [Google Scholar] [CrossRef]
- Bekuzarova, S.A.; Basiev, S.S.; Buyankin, V.I.; Kozirev, A.K.; Dulaev, T.A. Ecological significance of winter camelina in biological agriculture. J. Pharm. Sci. Res. 2018, 10, 893–895. [Google Scholar]
- Wagg, C.; van Erk, A.; Fava, E.; Comeau, L.-P.; Mitterboeck, T.F.; Goyer, C.; Li, S.; McKenzie-Gopsil, A.; Mills, A. Full-season cover crops and their traits that promote agroecosystem services. Agriculture 2021, 11, 830. [Google Scholar] [CrossRef]
- Ugrenović, V.; Filipović, V.; Jevremović, S.; Marjanović-Jeromela, A.; Popović, V.; Buntić, A.; Delić, D.I. Effect of Brassicaceae as cover crops. Sel. I Semen. 2019, 25, 1–8. [Google Scholar] [CrossRef]
- Abramovič, H.; Abram, V. Physico-chemical properties, composition and oxidative stability of Camelina sativa oil. Food Technol. Biotechnol. 2005, 43, 63–70. [Google Scholar]
- Rodríguez-Rodríguez, M.F.; Sánchez-García, A.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Characterization of the morphological changes and fatty acid profile of developing Camelina sativa seeds. Ind. Crops Prod. 2013, 50, 673–679. [Google Scholar] [CrossRef]
- Anderson, J.V.; Wittenberg, A.; Li, H.; Berti, M.T. High throughput phenotyping of Camelina sativa seeds for crude protein, total oil, and fatty acids profile by near infrared spectroscopy. Ind. Crops Prod. 2019, 137, 501–507. [Google Scholar] [CrossRef]
- Zubr, J. Dietary fatty acids and amino acids of Camelina sativa seed. J. Food Qual. 2003, 26, 451–462. [Google Scholar] [CrossRef]
- Zubr, J. Qualitative variation of Camelina sativa seed from different locations. Ind. Crops Prod. 2003, 17, 161–169. [Google Scholar] [CrossRef]
- Hotton, S.K.; Kammerzell, M.; Chan, R.; Hernandez, B.T.; Young, H.A.; Tobias, C.; McKeon, T.; Brichta, J.; Thomson, N.J.; Thomson, J.G. Phenotypic examination of Camelina sativa (L.) Crantz accessions from the USDA-ARS National Genetics Resource Program. Plants 2020, 9, 642. [Google Scholar] [CrossRef]
- Singer, S.D.; Zou, J.; Weselake, R.J. Abiotic factors influence plant storage lipid accumulation and composition. Plant Sci. 2016, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zubr, J.; Matthaus, B. Effects of growth conditions on fatty acids and tocopherols in Camelina sativa oil. Ind. Crops Prod. 2002, 15, 155–162. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J.; Tworkowski, J.; Puttick, D.; Eynck, C.; Załuski, D.; Kwiatkowski, J. Yield and seed composition of 10 spring camelina genotypes cultivated in the temperate climate of Central Europe. Ind. Crops Prod. 2019, 138, 111443. [Google Scholar] [CrossRef]
- Yang, J.; Caldwell, C.; Corscadden, K.; He, Q.S.; Li, J. An evaluation of biodiesel production from Camelina sativa grown in Nova Scotia. Ind. Crops Prod. 2016, 81, 162–168. [Google Scholar] [CrossRef]
- Moser, B.R.; Vaughn, S.F. Evaluation of alkyl esters from Camelina sativa oil as biodiesel and as blend components in ultra low-sulfur diesel fuel. Bioresour. Technol. 2010, 101, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Leung, D.Y.C. Optimization of biodiesel production from camelina oil using orthogonal experiment. Appl. Energy 2011, 88, 3615–3624. [Google Scholar] [CrossRef]
- Hrastar, R.; Petrišič, M.G.; Ogrinc, N.; Košir, I.J. Fatty acid and stable carbon isotope characterization of Camelina sativa oil: Implications for authentication. J. Agric. Food Chem. 2009, 57, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Voelker, T.; Kinney, A.J. Variations in the biosynthesis of seed-storage lipids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 335–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, H.; Sun, J.; Li, B.; Zhu, Q.; Chen, S.; Zhang, H. Arabidopsis fatty acid desaturase FAD2 is required for salt tolerance during seed germination and early seedling growth. PLoS ONE 2012, 7, e30355. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.A.; Choudhury, A.R.; Kancharla, P.K.; Arumugam, N. The FAD2 Gene in Plants: Occurrence, Regulation, and Role. Front Plant Sci. 2017, 8, 1789. [Google Scholar] [CrossRef] [Green Version]
- Bhunia, R.K.; Kaur, R.; Maiti, M.K. Metabolic engineering of fatty acid biosynthetic pathway in sesame (Sesamum indicum L.): Assembling tools to develop nutritionally desirable sesame seed oil. Phytochem. Rev. 2015, 15, 799–811. [Google Scholar] [CrossRef]
- Raziei, Z.; Kahrizi, D.; Rostami-Ahmadvandi, H. Effects of climate on fatty acid profile in Camelina sativa. Cell. Mol. Biol. 2018, 64, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Berti, M.T.; Wilckens, R.; Fischer, S.; Araos, R. Borage: A new crop for southern Chile. In Trends in New Crops and New Uses: Strengthin Diversity; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 501–505. [Google Scholar]
- Gilbertson, P.K.; Berti, M.T.; Johnson, B.L. Borage cardinal germination temperatures and seed development. Ind. Crops Prod. 2014, 59, 202–209. [Google Scholar] [CrossRef]
- Leclère, M.; Lorent, A.-R.; Jeuffroy, M.-H.; Butier, A.; Chatain, C.; Loyce, C. Diagnosis of camelina seed yield and quality across an on-farm experimental network. Eur. J. Agron. 2021, 122, 126190. [Google Scholar] [CrossRef]
- Borzoo, S.; Mohsenzadeh, S.; Moradshahi, A.; Kahrizi, D.; Zamani, H.; Zarei, M. Characterization of physiological responses and fatty acid compositions of Camelina sativa genotypes under water deficit stress and symbiosis with Micrococcus yunnanensis. Symbiosis 2021, 83, 79–90. [Google Scholar] [CrossRef]
- Singh, S.K.; Rajpurohit, B.; Singha, P. Camelina (Camelina sativa) seed. In Oilseeds: Health Attributes and Food Applications; Tanwar, B., Goyal, A., Eds.; Springer: Singapore, 2021; pp. 455–471. [Google Scholar]
- Karvonen, H.M.; Aro, A.; Tapola, N.S.; Salminen, I.; Uusitupa, M.I.; Sarkkinen, E.S. Effect of alpha-linolenic acid-rich Camelina sativa oil on serum fatty acid composition and serum lipids in hypercholesterolemic subjects. Metabolism 2002, 51, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Schwab, U.S.; Lankinen, M.A.; de Mello, V.D.; Manninen, S.M.; Kurl, S.; Pulkki, K.J.; Laaksonen, D.E.; Erkkilä, A.T. Camelina sativa oil, but not fatty fish or lean fish, improves serum lipid profile in subjects with impaired glucose metabolism—a randomized controlled trial. Mol. Nutr. Food Res. 2018, 62, 1700503. [Google Scholar] [CrossRef]
- Manninen, S.; Lankinen, M.; Erkkilä, A.; Nguyen, S.D.; Ruuth, M.; de Mello, V.; Öörni, K.; Schwab, U. The effect of intakes of fish and Camelina sativa oil on atherogenic and anti-atherogenic functions of LDL and HDL particles: A randomized controlled trial. Atherosclerosis 2019, 281, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Mello, V.D.; Dahlman, I.; Lankinen, M.; Kurl, S.; Pitkänen, L.; Laaksonen, D.E.; Schwab, U.; Erkkilä, A.T. The effect of different sources of fish and camelina sativa oil on immune cell and adipose tissue mRNA expression in subjects with abnormal fasting glucose metabolism: A randomized controlled trial. Nutr. Diabetes 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Saleh-Ghadimi, S.; Dehghan, P.; Farhangi, M.A.; Khoshbaten, M. Co-Supplementation of Camelina Oil and Prebiotic is more Effective in Improvement of Cardiometabolic Risk Factors and Mental Health in Patients with NAFLD: A Randomized Clinical Trial. Food Funct. 2021, in press. [Google Scholar] [CrossRef]
- Ergönül, P.G.; Özbek, Z.A. Cold pressed camelina (Camelina sativa L.) seed oil. In Cold Pressed Oils; Elsevier: Amsterdam, The Netherlands, 2020; pp. 255–266. [Google Scholar]
- Ibrahim, F.M.; El Habbasha, S. Chemical composition, medicinal impacts andcultivation of camelina (Camelina sativa): Review. Int. J. PharmTech Res. 2015, 8, 114–122. [Google Scholar]
- Ratusz, K.; Symoniuk, E.; Wroniak, M.; Rudzińska, M. Bioactive compounds, nutritional quality and oxidative stability of cold-pressed camelina (Camelina sativa L.) oils. Appl. Sci. 2018, 8, 2606. [Google Scholar] [CrossRef] [Green Version]
- Eidhin, D.N.; O’Beirne, D. Oxidative stability of camelina oil in salad dressings, mayonnaises and during frying. Int. J. Food Sci. + Technol. 2010, 45, 444–452. [Google Scholar] [CrossRef]
- Salminen, H.; Estévez, M.; Kivikari, R.; Heinonen, M. Inhibition of protein and lipid oxidation by rapeseed, camelina and soy meal in cooked pork meat patties. Eur. Food Res. Technol. Vol. 2006, 223, 461–468. [Google Scholar] [CrossRef]
- Cherian, G. Camelina sativa in poultry diets: Opportunities and challenges. In Biofuel Co-Products as Livestock Feed: Opportunities and Challenges; Harinder, P.S., Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; pp. 303–310. [Google Scholar]
- Bătrîna, Ș.L.; Jurcoane, Ș.; Popescu, I.; Marin, F.; Imbrea, I.M.; Crista, F.; Pop, G.; Imbrea, F. Camelina sativa: A study on amino acid content. Rom. Biotechnol Lett. 2020, 25, 1136–1142. [Google Scholar] [CrossRef]
- Singh, B.K.; Bala, M.; Rai, P.K. Fatty acid composition and seed meal characteristics of Brassica and allied genera. Natl. Acad. Sci. Lett. 2014, 37, 219–226. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Glucosinolates and sinapine in camelina meal. Food Nutr. Sci. 2017, 8, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Paula, E.M.; da Silva, L.G.; Brandao, V.L.N.; Dai, X.; Faciola, A.P. Feeding canola, camelina, and carinata meals to ruminants. Animals 2019, 9, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, H.; Castillejos, L.; López-Suárez, M.; Ferret, A. In vitro digestibility, in situ degradability, rumen fermentation and N metabolism of camelina co-products for beef cattle studied with a dual flow continuous culture system. Animals 2019, 9, 1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriel, P.; Nayigihugu, V.; Cappellozza, B.I.; Gonçalves, E.P.; Krall, J.M.; Foulke, T.; Cammack, K.M.; Hess, B.W. Camelina meal and crude glycerin as feed supplements for developing replacement beef heifers. J. Anim. Sci. 2011, 89, 4314–4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombini, S.; Broderick, G.A.; Galasso, I.; Martinelli, T.; Rapetti, L.; Russo, R.; Reggiani, R. Evaluation of Camelina sativa (L.) Crantz meal as an alternative protein source in ruminant rations. J. Sci. Food Agric. 2014, 94, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, D.A.; Rohwer, G.; Norberg, O.S.; Kimura, E.; Neibergs, J.S.; Fransen, S.C. Case study: Evaluating farm processed canola and Camelina meals as protein supplements for beef cattle. J. NACAA 2015, 8. Available online: https://www.nacaa.com/journal (accessed on 10 February 2022).
- Cappellozza, B.I.; Cooke, R.F.; Bohnert, D.W.; Cherian, G.; Carroll, J.A. Effects of camelina meal supplementation on ruminal forage degradability, performance, and physiological responses of beef cattle. J. Anim. Sci. 2012, 90, 4042–4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, R.D.; Anderson, J.L.; Clapper, J.A. Evaluation of camelina meal as a feedstuff for growing dairy heifers. J. Dairy Sci. 2016, 99, 6215–6228. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.D.; Anderson, J.L. Ruminal degradation and intestinal digestibility of camelina meal and carinata meal compared with other protein sources. Prof. Anim. Sci. 2018, 34, 10–18. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Antinutritive compounds in twelve Camelina sativa genotypes. Am. J. Plant Sci. 2012, 3, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Schill, S.R. Camelina meal approved for feedlot cattle. Biodiesel Mag. 2009. Available online: http://www.biodieselmagazine.com/articles/3837/camelinameal-approved-for-feedlot-cattle (accessed on 10 February 2022).
- Berhow, M.A.; Polat, U.; Glinski, J.A.; Glensk, M.; Vaughn, S.F.; Isbell, T.; Ayala-Diaz, I.; Marek, L.; Gardner, C. Optimized analysis and quantification of glucosinolates from Camelina sativa seeds by reverse-phase liquid chromatography. Ind. Crops Prod. 2013, 43, 119–125. [Google Scholar] [CrossRef]
- Russo, R.; Galasso, I.; Reggiani, R. Variability in glucosinolate content among Camelina species. Am. J. Plant Sci. 2014, 5, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Vetter, W.; Darwisch, V.; Lehnert, K. Erucic acid in Brassicaceae and salmon–An evaluation of the new proposed limits of erucic acid in food. NFS J. 2020, 19, 9–15. [Google Scholar] [CrossRef]
- Hurtaud, C.; Peyraud, J.L. Effects of feeding camelina (seeds or meal) on milk fatty acid composition and butter spreadability. J. Dairy Sci. 2007, 90, 5134–5145. [Google Scholar] [CrossRef]
- Halmemies-Beauchet-Filleau, A.; Kokkonen, T.; Lampi, A.M.; Toivonen, V.; Shingfield, K.J.; Vanhatalo, A. Effect of plant oils and camelina expeller on milk fatty acid composition in lactating cows fed diets based on red clover silage. J. Dairy Sci. 2011, 94, 4413–4430. [Google Scholar] [CrossRef] [PubMed]
- Halmemies-Beauchet-Filleau, A.; Shingfield, K.J.; Simpura, I.; Kokkonen, T.; Jaakkola, S.; Toivonen, V.; Vanhatalo, A. Effect of incremental amounts of camelina oil on milk fatty acid composition in lactating cows fed diets based on a mixture of grass and red clover silage and concentrates containing camelina expeller. J. Dairy Sci. 2017, 100, 305–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieslak, A.; Stanisz, M.; Wojtowski, J.; Pers-Kamczyc, E.; Szczechowiak, J.; El-Sherbiny, M.; Szumacher-Strabel, M. Camelina sativa affects the fattyacid contents in M. longissimus muscle of lambs. Eur. J. Lipid Sci. Technol. 2013, 115, 1258–1265. [Google Scholar] [CrossRef]
- Steppa, R.; Cieślak, A.; Szumacher-Strabel, M.; Bielińska-Nowak, S.; Bryszak, M.; Stanisz, M.; Szkudelska, K. Blood serum metabolic profile and fatty acid composition in sheep fed concentrates with Camelina sativa cake and distillers dried grains with solubles. Small Rumin. Res. 2017, 156, 20–26. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; Muir, S.K.; Plozza, T.E.; Kerr, M.G.; Brown, W.G.; Jacobs, J.L.; Knight, M.I. Lipid oxidation and colour stability of lamb and yearling meat (muscle longissimus lumborum) from sheep supplemented with Camelina-based diets after short-, medium-, and long-term storage. Antioxidants 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Szumacher-Strabel, M.; Cieślak, A.; Zmora, P.; Pers-Kamczyc, E.; Bielińska, S.; Stanisz, M.; Wójtowski, J. Camelina sativa cake improved unsaturated fatty acids in ewe’s milk. J. Sci. Food Agric. 2011, 91, 2031–2037. [Google Scholar] [CrossRef]
- Mierlita, D.; Daraban, S.; Lup, F.; Chereji, A. The effect of grazing management and camelina seed supplementation in the diet on milk performance and milk fatty acid composition of dairy ewes. J. Food Agric. Environ. 2011, 9, 368–373. [Google Scholar]
- Pikul, J.; Wójtowski, J.; Danków, R.; Teichert, J.; Czyżak-Runowska, G.; Cais-Sokolińska, D.; Cieślak, A.; Szumacher-Strabel, M.; Bagnicka, E. The effect of false flax (Camelina sativa) cake dietary supplementation in dairy goats on fatty acid profile of kefir. Small Rumin. Res. 2014, 122, 44–49. [Google Scholar] [CrossRef]
- Cais-Sokolińska, D.; Majcher, M.; Pikul, J.; Bielińska, S.; Czauderna, M.; Wójtowski, J. The effect of Camelina sativa cake diet supplementation on sensory and volatile profiles of ewe’s milk. Afr. J. Biotechnol. 2011, 10, 7245–7252. [Google Scholar] [CrossRef]
- Cais-Sokolińska, D.; Wójtowski, J.; Pikul, J.; Danków, R.; Majcher, M.; Teichert, J.; Bagnicka, E. Formation of volatile compounds in kefir made of goat and sheep milk with high polyunsaturated fatty acid content. J. Dairy Sci. 2015, 98, 6692–6705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierlita, D.; Vicas, S. Dietary effect of silage type and combination with camelina seed on milk fatty acid profile and antioxidant capacity of sheep milk. S. Afr. J. Anim. Sci. 2015, 45, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Christodoulou, C.; Mavrommatis, A.; Mitsiopoulou, C.; Symeon, G.; Dotas, V.; Sotirakoglou, K.; Kotsampasi, B.; Tsiplakou, E. Assessing the optimum level of supplementation with Camelina seeds in ewes’ diets to improve milk quality. Foods 2021, 10, 2076. [Google Scholar] [CrossRef] [PubMed]
- Danków, R.; Pikul, J.; Wójtowski, J.; Cais-Sokolińska, D.; Teichert, J.; Bagnicka, E.; Cieślak, A.; Szumacher-Strabel, M. The effect of supplementation with gold of pleasure (Camelina sativa) cake on the fatty acid profile of ewe milk and yoghurt produced from it. J. Anim. Feed Sci. 2015, 24, 193–202. [Google Scholar] [CrossRef]
- Mierlita, D. Conjugated linoleic acid and ω-3 fatty acid in sheep milk was increased by part-time grazing and camelina seed diet. J. Food Agric. Environ. 2015, 13, 76–81. [Google Scholar]
- Eidhin, D.N.; Burke, J.; Lynch, B.; O’Beirne, D. Effects of dietary supplementation with camelina oil on porcine blood lipids. J. Food Sci. 2003, 68, 671–679. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Hebean, V.; Țăranu, I.; Marin, D.; Ropotă, M.; Tamaș, V. Effects of the dietary Camelina oil on large white pigs’ meat quality. Arch. Zootech. 2009, 12, 31–45. [Google Scholar]
- Hăbeanu, M.; Hebean, V.; Taranu, I.; Ropota, M.; Lefter, N.; Marin, D. Dietary ecologic Camelina oil: A beneficial aource of n-3 PUFA in muscle tissue and health status in finishing pigs. Rom. Biotechnol. Lett. 2011, 16, 5564–5571. [Google Scholar]
- Taranu, I.; Gras, M.; Pistol, G.C.; Motiu, M.; Marin, D.E.; Lefter, N.; Ropota, M.; Habeanu, M. ω-3 PUFA rich Camelina oil by-products improve the systemic metabolism and spleen cell functions in fattening pigs. PLoS ONE 2014, 9, e110186. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.N.; Htoo, J.K.; Thomson, J.; Stein, H.-H. Amino acid digestibility in camelina products fed to growing pigs. Can. J. Anim. Sci. 2013, 93, 335–343. [Google Scholar] [CrossRef]
- Kahindi, R.K.; Woyengo, T.A.; Thacker, P.A.; Nyachoti, C.M. Energy and amino acid digestibility of camelina cake fed to growing pigs. Anim. Feed Sci. Technol. 2014, 193, 93–101. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Lefter, A.N.; Ropotă, M. Effects of the camelina cake on fatty acids composition in different organs of the pigs. Arch. Zootech. 2016, 19, 5–16. [Google Scholar]
- Smit, M.N.; Beltranena, E. Effects of feeding camelina cake to weaned pigs on safety, growth performance, and fatty acid composition of pork. J. Anim. Sci. 2017, 95, 2496–2508. [Google Scholar] [CrossRef] [PubMed]
- Meadus, W.J.; Duff, P.; McDonald, T.; Caine, W.R. Pigs fed camelina meal increase hepatic gene expression of cytochrome 8b1, aldehyde dehydrogenase, and thiosulfate transferase. J. Anim. Sci. Biotechnol. 2014, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilbrands, A.M.; Johnston, L.J.; Cox, R.B.; Forcella, F.; Gesch, R.; Li, Y.Z. Effects of increasing dietary inclusion of camelina cake on growth performance of growing-finishing pigs. Transl. Anim. Sci. 2021, 5, 1–10. [Google Scholar] [CrossRef]
- Kim, J.W.; Koo, B.; Nyachoti, C.M. Digestible, metabolizable, and net energy of camelina cake fed to growing pigs and additivity of energy in mixed diets. J. Anim. Sci. 2017, 95, 4037–4044. [Google Scholar] [CrossRef]
- Juodka, R.; Nainienė, R.; Juškienė, V.; Juška, R.; Leikus, R.; Kadžienė, G.; Stankevičienė, D. Camelina (Camelina sativa (L.) Crantz) as feedstuffs in meat type poultry diet: A source of protein and n-3 fatty acids. Animals 2022, 12, 295. [Google Scholar] [CrossRef]
- Aziza, A.E.; Quezada, N.; Cherian, G. Feeding Camelina sativa meal to meat-type chickens: Effect on production performance and tissue fatty acid composition. J. Appl. Poult. Res. 2010, 19, 157–168. [Google Scholar] [CrossRef]
- Aziza, A.; Panda, A.; Quezada, N.; Cherian, G. Nutrient digestibility, egg quality, and fatty acid composition of brown laying hens fed camelina or flaxseed meal. J. Appl. Poult. Res. 2013, 22, 832–841. [Google Scholar] [CrossRef]
- Nain, S.; Oryschak, M.A.; Betti, M.; Beltranena, E. Camelina sativa cake for broilers: Effects of increasing dietary inclusion from 0 to 24% on tissue fatty acid proportions at 14, 28, and 42 d of age. Poult. Sci. 2015, 94, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Patterson, R.; Slominski, B.A.; Beltranena, E.; Zijlstra, R.T. Nutritive value of cold-pressed camelina cake with or without supplementation of multi-enzyme in broiler chickens. Poult. Sci. 2016, 95, 2314–2321. [Google Scholar] [CrossRef] [PubMed]
- Jáskiewicz, T.; Sagan, A.; Puzio, I. Effect of the Camelina sativa oil on theperformance, essential fatty acid level in tissues and fat-soluble vitamins content in the livers of broiler chickens. Livest. Sci. 2014, 165, 74–79. [Google Scholar] [CrossRef]
- Pietras, M.P.; Orczewska-Dudek, S. The effect of dietary Camelina sativa oil on quality of broiler chicken meat. Ann. Anim. Sci. 2013, 13, 869–882. [Google Scholar] [CrossRef]
- Orczewska-Dudek, S.; Pietras, M. The effect of dietary Camelina sativa oil or cake in the diets of broiler chickens on growth performance, fatty acid profile, and sensory quality of meat. Animals 2019, 9, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juodka, R.; Juska, R.; Juskiene, V.; Leikus, R.; Stankeviciene, D.; Nainiene, R. The effect of feeding with hemp and Camelina cakes on the fatty acid profile of duck muscles. Arch. Anim. Breed. 2018, 61, 293–303. [Google Scholar] [CrossRef]
- Frame, D.D.; Palmer, M.; Peterson, B. Use of Camelina sativa in the diets of young turkeys. J. Appl. Poult. Res. 2007, 16, 381–386. [Google Scholar] [CrossRef]
- Pekel, A.Y.; Kim, J.I.; Chapple, C.; Adeola, O. Nutritional characteristics of camelina meal for 3-week-old broiler chickens. Poult Sci 2015, 94, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Ryhänen, E.L.; Perttilä, S.; Tupasela, T.; Valaja, J.; Eriksson, C.; Larkka, K.E. Effect of Camelina sativa expeller cake on performance and meat quality of broilers. J. Sci. Food Agric. 2007, 87, 1489–1494. [Google Scholar] [CrossRef]
- Bulbul, T.; Rahman, A.; Ozdemir, V. Effect of false flax meal on certain growth, serum and meat parameters of japanese quails. J. Anim. Plant Sci. 2015, 25, 1245–1250. [Google Scholar]
- Ye, C.L.; Anderson, D.M.; Lall, S.P. The effects of camelina oil and solvent extracted camelina meal on the growth: Carcass composition and hindguthistology of Atlantic salmon (Salmo salar) parr in freshwater. Aquaculture 2016, 450, 397–404. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Full substitution of fish oil with camelina (Camelina sativa) oil, with partial substitution of fish meal with camelina meal, in diets for farmed Atlantic salmon (Salmo salar) and its effect on tissue lipids and sensory quality. Food Chem. 2014, 157, 51–61. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Effect of replacement of fish oil with camelina (Camelina sativa) oil on growth, lipid class and fatty acid composition of farmed juvenile Atlantic cod (Gadus morhua). Fish Physiol. Biochem. 2013, 39, 1441–1456. [Google Scholar] [CrossRef] [PubMed]
- Hixson, S.M.; Parrish, C.C.; Wells, J.S.; Marie Winkowski, E.; Anderson, D.M. Inclusion of camelina meal as a protein source in diets for farmed Atlantic cod Gadus morhua. Aquac. Res. 2016, 47, 2607–2622. [Google Scholar] [CrossRef]
- Ruiz-Lopez, N.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Successful high-level accumulation of fish oil omega-3 long-chain polyunsaturated fatty acids in a transgenic oilseed crop. Plant J. 2014, 77, 198–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betancor, M.B.; MacEwan, A.; Sprague, M.; Gong, X.; Montero, D.; Han, L.; Napier, J.A.; Norambuena, F.; Izquierdo, M.; Tocher, D.R. Oil from transgenic Camelina sativa as a source of EPA and DHA in feed for European sea bass (Dicentrarchus labrax L.). Aquaculture 2021, 530, 735759. [Google Scholar] [CrossRef] [PubMed]
- Bullerwell, C.N.; Collins, S.A.; Lall, S.P.; Anderson, D.M. Growth performance, proximate and histological analysis of rainbow trout fed diets containing Camelina sativa seeds, meal (high-oil and solvent-extracted) and oil. Aquaculture 2016, 452, 342–350. [Google Scholar] [CrossRef]
- Liang, Y.; May, C.; Foon, C.; Ngan, M.; Hock, C.; Basiron, Y. The effect of natural and synthetic antioxidants on the oxidative stability of palm diesel. Fuel 2006, 85, 867–870. [Google Scholar] [CrossRef]
- ASTM. ASTM (D6751–15ce1) Standard Specification for Biodiesel Fuel BlendStock (B100) for Middle Distillate Fuels. Am. Soc. Test. Mater. 2015. Available online: https://dieselnet.com/tech/fuel_biodiesel_std.php (accessed on 10 February 2022).
- European Standard EN 14214. EN (14214) European Specifications for Biodiesel Fuels. Eur. Comm. Stand. 2012. Available online: https://en.wikipedia.org/wiki/EN_14214 (accessed on 10 February 2022).
- Soriano, N.U.; Narani, A. Evaluation of biodiesel rerived from Camelina sativa oil. J. Am. Oil Chem. Soc. 2011, 89, 917–923. [Google Scholar] [CrossRef]
- Miller, P.; Kumar, A. Techno-economic assessment of hydrogenation-derived renewable diesel production from canola and camelina. Sustain. Energy Technol. Assess. 2014, 6, 105–115. [Google Scholar] [CrossRef]
- Miller, P.; Kumar, A. Development of emission parameters and net energy ratio for renewable diesel from Canola and Camelina. Energy 2013, 58, 426–437. [Google Scholar] [CrossRef]
- Krohn, B.J.; Fripp, M. A life cycle assessment of biodiesel derived from the “niche filling” energy crop camelina in the USA. Appl. Energy 2012, 92, 92–98. [Google Scholar] [CrossRef]
- Corporan, E.; Edwards, T.; Shafer, L.; DeWitt, M.J.; Klingshirn, C.; Zabarnick, S.; West, Z.; Striebich, R.; Graham, J.; Klein, J. Chemical, thermal stability, sealswell, and emissions studies of alternative jet fuels. Energy Fuels 2011, 25, 955–966. [Google Scholar] [CrossRef]
- Sivakumar, D.; Vankeswaram, S.; Sakthikumar, R.; Raghunandan, B. Analysis on the atomization characteristics of aviation biofuel discharging from simplex swirl atomizer. Int. J. Multiph. Flow 2015, 72, 88–96. [Google Scholar] [CrossRef]
- Sivakumar, G.; Vail, D.R.; Xu, J.; Burner, D.M.; Lay, J.O.; Ge, X.; Weathers, P.J. Bioethanol and biodiesel: Alternative liquid fuels for future generations. Eng. Life Sci. 2010, 10, 8–18. [Google Scholar] [CrossRef]
- Agusdinata, D.B.; Zhao, F.; Ileleji, K.; DeLaurentis, D. Life cycle assessment of potential biojet fuel production in the United States. Environ. Sci. Technol. 2011, 45, 9133–9143. [Google Scholar] [CrossRef] [PubMed]
- Shonnard, D.R.; Williams, L.; Kalnes, T.N. Camelina-derived jet fuel and diesel: Sustainable advanced biofuels. Env. Prog Sustain Energy 2010, 29, 382–392. [Google Scholar] [CrossRef]
- Shila, J.; Johnson, M.E. Techno-economic analysis of Camelina-derived hydroprocessed renewable jet fuel within the US context. Appl. Energy 2021, 287, 116525. [Google Scholar] [CrossRef]
- Boateng, A.A.; Mullen, C.A.; Goldberg, N.M. Producing stable pyrolysis liquids from the oil-seed presscakes of mustard family plants: Pennycress (Thlaspi arvense L.) and Camelina (Camelina sativa). Energy Fuels 2010, 24, 6624–6632. [Google Scholar] [CrossRef]
- Hernando, H.; Fermoso, J.; Moreno, I.; Coronado, J.M.; Serrano, D.P.; Pizarro, P. Thermochemical valorization of camelina straw waste via fast pyrolysis. Biomass Convers. Biorefinery 2017, 7, 277–287. [Google Scholar] [CrossRef]
- Gómez-Monedero, B.; Bimbela, F.; Arauzo, J.; Faria, J.; Ruiz, M.P. Pyrolysis of red eucalyptus, camelina straw, and wheat straw in an ablative reactor. Energy Fuels 2015, 29, 1766–1775. [Google Scholar] [CrossRef]
- Mohammad, B.T.; Al-Shannag, M.; Alnaief, M.; Singh, L.; Singsaas, E.; Alkasrawi, M. Production of multiple biofuels from whole Camelina material: A renewable energy crop. BioResources 2018, 13, 4870–4883. [Google Scholar]
- Jankowski, K.J.; Sokólski, M. Spring camelina: Effect of mineral fertilization on the energy efficiency of biomass production. Energy 2021, 220, 119731. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Krzyżaniak, M.; Kwiatkowski, J.; Tworkowski, J.; Szczukowski, S. Energy and economic efficiency of camelina and crambe biomass production on a large-scale farm in north-eastern Poland. Energy 2018, 150, 770–780. [Google Scholar] [CrossRef]
- Kasetaite, S.; Ostrauskaite, J.; Grazuleviciene, V.; Svediene, J.; Bridziuviene, D. Camelina oil- and linseed oil-based polymers with bisphosphonate crosslinks. Appl. Polym. 2014, 131, 8536–8543. [Google Scholar] [CrossRef]
- Fox, N.J.; Stachowiak, G.W. Vegetable oil-based lubricants- a review of oxidation. Tribol. Int. 2007, 40, 1035–1046. [Google Scholar] [CrossRef]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Processing Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Kim, N.; Li, Y.; Sun, X.S. Epoxidation of Camelina sativa oil and peel adhesion properties. Ind. Crops Prod. 2015, 64, 1–8. [Google Scholar] [CrossRef]
- Nosal, H.; Nowicki, J.; Warzała, M.; Nowakowska-Bogdan, E.; Zarębska, M. Synthesis and characterization of alkyd resins based on Camelina sativa oil and polyglycerol. Prog. Org. Coat. 2015, 86, 59–70. [Google Scholar] [CrossRef]
- Kim, J.T.; Netravali, A.N. Non-food application of camelina meal: Development of sustainable and green biodegradable paper-camelina composite sheets and fibers. Polym. Compos. 2012, 33, 1969–1976. [Google Scholar] [CrossRef]
- Cao, Y.; Gu, Z.; Muthukumarappan, K.; Gibbons, W. Separation of glucosinolates from Camelina seed meal via membrane and acidic aluminum oxide column. J. Liq. Chromatogr. Relat. Technol. 2015, 38, 1273–1278. [Google Scholar] [CrossRef]
- Ma, Y.; Gentry, T.; Hu, P.; Pierson, E.; Gu, M.; Yin, S. Impact of brassicaceous seed meals on the composition of the soil fungal community and the incidence of Fusarium wilt on chili pepper. Appl. Soil Ecol. 2015, 90, 41–48. [Google Scholar] [CrossRef]
- Schuster, A.; Friedt, W. Glucosinolate content and composition as parameters of quality of Camelina seed. Ind. Crops Prod. 1998, 7, 297–302. [Google Scholar] [CrossRef]
- Matthäus, B.; Zubr, J. Variability of specific components in Camelina sativa oilseed cakes. Ind. Crops Prod. 2000, 12, 9–18. [Google Scholar] [CrossRef]
- Maršalkienė, N.; Žilėnaitė, L.; Karpavičienė, B. Oil content and composition in seeds of Camelina sativa and Crambe abyssinica cultivars. J. Ementol. 2020, 25, 1399–1412. [Google Scholar] [CrossRef]
- Zubr, J. Unique dietary oil from Camelina sativa seed. Agrafood Ind. Hi-Tech 2009, 20, 42–46. [Google Scholar]
- De Lorgeril, M.; Renaud, S.; Salen, P.; Monjaud, I.; Mamelle, N.; Martin, J.; Guidollet, J.; Touboul, P.; Delaye, J. Mediterranean alpha-linolenic acid-rich diet in secondary prevention of coronary heart disease. Lancet 1994, 343, 1454–1459. [Google Scholar] [CrossRef]
- Budin, J.T.; Breene, W.M.; Putnam, D.H. Some compositional properties of camelina (Camelina sativa L. Crantz) seeds and oils. J. Am. Oil Chem. Soc. 1995, 72, 309–315. [Google Scholar] [CrossRef]
- Onyilagha, J.; Bala, A.; Hallett, R.; Gruber, M.; Soroka, J.; Westcott, N. Leaf flavonoids of the cruciferous species, Camelina sativa, Crambe spp., Thlaspi arvense and several other genera of the family Brassicaceae. Biochem. Syst. Ecol. 2003, 31, 1309–1322. [Google Scholar] [CrossRef]
- Kumar, K.; Gupta, S.M.; Arya, M.C.; Nasim, M. In vitro antimicrobial and antioxidant activity of camelina seed extracts as potential source of bioactive compounds. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 521–526. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Ryńska, B.; Stuper-Szablewska, K. Analysis of distribution of selected bioactive compounds in Camelina sativa from seeds to pomace and oil. Agronomy 2019, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Cojocariu, R.O.; Balmus, I.-M.; Lefter, R.; Hritcu, L.; Ababei, D.C.; Ciobica, A.; Copaci, S.; Mot, S.E.L.; Copolovici, L.; Copolovici, D.M.; et al. Camelina sativa methanolic and ethanolic extract potential in alleviating oxidative stress, memory deficits, and affective impairments in stress exposure-based irritable bowel syndrome mouse models. Oxidative Med. Cell. Longev. 2020, 2020, 9510305. [Google Scholar] [CrossRef] [PubMed]
- Cojocariu, R.; Balmus, I.; Lefter, R.; Hritcu, L.; Ababei, D.; Ciobica, A.; Copaci, S.; Copolovici, L.; Copolovici, D.; Jurcoane, S. Beneficial effects of Camelina sativa oil on behavioural (memory, anxiety, depression and social-related) manifestations and oxidative stress parameters in a mice model of irritable bowel syndrome. Rom. Biotechnol. Lett. 2020, 25, 1532–1540. [Google Scholar] [CrossRef]
- Gao, L.; Caldwell, C.D.; Jiang, Y. Photosynthesis and growth of camelina and canola in response to water deficit and applied nitrogen. Crop Sci. 2018, 58, 393–401. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ahmad, R.; Ahmed, Z.; Ahmad, Z.; Barutcular, C.; Erman, M.; Cig, F.; Saneoka, H.; Öztürk, F. Comparative study of growth, physiology and yield attributes of camelina (Camelina sativa L.) and canola (Brassica napus L.) under different irrigation regimes. Pak. J. Bot. 2020, 52, 1537–1544. [Google Scholar] [CrossRef]
- Yadav, P.V.; Khatri, D.; Nasim, M. Salt and PEG induced osmotic stress tolerance at germination and seedling stage in Camelina sativa: A potential biofuel crop. J. Seed Sci. 2017, 10, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, Z.; Waraich, E.A.; Rashid, A.; Shahbaz, M. Morpho-physiological and biochemical responses of camelina (Camelina sativa Crantz) genotypes under drought stress. Int. J. Agric. Biol. 2017, 19, 1–7. [Google Scholar] [CrossRef]
- Čanak, P.; Jeromela, A.M.; Vujošević, B.; Kiprovski, B.; Mitrović, B.; Alberghini, B.; Facciolla, E.; Monti, A.; Zanetti, F. Is drought stress tolerance affected by biotypes and seed size in the emerging oilseed crop Camelina? Agronomy 2020, 10, 1856. [Google Scholar] [CrossRef]
- Ahmad, Z.; Anjum, S.; Skalicky, M.; Waraich, E.A.; Tariq, R.M.S.; Ayub, M.A.; Hossain, A.; Hassan, M.M.; Brestic, M.; Islam, M.S.; et al. Selenium alleviates the adverse effect of drought in oilseed crops Camelina (Camelina sativa L.) and Canola (Brassica napus L.). Molecules 2021, 26, 1699. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.; Waraich, E.A.; Barutcular, C.; Alharby, H.; Bamagoos, A.; Kizilgeci, F.; Öztürk, F.; Hossain, A.; Bayoumi, Y.; El Sabagh, A. Enhancing drought tolerance in Camelina sativa L. and canola (Brassica napus L.) through application of selenium. Pak. J. Bot. 2020, 52, 1927–1939. [Google Scholar] [CrossRef]
- Ahmad, M.; Waraich, E.A.; Zulfiqar, U.; Ullah, A.; Farooq, M. Thiourea application improves heat tolerance in camelina (Camelina sativa L. Crantz) by modulating gas exchange, antioxidant defense and osmoprotection. Ind. Crops Prod. 2021, 170, 113826. [Google Scholar] [CrossRef]
- Liang, C.; Liu, X.; Yiu, S.-M.; Lim, B.L. De novo assembly and characterization of Camelina sativa transcriptome by paired-end sequencing. BMC Genom. 2013, 14, 146. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.; Silva, J.E.; Podicheti, R.; Macrander, J.; Yang, W.; Nazarenus, T.J.; Nam, J.W.; Jaworski, J.G.; Lu, C.; Scheffler, B.E.; et al. Camelina seed transcriptome: A tool for meal and oil improvement and translational research. Plant Biotechnol. J. 2013, 11, 759–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, K.; Li, H.; Kang, J.; Lu, C. Mapping quantitative trait loci for seed traits in Camelina sativa. Theor. Appl. Genet. 2019, 132, 2567–2577. [Google Scholar] [CrossRef]
- Luo, Z.; Brock, J.; Dyer, J.M.; Kutchan, T.; Schachtman, D.; Augustin, M.; Ge, Y.; Fahlgren, N.; Abdel-Haleem, H. Genetic diversity and population structure of a Camelina sativa spring panel. Front. Plant Sci. 2019, 10, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Szczepanek, A.; Abdel-Haleem, H. Genome-wide association study (GWAS) analysis of Camelina seedling germination under salt stress sondition. Agronomy 2020, 10, 1444. [Google Scholar] [CrossRef]
- Vollmann, J.; A, D.; Baumgartner, S.; Ruckenbauer, P. Selection of induced mutants with improved linolenic acid content in camelina. Fett/Lipid 1997, 99, 357–361. [Google Scholar] [CrossRef]
- Lohaus, R.H.; Zager, J.J.; Kosma, D.K.; Cushman, J.C. Characterization of seed, oil, and fatty acid methyl esters of an ethyl methanesulfonate mutant of Camelina sativa with reduced seed-coat mucilage. J. Am. Oil Chem. Soc. 2020, 97, 157–174. [Google Scholar] [CrossRef]
- Lovett, J.V.; Sagar, G.R. Influence of bacteria in the phyllosphere of Camelina sativa (L.) Crantz on germination of Linum usitatissimum L. New Phytol. 1978, 81, 617–625. [Google Scholar] [CrossRef]
- Miyashita, H.; Fujita, K. Improvement of Camelina sativa yield using a plant growth-promoting rhizobacterium. Soil Sci. Plant Nutr. 2019, 65, 566–569. [Google Scholar] [CrossRef]
- Bekuzarova, S.A.; Weisfeld, L.I.; Dulaev, T.A. Biological methods increasing the productivity of the winter false flax Camelina silvestris L. In Antioxidants in Systems of Varying Complexity; Apple Academic Press: Palm Bay, FL, USA, 2019; pp. 293–304. [Google Scholar]
- Chhikara, S.; Abdullah, H.M.; Akbari, P.; Schnell, D.; Dhankher, O.P. Engineering Camelina sativa (L.) Crantz for enhanced oil and seed yields by combining diacylglycerol acyltransferase1 and glycerol-3-phosphate dehydrogenase expression. Plant Biotechnol J. 2018, 16, 1034–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heydarian, Z.; Yu, M.; Gruber, M.; Glick, B.R.; Zhou, R.; Hegedus, D.D. Inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression of the corresponding acdS gene in transgenic plants increases salinity tolerance in Camelina sativa. Front. Microbiol. 2016, 7, 1966. [Google Scholar] [CrossRef]
- Heydarian, Z.; Gruber, M.; Glick, B.R.; Hegedus, D.D. Gene expression patterns in roots of Camelina sativa with enhanced salinity tolerance arising from inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression the corresponding acds gene. Front. Microbiol. 2018, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Kanth, B.K.; Kumari, S.; Choi, S.H.; Ha, H.-J.; Lee, G.-J. Generation and analysis of expressed sequence tags (ESTs) of Camelina sativa to mine drought stress-responsive genes. Biochem. Biophys. Res. Commun. 2015, 467, 83–93. [Google Scholar] [CrossRef]
- Heydarian, Z.; Yu, M.; Gruber, M.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in gene expression in Camelina sativa roots and vegetative tissues in response to salinity stress. Sci. Rep. 2018, 8, 9804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.B.; Kim, H.; Kim, R.J.; Suh, M.C. Overexpression of Arabidopsis MYB96 confers drought resistance in Camelina sativa via cuticular wax accumulation. Plant Cell Rep. 2014, 33, 1535–1546. [Google Scholar] [CrossRef]
- Kim, R.J.; Kim, H.U.; Suh, M.C. Development of camelina enhanced with drought stress resistance and seed oil production by co-overexpression of MYB96A and DGAT1C. Ind. Crops Prod. 2019, 138, 111475. [Google Scholar] [CrossRef]
- Subedi, U.; Jayawardhane, K.N.; Pan, X.; Ozga, J.; Chen, G.; Foroud, N.A.; Singer, S.D. The potential of genome editing for improving seed oil content and fatty acid composition in oilseed crops. Lipids 2020, 55, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Kawall, K. Genome-edited Camelina sativa with a unique fatty acid content and its potential impact on ecosystems. Environ. Sci. Eur. 2021, 33, 38. [Google Scholar] [CrossRef]
- Yuan, L.; Li, R. Metabolic engineering a model oilseed Camelina sativa for the sustainable production of high-value designed oils. Front. Plant Sci. 2020, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Moreno, J.A.; Durrett, T.P. Simultaneous targeting of multiple gene homeologs to alter seed oil production in Camelina sativa. Plant Cell Physiol. 2017, 58, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-R.; Jeon, I.; Yu, H.; Kim, S.-G.; Kim, H.-S.; Ahn, S.-J.; Lee, J.; Lee, S.-K.; Kim, H.U. Increasing monounsaturated fatty acid contents in hexaploid Camelina sativa seed oil by FAD2 gene knockout using CRISPR-Cas9. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lyzenga, W.J.; Harrington, M.; Bekkaoui, D.; Wigness, M.; Hegedus, D.D.; Rozwadowski, K.L. CRISPR/Cas9 editing of three CRUCIFERIN C homoeologues alters the seed protein profile in Camelina sativa. BMC Plant Biol. 2019, 19, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veljković, V.B.; Kostić, M.D.; Stamenković, O.S. Camelina seed harvesting, storing, pretreating, and processing to recover oil: A review. Ind. Crops Prod. 2022, 178, 114539. [Google Scholar] [CrossRef]
- Pagnotta, E.; Ugolini, L.; Matteo, R.; Lazzeri, L.; Foschi, L.; Angelini, L.; Tavarini, S. Exploring the Camelina sativa value chain: A new opportunity for bio-based products and overall crop sustainability. La Riv. Italliana Delle Sostanze Grasse 2019, 96, 259–268. [Google Scholar]
- Righini, D.; Zanetti, F.; Monti, A. The bio-based economy can serve as the springboard for camelina and crambe to quit the limbo. Oilseeds Fats Crops Lipids (OCL) 2016, 23, D504. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Waraich, E.A.; Zulfiqar, U.; Ullah, A.; Farooq, M. Thiourea application increases seed and oil yields in Camelina under heat stress by modulating the plant water relations and antioxidant defense system. J. Soil Sci. Plant Nutr. 2022, 1–18. [Google Scholar] [CrossRef]


| Concentration of Major Fatty Acids (%) | Reference | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | C18:1 | C18:2 | C18:3 | C20:1 | C22:1 | SFA | MUFA | PUFA | |
| - | - | - | 21–23 | 27–29 | - | - | 11–12 | 35–36 | 51–52 | [75] |
| - | - | 14–16 | 15–23 | 31–40 | 12–15 | - | - | - | - | [2] |
| 5.3–5.6 | 2.2–2.7 | 14.7–16.5 | 12.9–16.3 | 35 | 15 | 3 | 12 | 34 | 54 | [159,160] |
| - | - | - | - | 30–43 | 11–19 | <3 | - | - | - | [6] |
| 5.4 | 2.4 | 14.3 | 20.6 | 36.9 | 13 | 2.2 | - | - | - | [3] |
| 6.29 | 2.73 | 16.5 | 17.7 | 32.5 | 15.6 | 3.1 | - | - | - | [64] |
| 5.16 | 2.68 | 15.2 | 17.9 | 34.6 | 15.1 | 2.6 | 8.6 | 33.0 | 54.1 | [26] |
| 5.68 | - | 13.9 | 16.6 | 35.1 | 14.3 | 3.0 | 10.1 | 33.5 | 55.9 | [164] |
| - | - | 9.1–22.1 | 15.2–27.1 | 22.8–38.4 | 11.6–18.2 | - | - | - | - | [158] |
| 5.5 | 2.4 | 14.4 | 19.1 | 33.5 | 15 | 3.1 | - | - | - | [165] |
| 7 | 2.5 | 6.9 | 14.5 | 41 | 10.9 | 3.5 | - | - | - | [157] |
| 6.8 | 2.7 | 18.6 | 19.6 | 32.6 | 12.4 | 2.3 | - | - | - | [166] |
| 5.1 | 2.4 | 17.6 | 18.7 | 28.6 | 11.9 | 4.2 | - | - | - | [167] |
| 5.4 | 2.6 | 14.3 | 14.3 | 38.4 | 16.8 | 2.9 | - | - | - | [25] |
| 5.7 | 3.4 | 15.0 | 18.5 | 34.7 | 12.7 | 3.2 | - | - | - | [168] |
| 4.6–5.2 | 2.2–2.5 | 12.8–14.7 | 16.3–17.2 | 36.2–39.4 | 14.0–15.5 | 2.5–3.1 | 8.7–8.8 | - | - | [39] |
| 5.5–9.5 | - | 9.1–17.1 | 16.1–28.6 | 23.5–36.2 | 10.5–16.4 | <2 | - | - | - | [161] |
| 6.2 | 2.6 | 16.2 | 17.5 | 37.3 | 13.1 | 2.3 | 9.7 | 31.7 | 57.7 | [71] |
| 5.8 | 2.6 | 14.7 | 18.3 | 34.4 | 14.0 | 2.9 | 10.1 | 33.5 | 54.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neupane, D.; Lohaus, R.H.; Solomon, J.K.Q.; Cushman, J.C. Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate. Plants 2022, 11, 772. https://doi.org/10.3390/plants11060772
Neupane D, Lohaus RH, Solomon JKQ, Cushman JC. Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate. Plants. 2022; 11(6):772. https://doi.org/10.3390/plants11060772
Chicago/Turabian StyleNeupane, Dhurba, Richard H. Lohaus, Juan K. Q. Solomon, and John C. Cushman. 2022. "Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate" Plants 11, no. 6: 772. https://doi.org/10.3390/plants11060772
APA StyleNeupane, D., Lohaus, R. H., Solomon, J. K. Q., & Cushman, J. C. (2022). Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate. Plants, 11(6), 772. https://doi.org/10.3390/plants11060772