
Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L.) under Iron (II) Sulfide

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
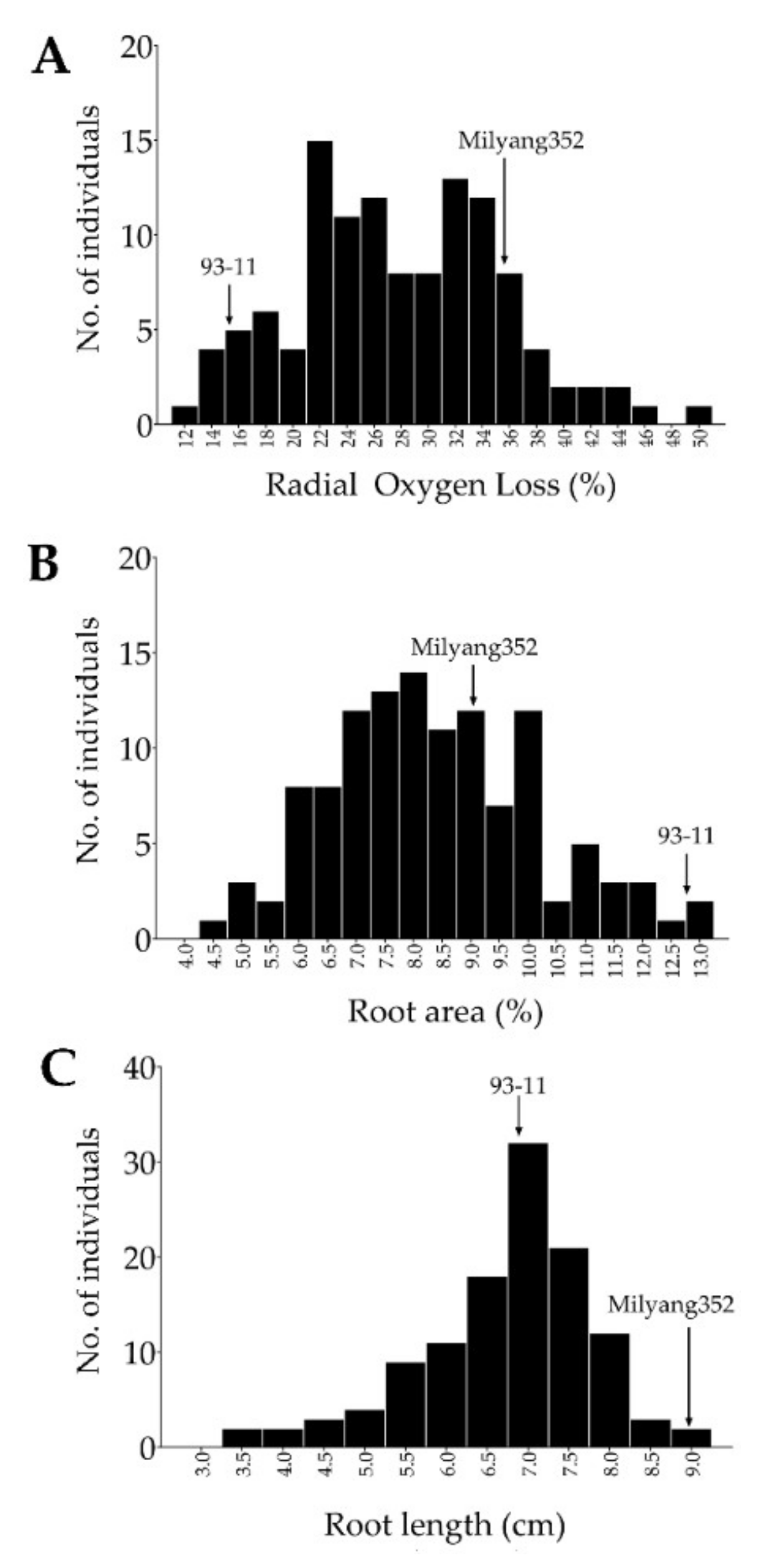
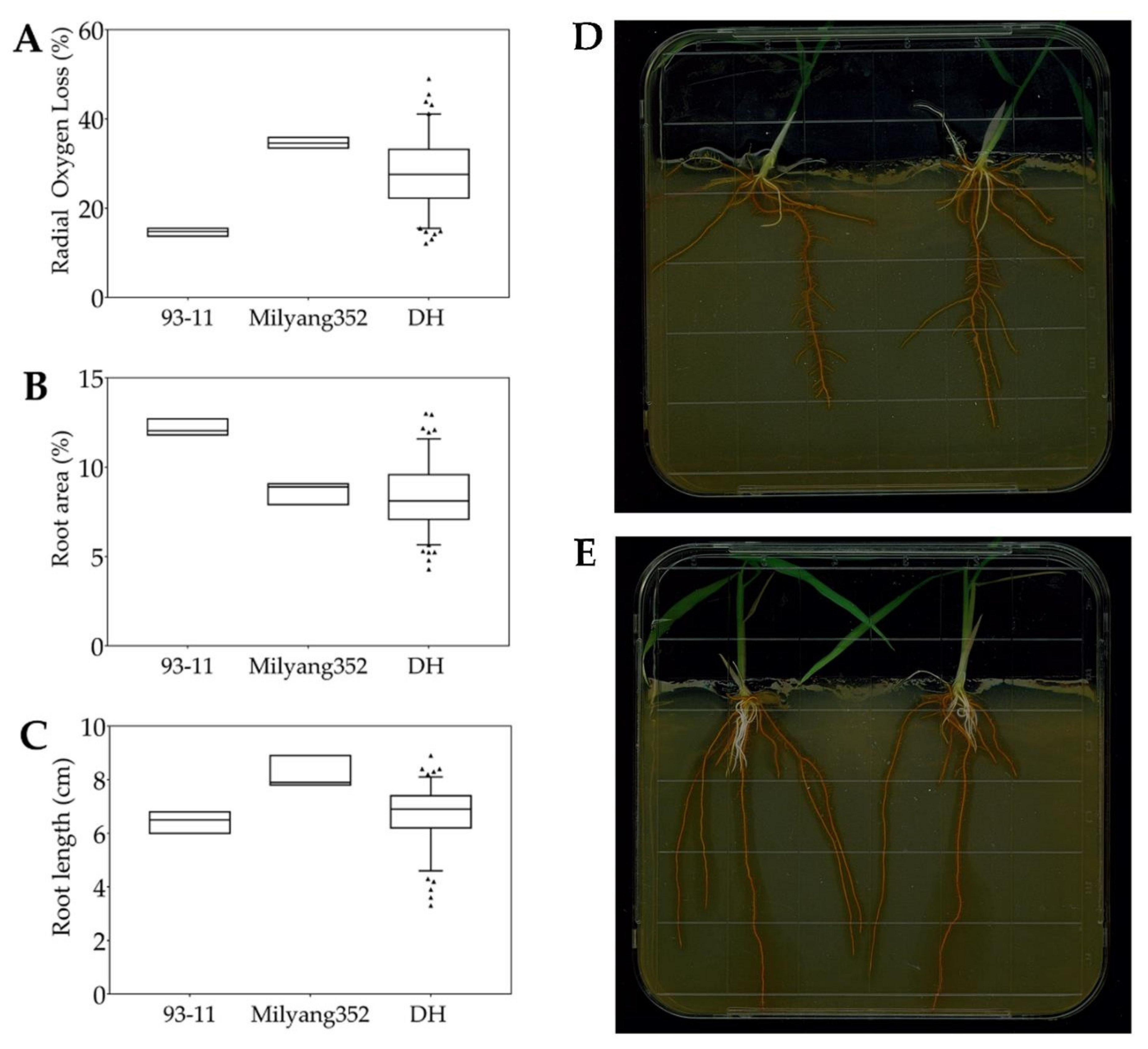
2.1. Growth Patterns of Doubled Haploid Lines in Response to FeS Treatment
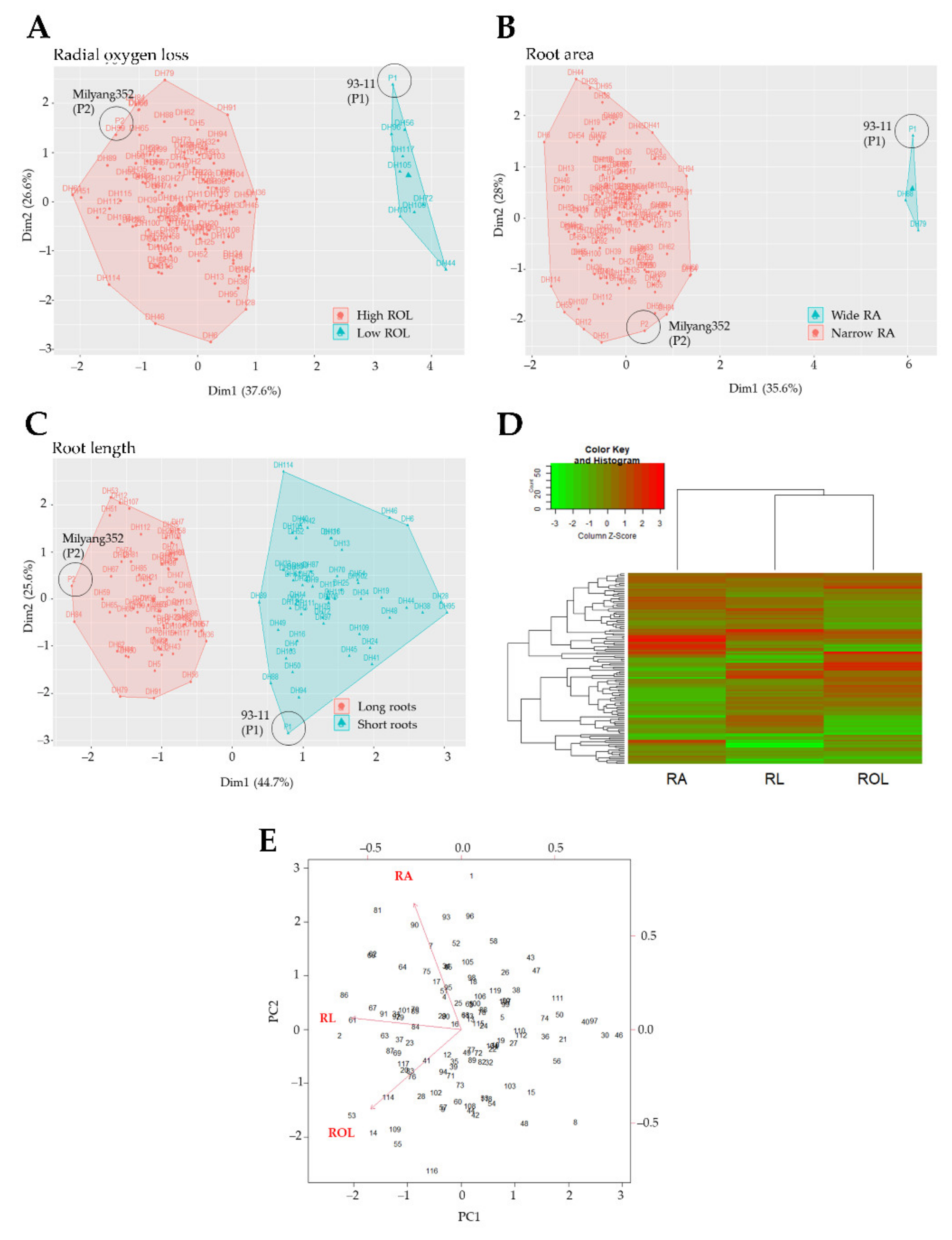
2.2. Principal Component Analysis (PCA) and Clustering of the Mapping Population
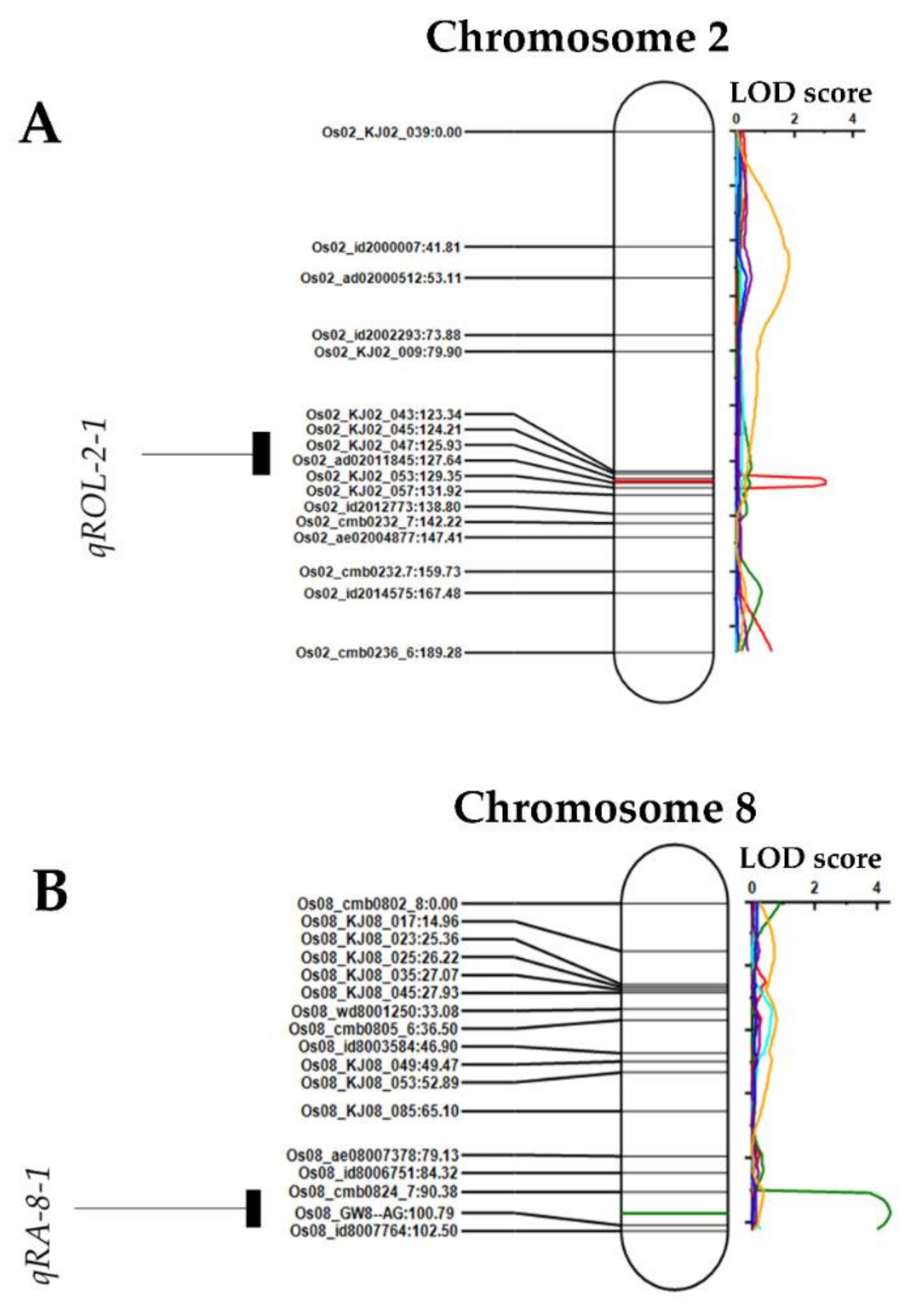
2.3. Quantitative Trait Loci (QTLs) Associated with Radial Oxygen Loss and Root Area under FeS in Rice
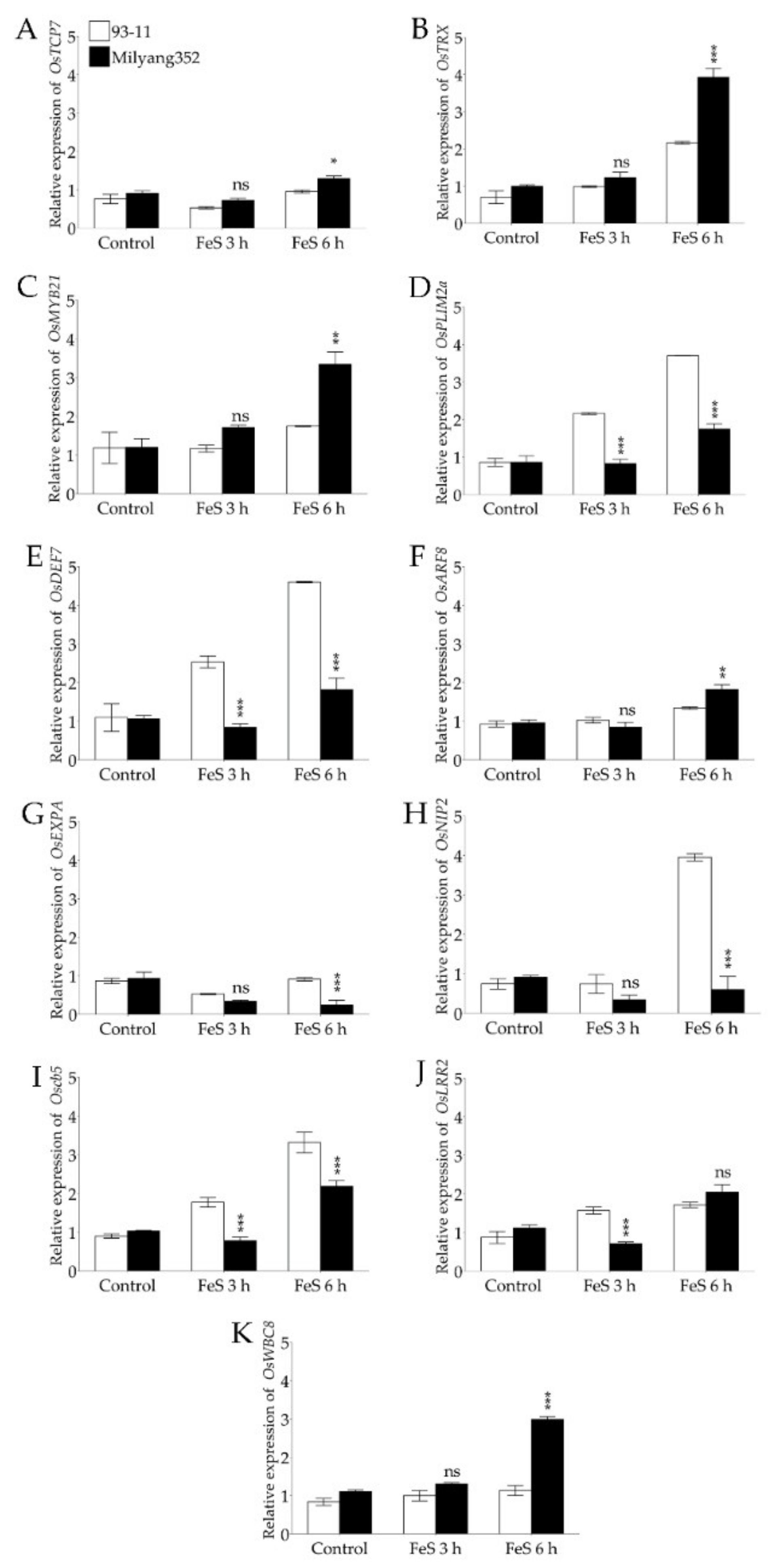
2.4. Validation of qROL-2-1-Related Genes Associated with Radial Oxygen Loss in Rice
3. Discussion
3.1. Differential Radial Oxygen Loss between Parental Lines and Doubled Haploid Rice Lines
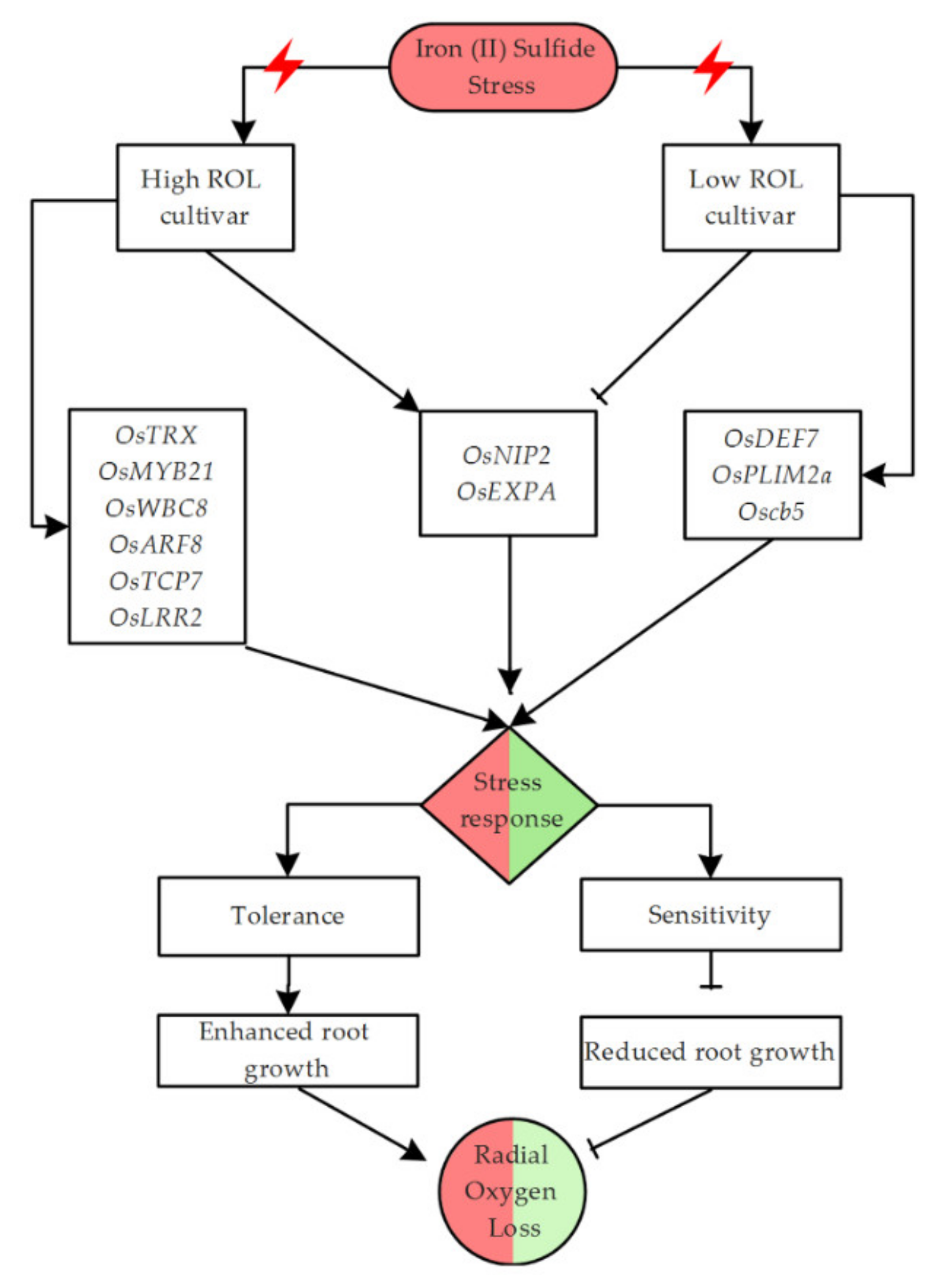
3.2. Stress-Responsive Genes Present within qROL-2-1 QTL Denotes Iron Sulfide-Induced Changes in ROL, May Involve Active Cellular Signaling Networks and Transcriptional Regulation in Rice
3.3. Other Candidate Genes with the Potential to Control Radial Oxygen Loss in Rice
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and FeS Medium Preparation
4.2. Determination of Root Oxidized Area, Radial Oxygen Loss, and Root Length
4.3. Frequency Distribution, Box Plots, and Principal Component Analysis
4.4. Construction of Linkage Mapping and QTL Analysis
4.5. qPCR Validation of Putative ROL Candidate Genes in Response to FeS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, C.; Motohashi, R.; Tsuchimoto, S.; Fukuta, Y.; Ohtsubo, H.; Ohtsubo, E. Polyphyletic Origin of Cultivated Rice: Based on the Interspersion Pattern of SINEs. Mol. Biol. Evol. 2003, 20, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lu, T.; Yu, S.; Li, Y.; Huang, Y.; Huang, T.; Zhang, L.; Zhu, J.; Zhao, Q.; Fan, D.; et al. A collection of 10,096 indica rice full-length cDNAs reveals highly expressed sequence divergence between Oryza sativa indica and japonica subspecies. Plant Mol. Biol. 2007, 65, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Colowit, P.M.; Mackill, D.J. Evaluation of Genetic Diversity in Rice Subspecies Using Microsatellite Markers. Crop Sci. 2002, 42, 601–607. [Google Scholar] [CrossRef]
- Sasaki, T.; Ashikari, M. Rice Genomics, Genetics and Breeding; Springer: Singapore, 2018; pp. 191–206. [Google Scholar]
- Nikkhah, A. Rice for Ruminants: Race for a Science under Shadow. Rice Res. Open Access 2015, 3, 134. [Google Scholar] [CrossRef]
- Asyifah, M.N.; Abd-Aziz, S.; Phang, L.Y.; Azlian, M.N. Brown rice as a potential feedstuff for poultry. J. Appl. Poult. Res. 2012, 21, 103–110. [Google Scholar] [CrossRef]
- Bodie, A.R.; Micciche, A.C.; Atungulu, G.G.; Rothrock, M.J.J.; Ricke, S.C. Current Trends of Rice Milling Byproducts for Agricultural Applications and Alternative Food Production Systems. Front. Sustain. Food Syst. 2019, 3, 47. [Google Scholar] [CrossRef]
- Van Keulen, H. Nitrogen Requirements of Rice with Special Reference to Java; CRIA: Bogor, Indonesia, 1977; Volume 30, pp. 1–67. [Google Scholar]
- Fageria, N.K.; Baligar, V.C. Lowland rice response to nitrogen fertilization. Commun. Soil Sci. Plant Anal. 2001, 32, 1405–1429. [Google Scholar] [CrossRef]
- Van Groenigen, K.J.; Van Kessel, C.; Hungate, B.A. Increased greenhouse-gas intensity of rice production under future atmospheric conditions. Nat. Clim. Change 2012, 3, 288–291. [Google Scholar] [CrossRef]
- Mohanty, S.R.; Bodelier, P.L.E.; Floris, V.; Conrad, R. Differential Effects of Nitrogenous Fertilizers on Methane-Consuming Microbes in Rice Field and Forest Soils. Appl. Environ. Microbiol. 2006, 72, 1346–1354. [Google Scholar] [CrossRef]
- Dubey, S.K. Microbial ecology of methane emission in rice agroecosystem: A review. Appl. Ecol. Envir. Res. 2005, 3, 1–27. [Google Scholar] [CrossRef]
- Schimel, J.P. Rice, microbes and methane. Nature 2000, 403, 375–377. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Lee, G.; Jin, Z.; Kim, B.; Chin, J.H.; Koh, H.-J. Development and application of indica–japonica SNP assays using the Fluidigm platform for rice genetic analysis and molecular breeding. Mol. Breed. 2020, 40, 39. [Google Scholar] [CrossRef]
- Ahmar, S.; Gill, R.A.; Jung, K.-H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [PubMed]
- Lenaerts, B.; Collard, B.C.; Demont, M. Review: Improving global food security through accelerated plant breeding. Plant Sci. 2019, 287, 110207. [Google Scholar] [CrossRef] [PubMed]
- Bush, W.S.; Moore, J.H. Chapter 11: Genome-Wide Association Studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef]
- Zargar, S.M.; Rai, V. Plant Omics and Crop Breeding; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Kim, S.; Mardis, E.; Tang, H. Genome Sequencing Technology and Algorithms; Artech House: Norwood, MA, USA, 2007. [Google Scholar]
- Kasahara, M.; Morishita, S. Large-Scale Genome Sequence Processing; Imperial College Press: London, UK, 2006. [Google Scholar]
- Fageria, N.K. The Role of Plant Roots in Crop Production; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Steudle, E. Water uptake by plant roots: An integration of views. Plant Soil 2000, 226, 45–56. [Google Scholar] [CrossRef]
- Kirkham, M.B.; Relations, P.W. Root Anatomy and Poiseuille’s Law for Water Flow in Roots in Principles of Soil and Plant Water Relations; Academic Press: Cambridge, MA, USA, 2014; pp. 243–266. [Google Scholar]
- Jungk, A. Root hairs and the acquisition of plant nutrients from soil. J. Plant Nut. Soil Sci. 2001, 164, 121–129. [Google Scholar] [CrossRef]
- Datta, S.; Kim, C.M.; Pernas, M.; Pires, N.D.; Proust, H.; Tam, T.; Vijayakumar, P.; Dolan, L. Root hairs: Development, growth and evolution at the plant-soil interface. Plant Soil 2011, 346, 1–14. [Google Scholar] [CrossRef]
- Lynch, J.P. Plant roots: Their growth, activity, and interaction with soils. Soil Sci. Soc. Am. J. 2007, 71, 636. [Google Scholar] [CrossRef]
- Green, M.S.; Etherington, J.R. Oxidation of Ferrous Iron by Rice (Oryza sativa L.) Roots: A Mechanism for Waterlogging Tolerance? J. Exp. Bot. 1977, 28, 678–690. [Google Scholar] [CrossRef]
- Moog, P.R.; Janiesch, P. Root Growth and Morphology of Carex Species as Influenced by Oxygen Deficiency. Funct. Ecol. 1990, 4, 201. [Google Scholar] [CrossRef]
- Borum, J.; Sand-Jensen, K.; Binzer, T.; Pedersen, O.; Greve, T.M. Oxygen movement in seagrasses. In Seagrasses: Biology, Ecologyand Conservation; Springer: Dordrecht, The Netherlands, 2007; pp. 255–270. [Google Scholar]
- Luxmoore, R.J.; Stolzy, L.H.; Letey, J. Oxygen Diffusion in the Soil-Plant System IV. Oxygen Concentration Profiles, Respiration Rates, and Radial Oxygen Losses Predicted for Rice Roots 1. Agron. J. 1970, 62, 329–332. [Google Scholar] [CrossRef]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Envir. 2003, 26, 17–36. [Google Scholar] [CrossRef]
- Yamauchi, T.; Shimamura, S.; Nakazono, M.; Mochizuki, T. Aerenchyma formation in crop species: A review. Field Crop. Res. 2013, 152, 8–16. [Google Scholar] [CrossRef]
- Evans, D.E. Aerenchyma formation. New Phytol. 2003, 161, 35–49. [Google Scholar] [CrossRef]
- Yamauchi, T.; Rajhi, I.; Nakazono, M. Lysigenous aerenchyma formation in maize root is confined to cortical cells by regulation of genes related to generation and scavenging of reactive oxygen species. Plant Signal. Behav. 2011, 6, 759–761. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.B.; Armstrong, W. Formation of Aerenchyma and the Processes of Plant Ventilation in Relation to Soil Flooding and Submergence. Plant Biol. 1999, 1, 274–287. [Google Scholar] [CrossRef]
- Jackson, M.B.; Fenning, T.M.; Jenkins, W. Aerenchyma (Gas-space) Formation in Adventitious Roots of Rice (Oryza sativa L.) is not Controlled by Ethylene or Small Partial Pressures of Oxygen. J. Exp. Bot. 1985, 36, 1566–1572. [Google Scholar] [CrossRef]
- Lai, W.-L.; Zhang, Y.; Chen, Z.-H. Radial oxygen loss, photosynthesis, and nutrient removal of 35 wetland plants. Ecol. Eng. 2011, 39, 24–30. [Google Scholar] [CrossRef]
- Smith, K.E.; Luna, T.O. Radial oxygen loss in wetland plants: Potential impacts on remediation of contaminated sediments. J. Envir. Eng. 2013, 139, 496–501. [Google Scholar] [CrossRef]
- Mitra, S.; Jain, M.; Kumar, S.; Bandyopadhyay, S.; Kalra, N. Effect of rice cultivars on methane emission. Agric. Ecosyst. Environ. 1999, 73, 177–183. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Papen, H.; Rennenberg, H. Impact of gas transport through rice cultivars on methane emission from rice paddy fields. Plant Cell Environ. 1997, 20, 1175–1183. [Google Scholar] [CrossRef]
- Zheng, H.; Fu, Z.; Zhong, J.; Long, W. Low methane emission in rice cultivars with high radial oxygen loss. Plant Soil 2018, 431, 119–128. [Google Scholar] [CrossRef]
- Visser, E.J.W.; Colmer, T.; Blom, C.W.P.M.; Voesenek, L.A.C.J. Changes in growth, porosity, and radial oxygen loss from adventitious roots of selected mono- and dicotyledonous wetland species with contrasting types of aerenchyma. Plant Cell Environ. 2000, 23, 1237–1245. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, M.; Wu, Y.; Wang, H.; Chen, Y.; Wu, W. Reducing CH4 and CO2 emissions from waterlogged paddy soil with biochar. J. Soils Sedim. 2011, 11, 930–939. [Google Scholar] [CrossRef]
- Liu, C.-W.; Wu, C.-Y. Evaluation of methane emissions from Taiwanese paddies. Sci. Total Environ. 2004, 333, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pedizzi, C.; Regueiro, L.; Rodriguez-Verde, I.; Lema, J.; Carballa, M. Effect of oxygen on the microbial activities of thermophilic anaerobic biomass. Bioresour. Technol. 2016, 211, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Tholen, A.; Pester, M.; Brune, A. Simultaneous methanogenesis and oxygen reduction by Methanobrevibacter cuticularis at low oxygen fluxes. FEMS Microbiol. Ecol. 2007, 62, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Kiener, A.; Leisinger, T. Oxygen Sensitivity of Methanogenic Bacteria. Syst. Appl. Microbiol. 1983, 4, 305–312. [Google Scholar] [CrossRef]
- Aschenbach, K.; Conrad, R.; Řeháková, K.; Doležal, J.; Janatková, K.; Angel, R. Methanogens at the top of the world: Occurrence and potential activity of methanogens in newly deglaciated soils in high-altitude cold deserts in the Western Himalayas. Front. Microbiol. 2013, 4, 359. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Han, S.J.; Yoon, E.K.; Lee, W.S. Evidence of an auxin signal pathway, microRNA167-ARF8-GH3, and its response to exogenous auxin in cultured rice cells. Nucleic Acids Res. 2006, 34, 1892–1899. [Google Scholar] [CrossRef] [PubMed]
- Velyvis, A.; Qin, J. LIM domain and its binding to target proteins. In Zinc Finger Proteins; Springer: Boston, MA, USA, 2007; pp. 99–105. [Google Scholar] [CrossRef]
- Yan, G.; Fan, X.; Tan, L.; Yin, C.; Li, T.; Liang, Y. Root silicon deposition and its resultant reduction of sodium bypass flow is modulated by OsLsi1 and OsLsi2 in rice. Plant Physiol. Biochem. 2020, 158, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Li, L.; Zhang, P.; Wang, D.; Yang, L.; Reza, B.M.; Lin, W. Lsi1 modulates the antioxidant capacity of rice and protects against ultraviolet-B radiation. Plant Sci. 2018, 278, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Cabral, P.D.; Herrera, M. Membrane-associated aquaporin-1 facilitates osmotically driven water flux across the basolateral membrane of the thick ascending limb. Am. J. Physiol. Physiol. 2012, 303, F621–F629. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Kaneko, T.; Sugimoto, G.; Sasano, S.; Panda, S.K.; Shibasaka, M.; Katsuhara, M. Mechanisms of Water Transport Mediated by PIP Aquaporins and Their Regulation Via Phosphorylation Events Under Salinity Stress in Barley Roots. Plant Cell Physiol. 2011, 52, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.-Y.; Su, Y.-H.; McGrath, S.P.; Zhao, F.-J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. High Silicon Accumulation in the Shoot is Required for Down-Regulating the Expression of Si Transporter Genes in Rice. Plant Cell Physiol. 2016, 57, 2510–2518. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Ponnamperuma, F.N. Effects of flooding on soils. In Flooding and Plant Growth; Academic Press: San Diego, CA, USA, 1984; Volume 10, pp. 9–45. [Google Scholar]
- Laanbroek, H. Bacterial cycling of minerals that affect plant growth in waterlogged soils: A review. Aquat. Bot. 1990, 38, 109–125. [Google Scholar] [CrossRef]
- Ejiri, M.; Shiono, K. Prevention of Radial Oxygen Loss Is Associated with Exodermal Suberin Along Adventitious Roots of Annual Wild Species of Echinochloa. Front. Plant Sci. 2019, 10, 254. [Google Scholar] [CrossRef]
- Armstrong, W. Aeration in Higher Plants. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 1979; Volume 7, pp. 225–332. [Google Scholar] [CrossRef]
- Colmer, T.D. Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland, paddy and deep-water rice (Oryza sativa L.). Ann. Bot. 2002, 91, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kotula, L.; Ranathunge, K.; Schreiber, L.; Steudle, E. Functional and chemical comparison of apoplastic barriers to radial oxygen loss in roots of rice (Oryza sativa L.) grown in aerated or deoxygenated solution. J. Exp. Bot. 2009, 60, 2155–2167. [Google Scholar] [CrossRef] [PubMed]
- Kotula, L.; Steudle, E. Measurements of oxygen permeability coefficients of rice (Oryza sativa L.) roots using a new perfusion technique. J. Exp. Bot. 2008, 60, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, N.; Li, T.; Yang, J.; Zhu, X.; Fang, C.; Li, S.; Si, H. Genome-wide identification and expression analysis of StTCP transcription factors of potato (Solanum tuberosum L.). Comput. Biol. Chem. 2018, 78, 53–63. [Google Scholar] [CrossRef]
- Zheng, K.; Ni, Z.; Qu, Y.; Cai, Y.; Yang, Z.; Sun, G.; Chen, Q. Genome-wide identification and expression analyses of TCP transcription factor genes in Gossypium barbadense. Sci. Rep. 2018, 8, 14526. [Google Scholar] [CrossRef] [PubMed]
- Danisman, S. TCP Transcription Factors at the Interface between Environmental Challenges and the Plant’s Growth Responses. Front. Plant Sci. 2016, 7, 1930. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Wang, Y.; Yang, A.; Zhang, W.-H. OsMYB2P-1, an R2R3 MYB Transcription Factor, Is Involved in the Regulation of Phosphate-Starvation Responses and Root Architecture in Rice. Plant Physiol. 2012, 159, 169–183. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, J.; Li, H.; Meng, D.; Li, R.; Dai, X.; Wang, S.; Liu, W.; Qu, H.; Xu, G. Maintenance of phosphate homeostasis and root development are coordinately regulated by MYB1, an R2R3-type MYB transcription factor in rice. J. Exp. Bot. 2017, 68, 3603–3615. [Google Scholar] [CrossRef]
- Na, J.-K.; Huh, S.-M.; Yoon, I.-S.; Byun, M.-O.; Lee, Y.-H.; Lee, K.-O.; Kim, D.-Y. Rice LIM protein OsPLIM2a is involved in rice seed and tiller development. Mol. Breed. 2014, 34, 569–581. [Google Scholar] [CrossRef]
- Ochiai, A.; Ogawa, K.; Fukuda, M.; Ohori, M.; Kanaoka, T.; Tanaka, T.; Taniguchi, M.; Sagehashi, Y. Rice Defensin OsAFP1 is a New Drug Candidate against Human Pathogenic Fungi. Sci. Rep. 2018, 8, 11434. [Google Scholar] [CrossRef]
- Sagehashi, Y.; Takaku, H.; Yatou, O. Partial peptides from rice defensin OsAFP1 exhibited antifungal activity against the rice blast pathogen Pyricularia oryzae. J. Pestic. Sci. 2017, 42, 172–175. [Google Scholar] [CrossRef]
- Luo, J.-S.; Huang, J.; Zeng, D.-L.; Peng, J.-S.; Zhang, G.-B.; Ma, H.-L.; Guan, Y.; Yi, H.-Y.; Fu, Y.-L.; Han, B.; et al. A defensin-like protein drives cadmium efflux and allocation in rice. Nat. Commun. 2018, 9, 645. [Google Scholar] [CrossRef]
- Ochiai, A.; Ogawa, K.; Fukuda, M.; Suzuki, M.; Ito, K.; Tanaka, T.; Sagehashi, Y.; Taniguchi, M. Crystal structure of rice defensin OsAFP1 and molecular insight into lipid-binding. J. Biosci. Bioeng. 2020, 130, 6–13. [Google Scholar] [CrossRef]
- Sampedro, J.; Lee, Y.; Carey, R.E.; DePamphilis, C.; Cosgrove, D.J. Use of genomic history to improve phylogeny and understanding of births and deaths in a gene family. Plant J. 2005, 44, 409–419. [Google Scholar] [CrossRef]
- Li, Y.; Darley, C.P.; Ongaro, V.; Fleming, A.; Schipper, O.; Baldauf, S.L.; McQueen-Mason, S.J. Plant Expansins Are a Complex Multigene Family with an Ancient Evolutionary Origin. Plant Physiol. 2002, 128, 854–864. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant expansins: Diversity and interactions with plant cell walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef]
- Che, J.; Yamaji, N.; Shen, R.F.; Ma, J.F. An Al-inducible expansin gene, OsEXPA10 is involved in root cell elongation of rice. Plant J. 2016, 88, 132–142. [Google Scholar] [CrossRef]
- Tan, J.; Wang, M.; Shi, Z.; Miao, X. OsEXPA10 mediates the balance between growth and resistance to biotic stress in rice. Plant Cell Rep. 2018, 37, 993–1002. [Google Scholar] [CrossRef]
- Jadamba, C.; Kang, K.; Paek, N.-C.; Lee, S.I.; Yoo, S.-C. Overexpression of Rice Expansin7 (Osexpa7) Confers Enhanced Tolerance to Salt Stress in Rice. Int. J. Mol. Sci. 2020, 21, 454. [Google Scholar] [CrossRef]
- ZhiMing, Y.; Bo, K.; XiaoWei, H.; ShaoLei, L.; YouHuang, B.; WoNa, D.; Ming, C.; Hyung-Taeg, C.; Ping, W. Root hair-specific expansins modulate root hair elongation in rice. Plant J. 2011, 66, 725–734. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhao, Y. The diverse biofunctions of LIM domain proteins: Determined by subcellular localization and protein-protein interaction. Biol. Cell 2007, 99, 489–502. [Google Scholar] [CrossRef]
- Fleck, A.T.; Nye, T.; Repenning, C.; Stahl, F.; Zahn, M.; Schenk, M.K. Silicon enhances suberization and lignification in roots of rice (Oryza sativa). J. Exp. Bot. 2010, 62, 2001–2011. [Google Scholar] [CrossRef]
- Trolldenier, G. Visualisation of oxidizing power of rice roots and of possible participation of bacteria in iron deposition. Z. Pflanz. Bodenkd. 1988, 151, 117–121. [Google Scholar] [CrossRef]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Bioph. Int. 2004, 11, 36–42. [Google Scholar]
- Thorpe, R.S. Multiple group principal component analysis and population differentiation. J. Zool. 1988, 216, 37–40. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2013. [Google Scholar]
- Cheon, K.-S.; Baek, J.; Cho, Y.-I.; Jeong, Y.-M.; Lee, Y.-Y.; Oh, J.; Won, Y.J.; Kang, D.-Y.; Oh, H.; Kim, S.L.; et al. Single Nucleotide Polymorphism (SNP) Discovery and Kompetitive Allele-Specific PCR (KASP) Marker Development with Korean Japonica Rice Varieties. Plant Breed. Biotechnol. 2018, 6, 391–403. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait (a) | QTL (b) | Chr (c) | Position (cM) (d) | Left Marker (e) | Right Marker (f) | LOD (g) | PVE (%) (h) | Add (i) | LeftCI (j) | RightCI (k) |
|---|---|---|---|---|---|---|---|---|---|---|
| ROL | qROL-2-1 | 2 | 127 | Os02_KJ02_047 | Os02_ad02011845 | 3.038 | 11.6082 | 2.5317 | 125.5 | 129.5 |
| RA | qRA-8-1 | 8 | 97 | Os08_cmb0824_7 | Os08_GW8--AG | 4.394 | 15.9468 | 0.7613 | 90.5 | 102 |
| No | Gene Name | MSU ID | Annotation | Biological Process | Molecular Function | Cellular Component |
|---|---|---|---|---|---|---|
| 1 | ARF8 TF | Os02g41800 | Auxin response factor, putative, expressed; ARF family protein | Multicellular organismal development; response to endogenous stimulus | RNA binding; sequence-specific DNA binding transcription factor activity | Nucleus |
| 2 | OsLsi1/ NIP2 | Os02g41860 | Aquaporin protein, putative | Transport; cell differentiation; response to endogenous stimulus | Transporter activity | Membrane |
| 3 | OsPSKR1 | Os02g41890 | Phytosulfokine receptor precursor | Response to stress; signal transduction; protein modification process | Kinase activity; receptor activity; binding; signal transducer activity | Cell |
| 4 | OsDEF7/CAL1/ OsAFP1 | Os02g41904 | Defensin-like DEFL family | Response to stress; defense response | Enzyme regulator activity | Cell wall; plasma membrane |
| 5 | OsWBC8 | Os02g41920 | White-brown complex homolog protein 8, ABC-2 type transporter domain containing protein | Transport; catabolic process; nucleic acid metabolic process | Hydrolase activity; transporter activity | Membrane |
| 6 | OsRR11/OsRRA8 | Os02g42060 | Response regulator receiver domain containing protein; A-TYPE response regulator 11 | Signal transduction; response to abiotic stimulus; response to auxin and cytokinin stimuli; two-component signal transduction system (phosphorelay) | Protein binding; signal transducer activity | Nucleus |
| 7 | OsWee1 | Os02g42110 | Wall-associated receptor kinase-like 22 precursor; cell cycle-associated protein kinase | Protein modification process; cell surface receptor linked signal transduction; calcium ion binding; | Kinase activity; protein binding; negative regulation of cell division; protein serine/threonine kinase activity | Plasma membrane |
| 8 | OsARF | Os02g42134 | ARF GTPase-activating domain-containing protein | Signal transduction; regulation of ARF GTPase activity | ARF GTPase activator activity; phospholipid binding; zinc ion binding; metal ion binding | Cytoplasm |
| 9 | OsWAK14 | Os02g42150 | Wall-associated kinase, receptor-like protein kinase; EGF-like calcium-binding domain containing protein. | Protein modification process | Kinase activity; calcium ion binding; polysaccharide binding | Plasma membrane |
| 10 | Lipase | Os02g42170 | Phospholipase, putative, expressed; lipase, class 3 domain containing protein | Response to stress; response to biotic stimulus; cellular process; lipid metabolic process | Hydrolase activity; triglyceride lipase activity | Cytoplasm; plastid |
| 11 | OsDLN61 | Os02g42200 | DLN REPRESSOR 61, B-block binding subunit of TFIIIC domain containing protein | Transcription initiation from RNA polymerase III promoter | 5S class rRNA transcription | |
| 12 | OsUGT | Os02g42280 | UDP-glucoronosyl/UDP-glucosyl transferase family protein | metabolic process | Transferase activity | Cell wall |
| 13 | OsE2/ RWD | Os02g42314 | Ubiquitin-conjugating enzyme/RWD-like domain containing protein | Fatty acid beta-oxidation; protein modification process; transport | Protein binding; ligase activity; acid-amino acid ligase activity | |
| 14 | OsTCP7 | Os02g42380 | TCP domain containing protein | Biosynthetic process | Protein binding; sequence-specific DNA binding transcription factor activity | Nucleus |
| 15 | OsLRR2 | Os02g42412 | Leucine-rich repeat 2, cysteine-containing subtype containing protein | RNA-dependent DNA biosynthetic process | RNA-directed DNA polymerase activity; RNA binding | |
| 16 | Os02g42520 | Dehydrogenase | Metabolic process | Binding; catalytic activity | plastid | |
| 17 | OsFTR | Os02g42570 | Ferredoxin-thioredoxin reductase, variable chain | Generation of precursor metabolites and energy; photosynthesis | Catalytic activity | Plastid |
| 18 | OsBRO1 | Os02g42580 | Vacuolar protein-sorting protein bro1 | - | - | Cytosol |
| 19 | OsAP2 TF | Os02g42585 | AP2 domain containing protein | Biosynthetic process; nucleic acid metabolic process | DNA binding; sequence-specific DNA binding transcription factor activity | Nucleus; cytoplasm |
| 20 | OsWD-40 | Os02g42590 | WD-40 repeat family protein, putative, expressed | Signal transduction | Signal transducer activity | Intracellular; plasma membrane |
| 21 | Os02g42600 | Double-stranded RNA binding motif containing protein | Response to stress; signal transduction; response to abiotic stimulus | Hydrolase activity; RNA binding; protein binding; | Nucleus; intracellular | |
| 22 | OsEXPA | Os02g42650 | Expansin precursor, putative, expressed | Anatomical structure morphogenesis; cell growth; cellular process | - | - |
| 23 | OsMLD/ DES1 | Os02g42660 | Degenerative spermatocyte homolog 1, lipid desaturase/migration-inducing gene 15 protein/sphingolipid delta 4 desaturase protein | Sphingolipid biosynthetic process; plant-type cell wall modification; fatty acid biosynthetic process; oxidation-reduction process | Sphingolipid delta-4 desaturase activity; oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen | Integral component of membrane |
| 24 | OsDUF1771 | Os02g42670 | DUF1771 domain containing protein | Regulation of gene expression, epigenetic | - | - |
| 25 | Os02g42690 | Zinc finger, C3HC4 type domain containing protein | Transport; protein modification process; protein metabolic process | Catalytic activity; binding | Membrane; endoplasmic reticulum; | |
| 26 | OsTRX | Os02g42700 | Thioredoxin | Response to stress; metabolic process | Enzyme regulator activity | Membrane; plastid; thylakoid |
| 27 | Oscb5 | Os02g42740 | Cytochrome b5-like Heme/Steroid binding domain containing protein | Binding | Membrane; plastid; thylakoid; nucleus | |
| 28 | Lectin | Os02g42780 | Lectin receptor-type protein kinase | Metabolic process; cellular process | Kinase activity | Plasma membrane |
| 29 | Os02g42790 | Short-chain dehydrogenase/reductase | Metabolic process; response to abiotic stimulus; | Catalytic activity; response to stress; oxidoreductase activity | Cytosol; plasma membrane | |
| 30 | Os02g42810 | Oxidoreductase, short-chain dehydrogenase/reductase family domain containing protein | Metabolic process | Catalytic activity; binding | Cell wall | |
| 31 | OsPLIM2a | Os02g42820 | LIM domain protein, putative actin-binding protein, and transcription factor | Cellular component organization; cellular process | Protein binding | Cytoskeleton |
| 32 | OsMYB21 | Os02g42850 | MYB family transcription factor | Biosynthetic process; nucleobase, nucleoside, nucleotide, and nucleic acid metabolic process | Sequence-specific DNA binding transcription factor activity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duyen, D.V.; Kwon, Y.; Kabange, N.R.; Lee, J.-Y.; Lee, S.-M.; Kang, J.-W.; Park, H.; Cha, J.-K.; Cho, J.-H.; Shin, D.; et al. Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L.) under Iron (II) Sulfide. Plants 2022, 11, 788. https://doi.org/10.3390/plants11060788
Duyen DV, Kwon Y, Kabange NR, Lee J-Y, Lee S-M, Kang J-W, Park H, Cha J-K, Cho J-H, Shin D, et al. Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L.) under Iron (II) Sulfide. Plants. 2022; 11(6):788. https://doi.org/10.3390/plants11060788
Chicago/Turabian StyleDuyen, Dang Van, Youngho Kwon, Nkulu Rolly Kabange, Ji-Yoon Lee, So-Myeong Lee, Ju-Won Kang, Hyeonjin Park, Jin-Kyung Cha, Jun-Hyeon Cho, Dongjin Shin, and et al. 2022. "Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L.) under Iron (II) Sulfide" Plants 11, no. 6: 788. https://doi.org/10.3390/plants11060788
APA StyleDuyen, D. V., Kwon, Y., Kabange, N. R., Lee, J. -Y., Lee, S. -M., Kang, J. -W., Park, H., Cha, J. -K., Cho, J. -H., Shin, D., & Lee, J. -H. (2022). Novel QTL Associated with Aerenchyma-Mediated Radial Oxygen Loss (ROL) in Rice (Oryza sativa L.) under Iron (II) Sulfide. Plants, 11(6), 788. https://doi.org/10.3390/plants11060788