
Improved Medicago sativa Nodulation under Stress Assisted by Variovorax sp. Endophytes

 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Characterization of Strains
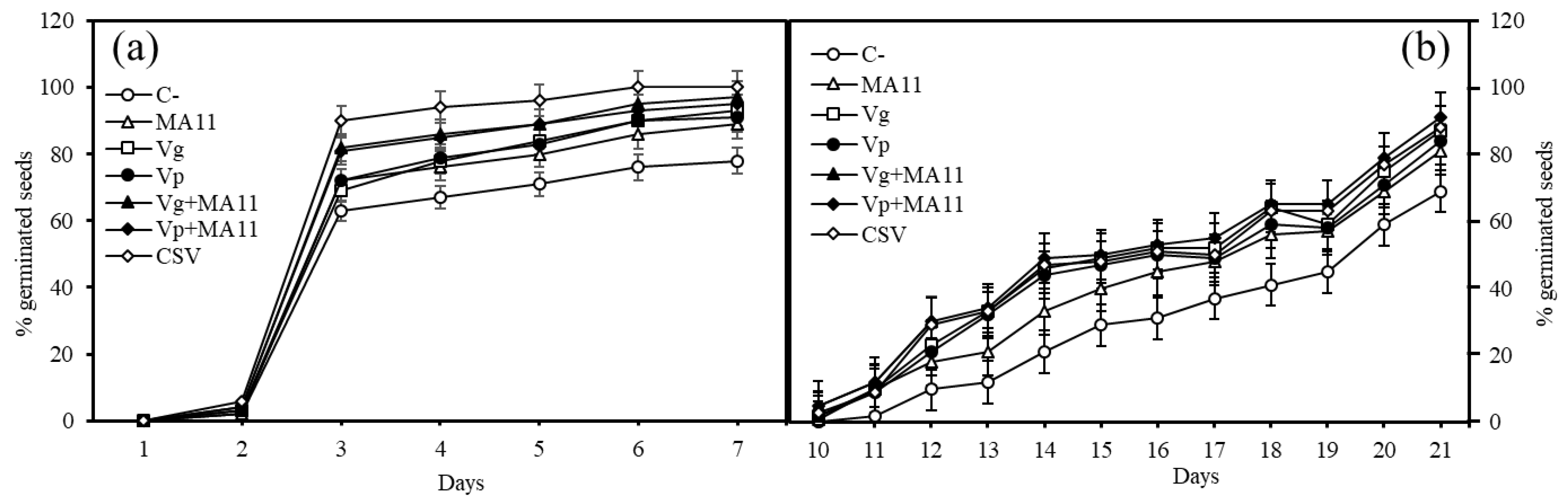
2.2. Inoculation Enhanced M. sativa Seeds Germination
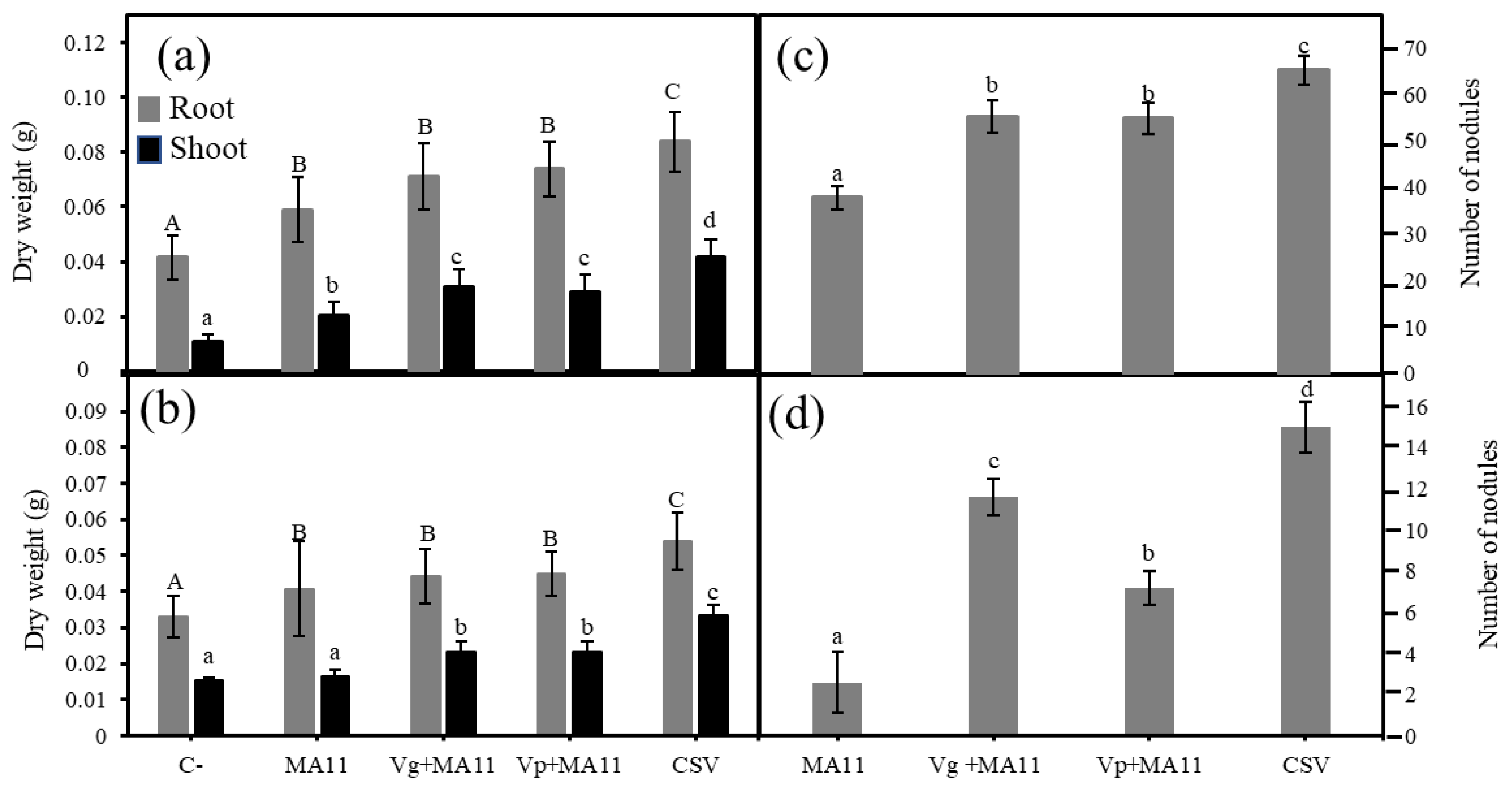
2.3. Variovorax Increased M. sativa Growth and Nodulation In Vitro
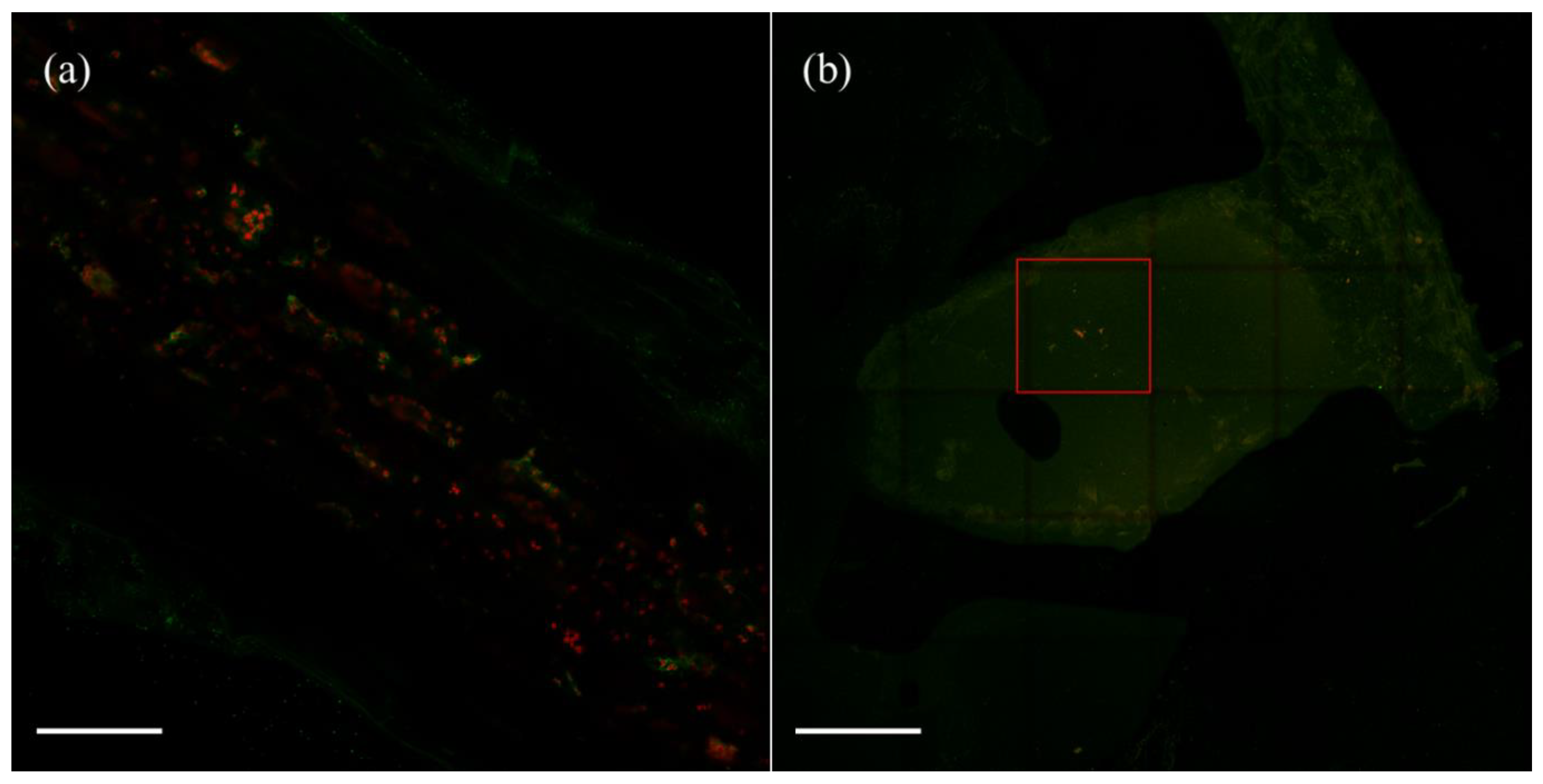
2.4. Variovorax Behaves as Nodule Endophytes
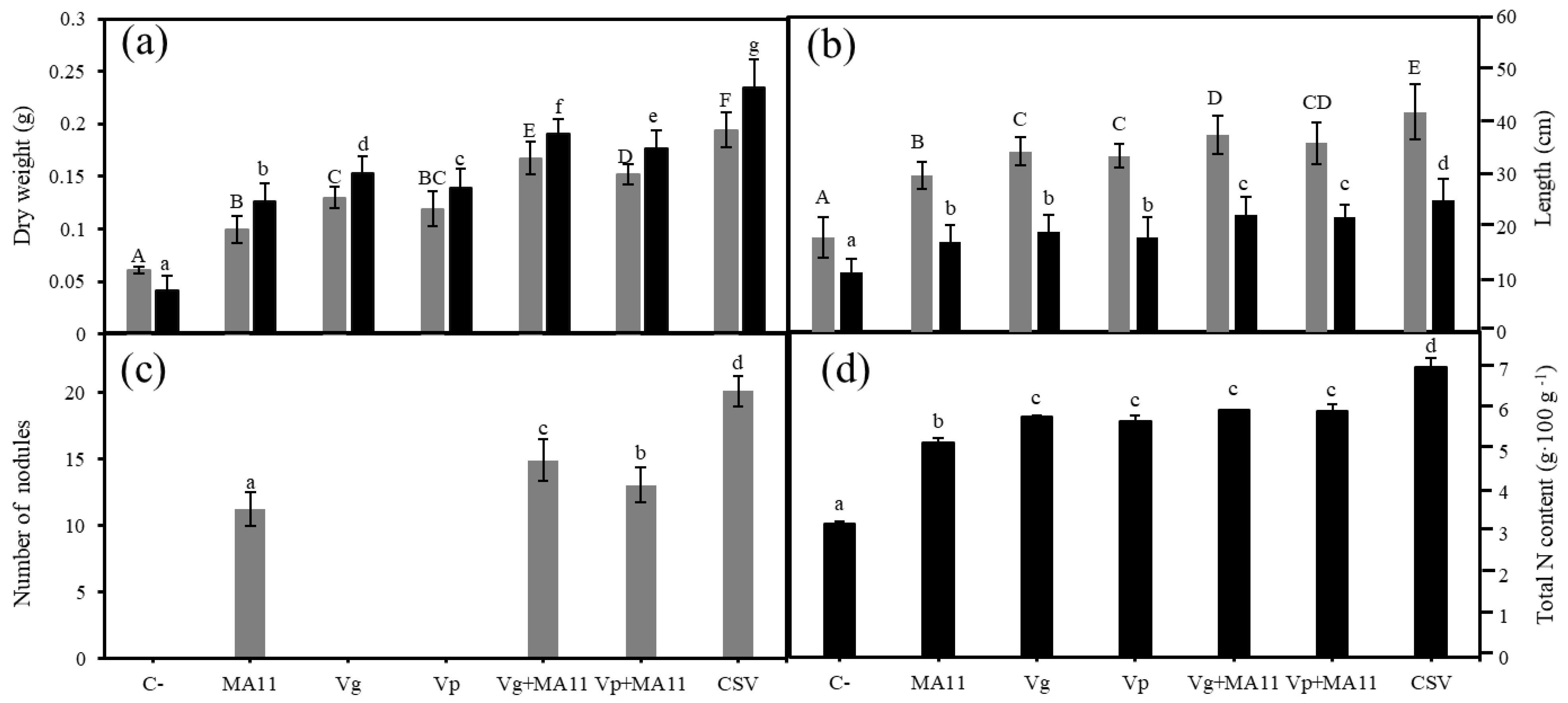
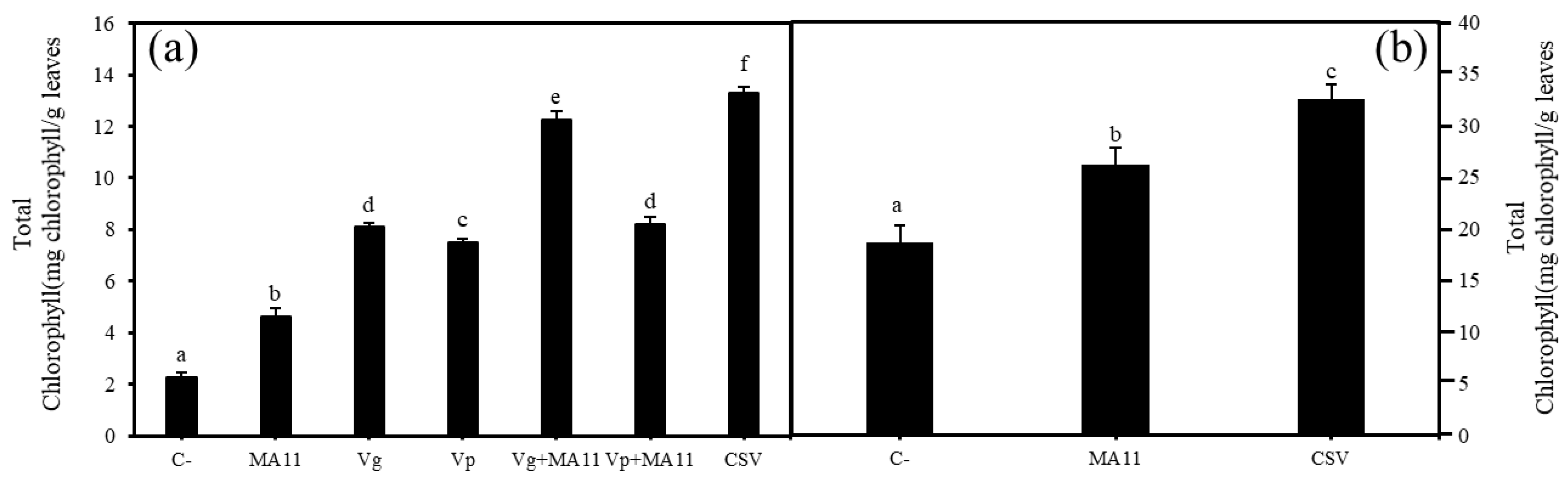
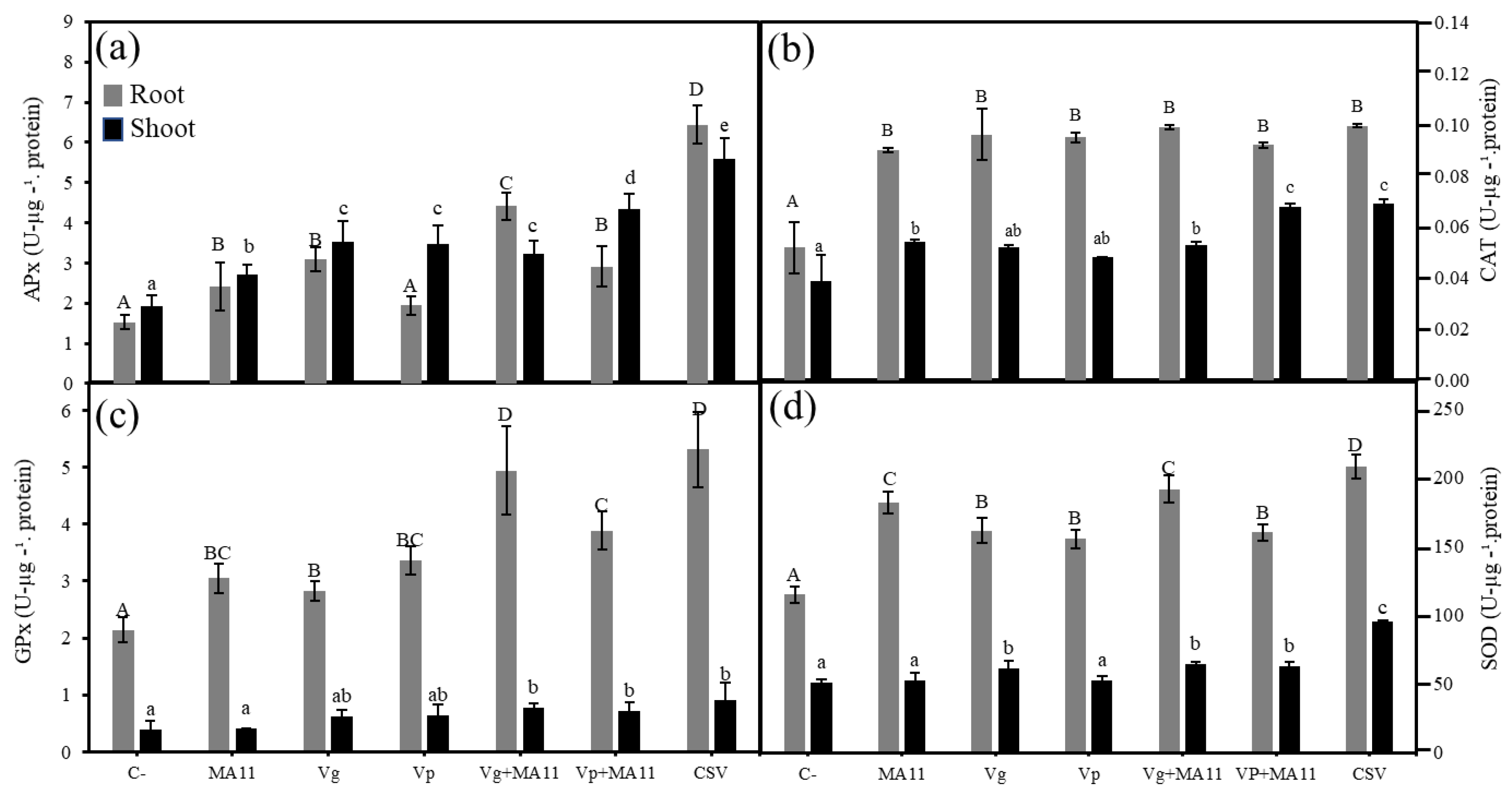
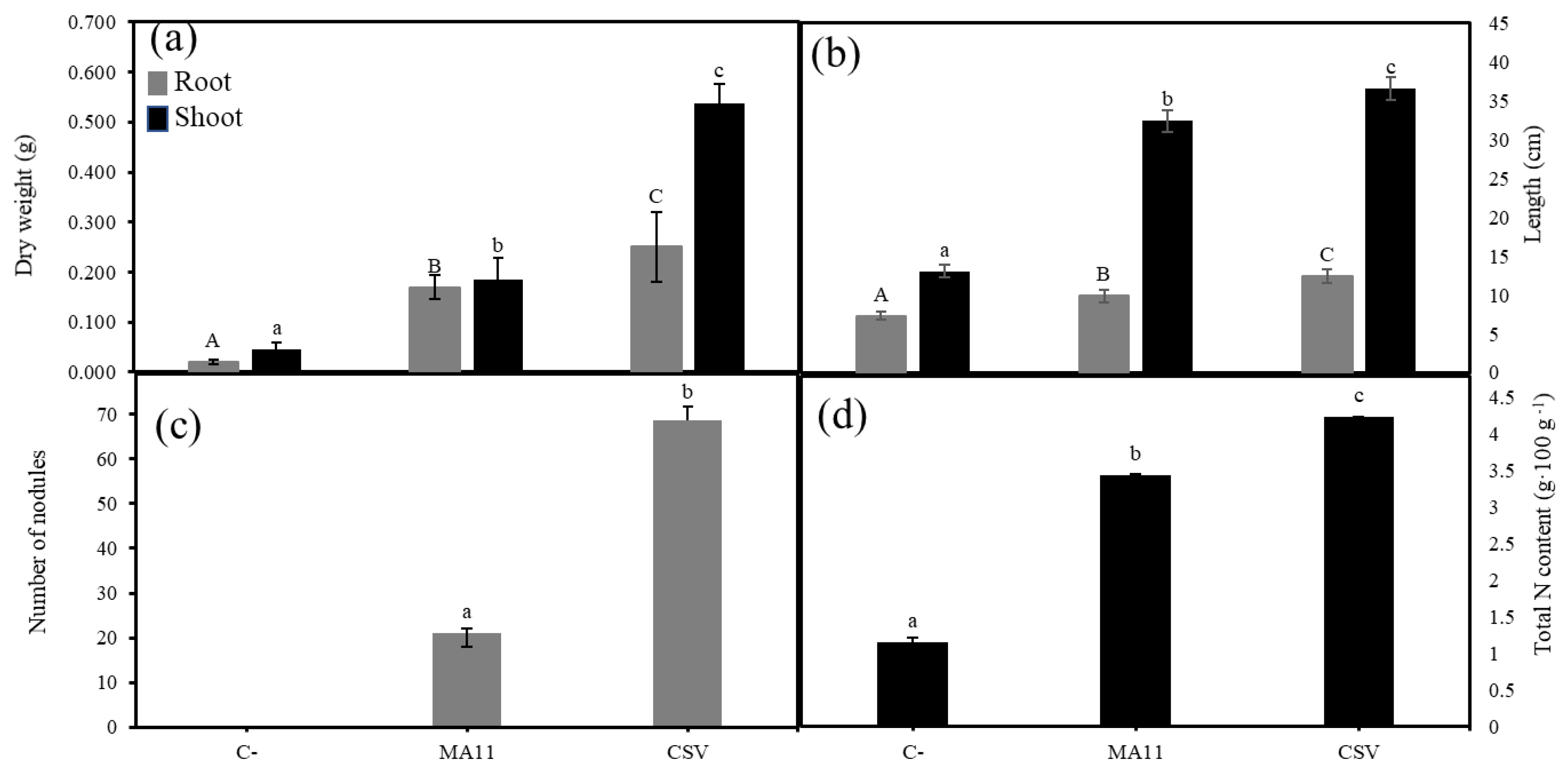
2.5. Variovorax Increased M. sativa Growth and Nodulation in Estuarine Soils with Nutrient Poverty
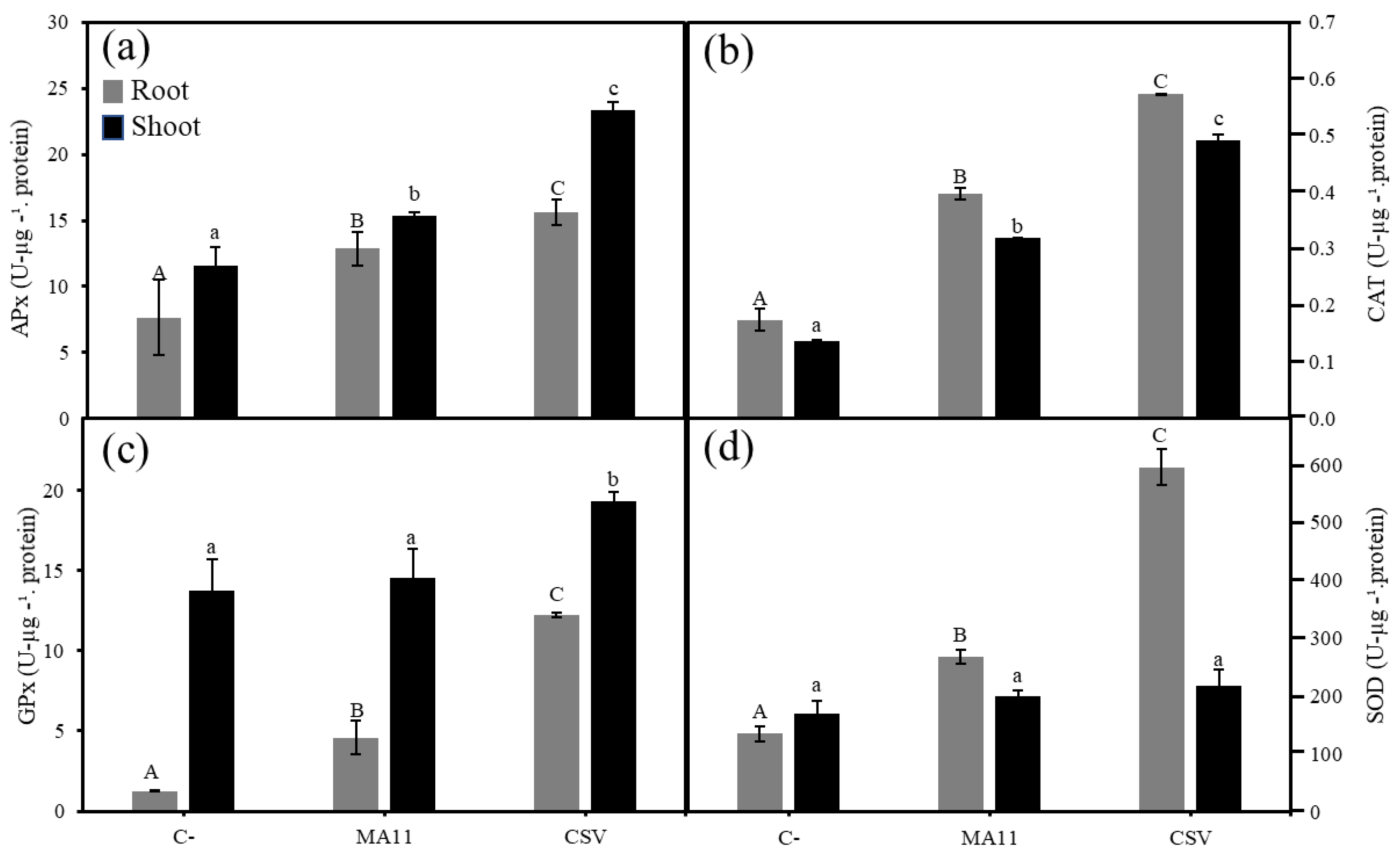
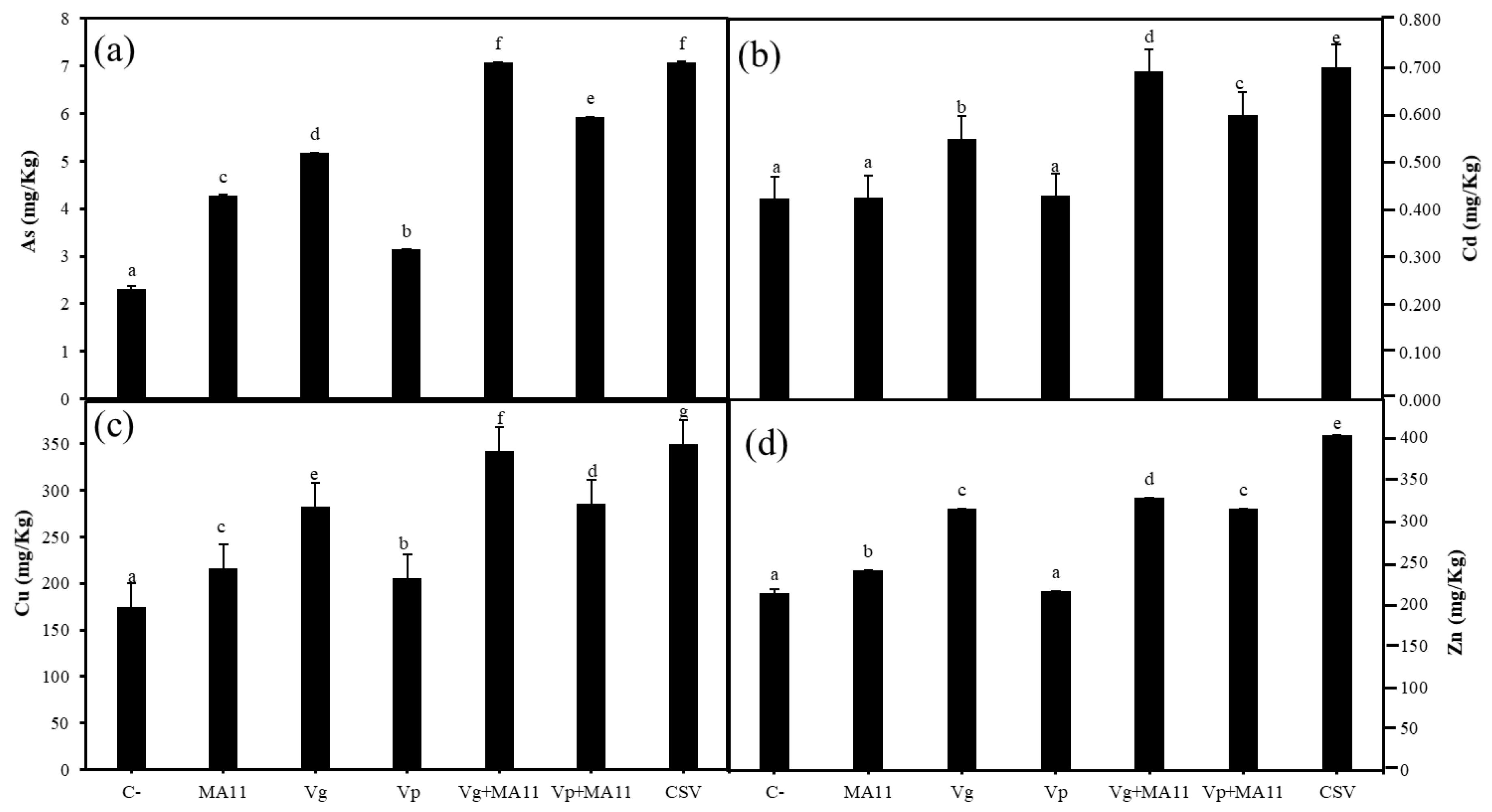
2.6. Variovorax Increased M. sativa Growth and Nodulation in Estuarine Soils Contaminated with Metals
2.7. Variovorax Increased Metal Accumulation on M. sativa Roots
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Metal Tolerance
4.2. Determination of PGP Properties
4.3. Screening for Bacterial Enzyme Activities
4.4. PGP Properties and Enzyme Activities in Presence of Metals
4.5. Labelled of Variovorax with Fluorescence and Microscopy
4.6. Medicago sativa Seeds Germination and Growth on Plates
4.7. Collection and Characterization of Soil
4.8. Pot Inoculation in Greenhouse Conditions
4.9. Chlorophyll Content
4.10. Antioxidant Enzymes Determination
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO; ITPS. Status of the World’s Soil Resources (SWSR)—Main Report; FAO and Intergovernmental Technical Panel on Soils: Roma, Italy, 2015. [Google Scholar]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Poole, P.; Ramachandran, V.; Terpolilli, V. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Olivares, J.; Bedmar, E.J.; Sanjuán, J. Biological nitrogen fixation in the context of global change. Mol. Plant Microb. Interac. 2013, 26, 486–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hidalgo, P.; Hirsch, A.M. The nodule microbiome: N2-fixing rhizobia do not live alone. Phytobiomes J. 2017, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, S.E.; De Beuf, K.; Vekeman, B. A large diversity of non-rhizobial endophytes found in legume root nodules in Flanders (Belgium). Soil Biol. Biochem. 2015, 83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, G.; Patel, M.H.; Joshi, S.J. Isolation and characterization of nodule-associated Exiguobacterium sp. from the root nodules of fenugreek (Trigonella foenu-graecum) and their possible role in plant growth promotion. Int. J. Microbiol. 2012, 2012, 693982. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, E.; Martínez-Hidalgo, P.; Carro, L.; Alonso, P.; Peix, A.; Trujillo, M.E.; Martínez-Molina, E. Nodular endophytes: An untapped diversity. In Beneficial Plant—Microbial Interactions: Ecology and Applications; González, B.R.M., González-López, J., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 214–236. [Google Scholar]
- Aserse, A.A.; Räsänen, L.A.; Aseffa, F.; Hailemariam, A.; Lindström, K. Diversity of sporadic symbionts and nonsymbiotic endophytic bacteria isolated from nodules of woody, shrub, and food legumes in Ethiopia. Appl. Microbiol. Biotechnol. 2013, 97, 10117–10134. [Google Scholar] [CrossRef]
- Hoque, M.S.; Broadhurst, L.M.; Thrall, P.H. Genetic characterization of root-nodule bacteria associated with Acacia salicina and A. stenophylla (Mimosaceae) across south-eastern Australia. Int. J. Syst. Evol. Microbiol. 2011, 61, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Bessadok, K.; Navarro-Torre, S.; Pajuelo, E.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Caviedes, M.Á.; Fterich, A.; Mars, M.; Rodríguez-Llorente, I.D. The ACC-deaminase producing bacterium Variovorax sp. CT7.15 as a tool for improving Calicotome villosa nodulation and growth in arid regions of Tunisia. Microorganisms 2020, 8, 541. [Google Scholar] [CrossRef] [Green Version]
- Satola, B.; Wübbeler, J.H.; Steinbüchel, A. Metabolic characteristics of the species Variovorax paradoxus. Appl. Microbiol. Biotechnol. 2013, 97, 541–560. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P.; Busse, H.J.; McInroy, J.A.; Glaeser, S.P. Variovorax gossypii sp. nov., isolated from Gossypium hirsutum. Int. J. Syst. Evol. Microbiol. 2015, 65, 4335–4340. [Google Scholar] [CrossRef] [PubMed]
- Pajuelo, E.; Rodríguez-Llorente, I.D.; Dary, M.; Palomares, A.J. Toxic effects of arsenic on Sinorhizobium–Medicago sativa symbiotic interaction. Environ. Pollut. 2008, 154, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.D.; Köhler, C. Auxin: A molecular trigger of seed development. Genes Dev. 2018, 32, 479–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under stress: Involvement of auxin and cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Zhang, H.; Yan, H.; Qiu, L.; Baskin, C.C. Mobilization and role of starch, protein, and fat reserves during seed germination of six wild grassland species. Front. Plant Sci. 2018, 9, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walitang, D.I.; Kim, K.; Madhaiyan, M.; Kim, Y.K.; Kang, Y.; Sa, T. Characterizing endophytic competence and plant growth promotion of bacterial endophytes inhabiting the seed endosphere of rice. BMC Microbiol. 2017, 17, 209. [Google Scholar] [CrossRef]
- Caba, J.M.; Recalde, L.; Ligero, F. Nitrate-induced ethylene biosynthesis and the control of nodulation in alfalfa. Plant Cell Environ. 1998, 21, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Oldroyd, G.E.; Engstrom, E.M.; Long, S.R. Ethylene inhibits the Nod factor signal transduction pathway of Medicago truncatula. Plant Cell 2001, 13, 1835–1849. [Google Scholar] [CrossRef] [Green Version]
- Peters, N.K.; Crist-Estes, D.K. Nodule formation is stimulated by the ethylene inhibitor aminoethoxyvinylglycine. Plant Physiol. 1989, 91, 690–693. [Google Scholar] [CrossRef]
- Nukui, N.; Ezura, H.; Yuhashi, K.-I.; Yasuta, T.; Minamisawa, K. Effects of ethylene precursor and inhibitors for ethylene biosynthesis and perception on nodulation in Lotus japonicus and Macroptilium atropurpureum. Plant Cell Physiol. 2000, 41, 893–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar] [CrossRef]
- Selim, S.M.; Zayed, M.S. Role of biofertilizers in sustainable agriculture under abiotic stresses. In Microorganisms for Green Revolution. Microorganisms for Sustainability; Panpatte, D., Jhala, Y., Vyas, R., Shelat, H., Eds.; Springer: Singapore, 2017; Volume 6, pp. 281–301. [Google Scholar] [CrossRef]
- Kang, B.G.; Kim, W.T.; Yun, H.S.; Chang, S.C. Use of plant growth-promoting rhizobacteria to control stress responses of plant roots. Plant Biotechnol. Rep. 2010, 4, 179–183. [Google Scholar] [CrossRef]
- Turan, M.; Kıtır, N.; Alkaya, Ü.; Günes, A.; Tüfenkçi, S.; Yıldırım, E.; Nikerel, E. Making soil more accessible to plants: The case of plant growth promoting rhizobacteria. In Plant Growth; Rigobelo, E.C., Ed.; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Benito, P.; Alonso-Vega, P.; Aguado, C.; Luján, R.; Anzai, Y.; Hirsch, A.M.; Trujillo, M.E. Monitoring the colonization and infection of legume nodules by Micromonospora in co-inoculation experiments with rhizobia. Sci. Rep. 2017, 7, 11051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinel, F.C. Ethylene, a hormone at the center-stage of nodulation. Front. Plant Sci. 2015, 6, 1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D.; Denton, M.; Zhou, Y. Mechanisms in plant growth-promoting rhizobacteria that enhance legume-rhizobial symbioses. J. Appl. Microbiol. 2020, 129, 1133–1156. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Alori, E.T.; Babalola, O.O. Microbial inoculants for improving crop quality and human health in Africa. Front. Microbiol. 2018, 9, 2213. [Google Scholar] [CrossRef] [Green Version]
- Jha, C.K.; Saraf, M. Plant growth promoting rhizobacteria (PGPR): A review. E3 J. Agric. Res. Dev. 2015, 5, 108–119. [Google Scholar]
- Honma, M.; Shimomura, T. Metabolism of 1-aminocyclopropane-1-carboxylic acid. Agric. Biol. Chem. 1978, 43, 1825–1831. [Google Scholar]
- Redondo-Gómez, S.; Andrades-Moreno, L.; Mateos-Naranjo, E.; Parra, R.; Valera-Burgos, J.; Arcoa, R. Synergic effect of salinity and zinc stress on growth and photosynthetic responses of the cordgrass, Spartina densiflora. J. Exp. Bot. 2011, 62, 5521–5530. [Google Scholar] [CrossRef] [PubMed]
- Mesnoua, M.; Mateos-Naranjo, E.; Barcia-Piedras, J.M.; Perez-Romero, J.A.; Lotmani, B.; Redondo-Gomez, S. Physiological and biochemical mechanisms preventing Cd-toxicity in the hyperaccumulator Atriplex halimus L. Plant Physiol. Biochem. 2016, 106, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Noori, F.; Etesami, H.; Najafi Zarini, H.; Khoshkholgh-Sima, N.A.; Hosseini Salekdeh, G.; Alishahi, F. Mining alfalfa (Medicago sativa L.) nodules for salinity tolerant non-rhizobial bacteria to improve growth of alfalfa under salinity stress. Ecotoxicol. Environ. Saf. 2018, 162, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, H.; Leng, J.; Niu, H.; Chen, X.; Liu, D.; Chen, Y.; Gao, N.; Ying, H. Isolation and characterization of plant growth-promoting rhizobacteria and their effects on the growth of Medicago sativa L. under salinity conditions. Antonie Leeuwenhoek 2020, 113, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Delgadillo, J.; Lafuente, A.; Doukkali, B.; Redondo-Gómez, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Improving legume nodulation and Cu rhizostabilization using a genetically modified rhizobia. Environ. Technol. 2015, 36, 1237–1245. [Google Scholar] [CrossRef]
- Dary, M.; Chamber-Perez, M.A.; Palomares, A.J.; Pajuelo, E. In situ phytostabilisation of heavy metal polluted soils using Lupinus luteus inoculated with metal resistant plant growth promoting rhizobacteria. J. Hazard. Mater. 2010, 177, 323–330. [Google Scholar] [CrossRef]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.W.; Fallmann, K.; Puschenreiter, M. The role of plant-associated bacteria in the mobilization and phytoextraction of trace elements in contaminated soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Méndez, M.O.; Maier, R.M. Phytoestabilisation of mine tailings in arid and semiarid environments: An emerging remediation technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F. Beneficial biofilms for land rehabilitation and fertilization. FEMS Microbiol. Lett. 2020, 367, fnaa184. [Google Scholar] [CrossRef]
- Vincent, J.M. A manual for the practical study of root nodule bacteria. IBM Handb. 1970, 45, 440. [Google Scholar]
- Döebereiner, J. Isolation and identification of aerobic nitrogen-fixing bacteria from soil and plants. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 134–141. [Google Scholar]
- Buendía-Clavería, A.M.; Chamber, M.; Ruiz-Sainz, J.E. A comparative study of the physiological characteristics, plasmid content and symbiotic properties of different Rhizobium fredii strains. Syst. Appl. Microbiol. 1989, 12, 203–209. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Endophytic cultivable bacteria of the metal bioaccumulator Spartina maritima improve plant growth but not metal uptake in polluted marshes soils. Front. Microbiol. 2015, 6, 1450. [Google Scholar] [CrossRef]
- Dworkin, M.; Foster, J.W. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Scouting contaminated estuaries: Heavy metal resistant and plant growth promoting rhizobacteria in the native metal rhizoaccumulator Spartina maritima. Mar. Pollut. Bull. 2015, 90, 150–159. [Google Scholar] [CrossRef]
- Elbeltagy, A.; Nishioka, K.; Suzuki, H.; Sato, T.; Sato, Y.I.; Morisaki, H.; Mitsui, H.; Minamisawa, K. Isolation and characterization of endophytic bacteria from wild and traditionally cultivated rice varieties. Soil Sci. Plant Nutr. 2000, 46, 617–629. [Google Scholar] [CrossRef]
- Harley, J.P.; Prescott, L.M. Laboratory Exercises in Microbiology, 5th ed.; McGraw-Hill: New York, NY, USA, 2002; p. 320. [Google Scholar]
- Lagendijk, E.L.; Validov, S.; Lamers, G.E.M.; de Weert, S.; Bloemberg, G.V. Genetic tools for tagging Gram-negative bacteria with mCherry for visualization in vitro and in natural habitats, biofilm and pathogenicity studies. FEMS Microbiol. Lett. 2010, 305, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Finan, T.M.; Kunkel, B.; De Vos, G.F.; Signer, E.R. Second symbiotic megaplasmid in Rhizobium meliloti carrying exopolysaccharide and thiamine synthesis genes. J. Bacteriol. 1986, 167, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Torre, S.; Rodríguez-Llorente, I.D.; Doukkali, B.; Caviedes, M.A.; Pajuelo, E. Competition for alfalfa nodulation under metal stress by the metal-tolerant strain Ochrobactrum cytisi Azn6.2. Ann. Appl. Biol. 2019, 175, 184–192. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Mesa, J.; Pajuelo, E.; Perez-Martin, A.; Caviedes, M.A.; Rodríguez-Llorente, I.D. Deciphering the role of plant growth promoting rhizobacteria in the tolerance of the invasive cordgrass Spartina densiflora to physicochemical properties of marshes soils. Plant Soil. 2015, 394, 45–55. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Directions for making mechanical analyses of soil by the hydrometer method. Soil Sci. 1936, 42, 225–229. [Google Scholar] [CrossRef]
- Carrasco, J.A.; Armario, P.; Pajuelo, E.; Burgos, A.; Caviedes, M.A.; López, R.; Chamber, M.A.; Palomares, A.J. Isolation and characterisation of symbiotically effective Rhizobium resistant to arsenic and heavy metals after the toxic spill at the Aznalcóllar pyrite mine. Soil Biol. Biochem. 2005, 37, 1131–1140. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 2011, 57, 1332–1334. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [Green Version]
- Duarte, B.; Goessling, J.W.; Marques, J.C.; Caçador, I. Ecophysiological constraints of Aster tripolium under extreme thermal events impacts: Merging biophysical, biochemical and genetic insights. Plant Physiol. Biochem. 2015, 97, 217–228. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | As (mM) | Cd (mM) | Cu (mM) | Zn (mM) |
|---|---|---|---|---|
| MA11 | 10 | 1.5 | 3 | 8 |
| Vg | 9 | 5.9 | 10.6 | 10.7 |
| Vp | 8.9 | 4 | 10.3 | 12 |
| PGP Properties | Without Metal | With Metal | ||||
|---|---|---|---|---|---|---|
| MA11 | Vg | Vp | MA11 | Vg | Vp | |
| Phosphate solubilizing | 1.70 | - | - | 1.10 | 1.20 | 1.00 |
| Siderophores production | 1.41 | 5.00 | 4.00 | n. d | n. d | n. d |
| IAA production | 1.76 | 4.26 | 2.11 | 1.07 | 2.28 | 1.51 |
| Biofilm formation | 0.14 | - | 0.56 | 0.27 | 0.10 | 0.18 |
| ACC deaminase activity | 1.03 | 2.66 | 2.11 | 9.08 | 10.03 | 8.55 |
| Enzymatic activities | ||||||
| DNAse | - | + | - | - | - | - |
| Amylase | + | + | + | + | - | - |
| Cellulase | - | - | - | - | - | - |
| Lipase | - | + | - | - | - | - |
| Pectinas | - | + | - | - | - | - |
| Protease | - | + | - | - | + | - |
| Chitinase | - | - | - | - | - | - |
| Physicochemical Properties | ||||||||
| Location | Texture (%) * | Organic Material (%) | Conductivity (μS·cm−1) | pH | ||||
| High marsh | 70/15/15 | 0.9 ± 0.05 | 12.8 ± 0.6 | 6.9 ± 0.1 | ||||
| Low marsh | 28.51/44.99/26.49 | 3.66 ± 0.1 | 15.5 ± 0.2 | 6.8 ± 0.05 | ||||
| Metal/Loids Concentration (mg·kg−1) | ||||||||
| Location | As | Cd | Cu | Zn | Mg | Na | Fe | p |
| High marsh | 26.7 ± 3.6 | 0.36 ± 0.1 | 311.1 ± 14.3 | 375.0 ± 9.2 | 0.777 ± 0.02 | 0.359 ± 0.006 | 9125.78 ± 12.4 | 0.054 ± 0.012 |
| Low marsh | 565.9 ± 10.9 | 2.3 ± 0.0 | 1200.3 ± 36.7 | 2425.7 ± 4.0 | 1.047± 0.064 | 1.707 ± 0.04 | 84,988.68 ± 6.1 | 0.412 ± 0.007 |
| As (mg·kg−1) | Cd (mg·kg−1) | Cu (mg·kg−1) | Zn (mg·kg−1) | |
|---|---|---|---|---|
| Shoot | ||||
| C– | 7.23 ± 0.00 a | 0.12 ± 0.00 a | 36.08 ± 0.02 a | 57.61 ± 0.07 a |
| MA11 | 8.97 ± 0.02 b | 0.27 ± 0.00 b | 48.51 ± 0.00 b | 90.92 ± 0.06 b |
| CSV | 6.98 ± 0.07 c | 0.18 ± 0.01 c | 50.83 ± 0.02 c | 68.39 ± 0.72 c |
| Root | ||||
| C– | 17.45 ± 0.41 a | 0.30 ± 0.01 a | 78.95 ± 0.07 a | 214.82 ± 0.49 a |
| MA11 | 51.82 ± 1.31 b | 1.23 ± 0.07 b | 218.30 ± 0.51 b | 401.45 ± 0.07 b |
| CSV | 39.43 ± 0.70 c | 1.51 ± 0.01 c | 326.62 ± 0.72 c | 430.07 ± 0.01 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Duarte, N.J.; Pérez-Pérez, J.; Navarro-Torre, S.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Improved Medicago sativa Nodulation under Stress Assisted by Variovorax sp. Endophytes. Plants 2022, 11, 1091. https://doi.org/10.3390/plants11081091
Flores-Duarte NJ, Pérez-Pérez J, Navarro-Torre S, Mateos-Naranjo E, Redondo-Gómez S, Pajuelo E, Rodríguez-Llorente ID. Improved Medicago sativa Nodulation under Stress Assisted by Variovorax sp. Endophytes. Plants. 2022; 11(8):1091. https://doi.org/10.3390/plants11081091
Chicago/Turabian StyleFlores-Duarte, Noris J., Julia Pérez-Pérez, Salvadora Navarro-Torre, Enrique Mateos-Naranjo, Susana Redondo-Gómez, Eloísa Pajuelo, and Ignacio D. Rodríguez-Llorente. 2022. "Improved Medicago sativa Nodulation under Stress Assisted by Variovorax sp. Endophytes" Plants 11, no. 8: 1091. https://doi.org/10.3390/plants11081091
APA StyleFlores-Duarte, N. J., Pérez-Pérez, J., Navarro-Torre, S., Mateos-Naranjo, E., Redondo-Gómez, S., Pajuelo, E., & Rodríguez-Llorente, I. D. (2022). Improved Medicago sativa Nodulation under Stress Assisted by Variovorax sp. Endophytes. Plants, 11(8), 1091. https://doi.org/10.3390/plants11081091