
Choosing the Right Path for the Successful Storage of Seeds



{kind=link}
Abstract
:1. Introduction
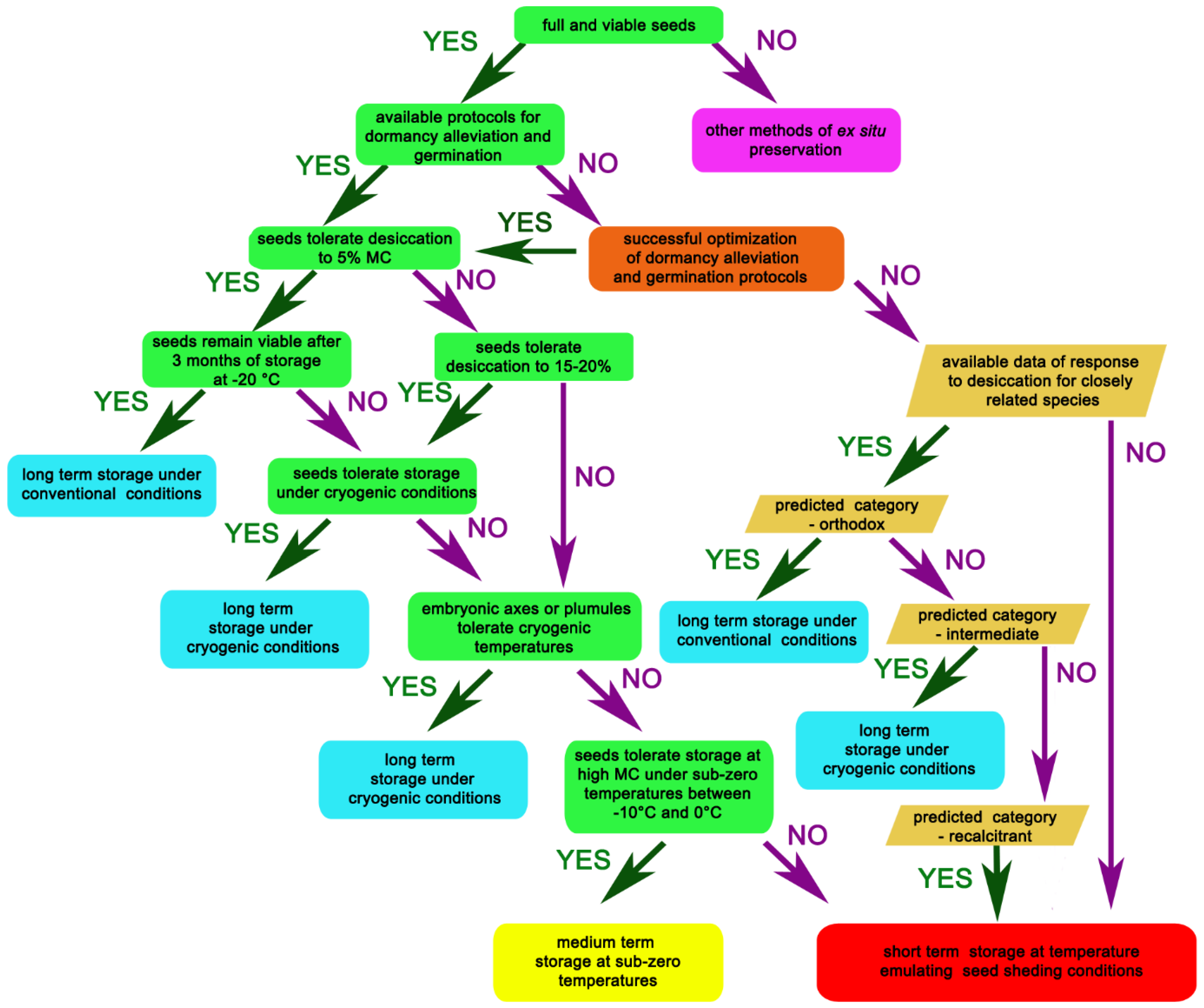
2. Important Questions That Need to Be Addressed Prior to Seed Storage
3. How to Successfully Store Seeds for a Long Time? The Role of Seed Moisture Content (MC) and Low Temperature in an Extension of Seed Longevity
4. Conventional Storage of Seeds
4.1. Conventional Storage of Orthodox Seeds
4.2. Conventional Storage of Intermediate Seeds
4.3. Conventional Storage of Recalcitrant Species
4.4. Conventional Storage of Woody Plant Species in Gene Banks
5. Cryogenic Storage of Seeds
5.1. Cryogenic Storage of Orthodox Seeds
5.2. Cryogenic Storage of Intermediate Seeds
5.3. Cryogenic Storage of Recalcitrant Seeds
5.4. Cryogenic Storage of Woody Plant Species at Gene Banks
6. Future Possibility of Improving Storability of Seeds with Conventional Methods
7. Future Prospects of Improving the Cryostorage of Seeds or Explants
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Roe, D.; Seddon, N.; Elliott, J. Biodiversity Loss—More than an Environmental Emergency. Lancet Planet. Health 2019, 3, e287–e289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soler, Z.; Kostecka, J. Biodiversity Loss, the Causes, the State and Basic Form of Nature Protection in Spain and Poland. Pol. J. Sustain. Dev. 2019, 22, 75–84. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate Change Threats to Plant Diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Applequist, W.; Brinckmann, J.; Cunningham, A.; Hart, R.; Heinrich, M.; Katerere, D.; Andel, T. Scientists’ Warning on Climate Change and Medicinal Plants. Planta Med. 2019, 86, 10–18. [Google Scholar] [CrossRef]
- Martin, M.A.; Sendra, O.A.; Bastos, A.; Bauer, N.; Bertram, C.; Blenckner, T.; Bowen, K.; Brando, P.M.; Rudolph, T.B.; Büchs, M.; et al. Ten New Insights in Climate Science 2021: A Horizon Scan. Glob. Sustain. 2021, 4, e25. [Google Scholar] [CrossRef]
- Aurelle, D.; Thomas, S.; Albert, C.; Bally, M.; Bondeau, A.; Boudouresque, C.-F.; Cahill, A.E.; Carlotti, F.; Chenuil, A.; Cramer, W.; et al. Biodiversity, Climate Change, and Adaptation in the Mediterranean. Ecosphere 2022, 13, e3915. [Google Scholar] [CrossRef]
- de Martins, R.S.; dos Santos Junior, N.A.; Barbedo, C.J. Seed Pathology of Non-Domesticated Species of Tropical Ecosystems. J. Seed Sci. 2022, 44, e202244029. [Google Scholar] [CrossRef]
- Kahane, R.; Hodgkin, T.; Jaenicke, H.; Hoogendoorn, C.; Hermann, M.; (Dyno) Keatinge, J.D.H.; d’Arros Hughes, J.; Padulosi, S.; Looney, N. Agrobiodiversity for Food Security, Health and Income. Agron. Sustain. Dev. 2013, 33, 671–693. [Google Scholar] [CrossRef] [Green Version]
- Corlett, R.T. Safeguarding Our Future by Protecting Biodiversity. Plant Divers. 2020, 42, 221. [Google Scholar] [CrossRef]
- Muluneh, M.G. Impact of Climate Change on Biodiversity and Food Security: A Global Perspective—A Review Article. Agric. Food Secur. 2021, 10, 36. [Google Scholar] [CrossRef]
- Hay, F.R.; Whitehouse, K.J.; Ellis, R.H.; Sackville Hamilton, N.R.; Lusty, C.; Ndjiondjop, M.N.; Tia, D.; Wenzl, P.; Santos, L.G.; Yazbek, M.; et al. CGIAR Genebank Viability Data Reveal Inconsistencies in Seed Collection Management. Glob. Food Secur. 2021, 30, 100557. [Google Scholar] [CrossRef]
- Singh, N.; Wu, S.; Raupp, W.J.; Sehgal, S.; Arora, S.; Tiwari, V.; Vikram, P.; Singh, S.; Chhuneja, P.; Gill, B.S.; et al. Efficient Curation of Genebanks Using next Generation Sequencing Reveals Substantial Duplication of Germplasm Accessions. Sci. Rep. 2019, 9, 650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Varaprasad, K.S.; Venkateswaran, K. Conservation Costs of Plant Genetic Resources for Food and Agriculture: Seed Genebanks. Agric. Res. 2012, 1, 223–239. [Google Scholar] [CrossRef] [Green Version]
- Panis, B.; Nagel, M.; Van den Houwe, I. Challenges and Prospects for the Conservation of Crop Genetic Resources in Field Genebanks, in In Vitro Collections and/or in Liquid Nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef] [PubMed]
- Jose Diez, M.; De la Rosa, L.; Martin, I.; Guasch, L.; Elena Cartea, M.; Mallor, C.; Casals, J.; Simo, J.; Rivera, A.; Anastasio, G.; et al. Plant Genebanks: Present Situation and Proposals for Their Improvement. the Case of the Spanish Network. Front. Plant Sci. 2018, 9, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Börner, A.; Khlestkina, E.K. Ex-Situ Genebanks—Seed Treasure Chambers for the Future. Russ. J. Genet. 2019, 55, 1299–1305. [Google Scholar] [CrossRef]
- Zimnoch-Guzowska, E.; Chmielarz, P.; Wawrzyniak, M.K.; Plitta-Michalak, B.P.; Michalak, M.; Pałucka, M.; Wasileńczyk, U.; Kosek, P.; Kulus, D.; Rucińska, A.; et al. Polish Cryobanks: Research and Conservation of Plant Genetic Resources. Acta Soc. Bot. Pol. 2022, 91, 9121. [Google Scholar] [CrossRef]
- Engels, J.M.M.; Ebert, A.W. A Critical Review of the Current Global Ex Situ Conservation System for Plant Agrobiodiversity. II. Strengths and Weaknesses of the Current System and Recommendations for Its Improvement. Plants 2021, 10, 1904. [Google Scholar] [CrossRef]
- Lusty, C.; Sackville Hamilton, R.; Guarino, L.; Richards, C.; Jamora, N.; Hawtin, G. Envisaging an Effective Global Long-Term Agrobiodiversity Conservation System That Promotes and Facilitates Use. Plants 2021, 10, 2764. [Google Scholar] [CrossRef]
- Lusty, C.; Guarino, L.; Toll, J.; Lainoff, B. Genebanks: Past, Present, and Optimistic Future. In Encyclopedia of Agriculture and Food Systems; Elsevier: Amsterdam, The Netherlands, 2014; pp. 417–432. ISBN 978-0-08-093139-5. [Google Scholar]
- Gonzalez-Arnao, M.T.; Martinez-Montero, M.E.; Cruz-Cruz, C.A.; Engelmann, F. Advances in Cryogenic Techniques for the Long-Term Preservation of Plant Biodiversity. In Sustainable Development and Biodiversity; Springer: Cham, Switzerland, 2014; pp. 129–170. [Google Scholar]
- Hay, F.; Sershen, N. New Technologies to Improve the Ex Situ Conservation of Plant Genetic Resources. In Plant Genetic Resources: A Review of Current Research and Future Needs; Burleigh Dodds Science Publishing: London, United Kingdom, 2020; ISBN 978-1-78676-451-5. [Google Scholar]
- Smith, P.P.; Trivedi, C.; Cochrane, A.; Crawford, A.; Way, M. The Millennium Seed Bank Project Delivering Target 8 of The. BG J. 2007, 4, 9–12. [Google Scholar]
- Ohga, I. The Germination of Century-Old and Recently Harvested Indian Lotus Fruits, with Special Reference to the Effect of Oxygen Supply. Am. J. Bot. 1926, 13, 754–759. [Google Scholar] [CrossRef]
- Porsild, A.E.; Harington, C.R.; Mulligan, G.A. Lupinus Arcticus Wats. Grown from Seeds of Pleistocene Age. Science 1967, 158, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Yashina, S.; Gubin, S.; Maksimovich, S.; Yashina, A.; Gakhova, E.; Gilichinsky, D. Regeneration of Whole Fertile Plants from 30,000-y-Old Fruit Tissue Buried in Siberian Permafrost. Proc. Natl. Acad. Sci. USA 2012, 109, 4008–4013. [Google Scholar] [CrossRef] [Green Version]
- Manning, S.W.; Griggs, C.; Lorentzen, B.; Ramsey, C.B.; Chivall, D.; Jull, A.J.T.; Lange, T.E. Fluctuating Radiocarbon Offsets Observed in the Southern Levant and Implications for Archaeological Chronology Debates. Proc. Natl. Acad. Sci. USA 2018, 115, 6141–6146. [Google Scholar] [CrossRef] [Green Version]
- Telewski, F.W.; Zeevaart, J.A.D. The 120-Yr Period for Dr. Beal’s Seed Viability Experiment. Am. J. Bot. 2002, 89, 1285–1288. [Google Scholar] [CrossRef] [Green Version]
- Kameswara Rao, N. Manual of Seed Handling in Genebanks; Bioversity International: Rome, Italy, 2006; ISBN 978-92-9043-740-6. [Google Scholar]
- Hong, T.D.; Ellis, R.H. A Protocol to Determine Seed Storage Behavior; IPGRI Technical Bulletin; International Plant Genetic Resources Institute: Rome, Italy, 1996; ISBN 978-92-9043-279-1. [Google Scholar]
- De Vitis, M.; Hay, F.R.; Dickie, J.B.; Trivedi, C.; Choi, J.; Fiegener, R. Seed Storage: Maintaining Seed Viability and Vigor for Restoration Use. Restor. Ecol. 2020, 28, S249–S255. [Google Scholar] [CrossRef]
- Pence, V.C.; Bruns, E.B. The Tip of the Iceberg: Cryopreservation Needs for Meeting the Challenge of Exceptional Plant Conservation. Plants 2022, 11, 1528. [Google Scholar] [CrossRef] [PubMed]
- Breman, E.; Ballesteros, D.; Castillo-Lorenzo, E.; Cockel, C.; Dickie, J.; Faruk, A.; O’Donnell, K.; Offord, C.A.; Pironon, S.; Sharrock, S.; et al. Plant Diversity Conservation Challenges and Prospects—The Perspective of Botanic Gardens and the Millennium Seed Bank. Plants 2021, 10, 2371. [Google Scholar] [CrossRef]
- Wyse, S.V.; Dickie, J.B.; Willis, K.J. Seed Banking Not an Option for Many Threatened Plants. Nat. Plants 2018, 4, 848–850. [Google Scholar] [CrossRef]
- Lan, Q.; Xia, K. Seed Storage Behaviour of 101 Woody Species from the Tropical Rainforest of Southern China: A Test of the Seed-Coat Ratio–Seed Mass (SCR–SM) Model for Determination of Desiccation Sensitivity. Aust. J. Bot. 2014, 62, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Montero, M.E.; Harding, K. Cryobionomics: Evaluating the Concept in Plant Cryopreservation. In PlantOmics: The Omics of Plant Science; Springer: New Delhi, India, 2015; pp. 655–682. [Google Scholar]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.-C. Cryobiotechnologies: Tools for Expanding Long-Term Ex Situ Conservation to All Plant Species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Walters, C.; Pence, V.C. The Unique Role of Seed Banking and Cryobiotechnologies in Plant Conservation. Plants People Planet 2021, 3, 83–91. [Google Scholar] [CrossRef]
- Benelli, C. Plant Cryopreservation: A Look at the Present and the Future. Plants 2021, 10, 2744. [Google Scholar] [CrossRef] [PubMed]
- Pence, V.C.; Meyer, A.; Linsky, J.; Gratzfeld, J.; Pritchard, H.W.; Westwood, M.; Bruns, E.B. Defining Exceptional Species—A Conceptual Framework to Expand and Advance Ex Situ Conservation of Plant Diversity beyond Conventional Seed Banking. Biol. Conserv. 2022, 266, 109440. [Google Scholar] [CrossRef]
- Pence, V.C. In Vitro Methods and the Challenge of Exceptional Species for Target 8 of the Global Strategy for Plant Conservation1. Ann. Mo. Bot. Gard. 2013, 99, 214–220. [Google Scholar] [CrossRef]
- Kilgore, S.; Havens, K.; Kramer, A.; Lythgoe, A.; MacKechnie, L.; Vitis, M.D. Seed Collection, Storage, and Germination Practices May Affect Viola Reintroduction Outcomes. Nativ. Plants J. 2022, 23, 40–55. [Google Scholar] [CrossRef]
- Lopez Del Egido, L.; Navarro-Miró, D.; Martinez-Heredia, V.; Toorop, P.E.; Iannetta, P.P.M. A Spectrophotometric Assay for Robust Viability Testing of Seed Batches Using 2,3,5-Triphenyl Tetrazolium Chloride: Using Hordeum vulgare L. as a Model. Front. Plant Sci. 2017, 8, 747. [Google Scholar] [CrossRef] [Green Version]
- Musaev, F.; Priyatkin, N.; Potrakhov, N.; Beletskiy, S.; Chesnokov, Y. Assessment of Brassicaceae Seeds Quality by X-Ray Analysis. Horticulturae 2022, 8, 29. [Google Scholar] [CrossRef]
- Kavelenova, L.; Roguleva, N.; Yankov, N.; Ruzaeva, I.; Pavlova, E.; Nakrainikova, D.; Potrachov, N. Assessment of the Quality of Seeds Formed in Situ and Ex Situ as a Mandatory Element of Maintaining Seed Banks of Rare Plants. E3S Web Conf. 2021, 265, 05012. [Google Scholar] [CrossRef]
- Baskin, C.; Baskin, J.M. Seed Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: Cambridge, MA, USA, 2014; p. 1586. [Google Scholar]
- Kildisheva, O.A.; Dixon, K.W.; Silveira, F.A.O.; Chapman, T.; Di Sacco, A.; Mondoni, A.; Turner, S.R.; Cross, A.T. Dormancy and Germination: Making Every Seed Count in Restoration. Restor. Ecol. 2020, 28, S256–S265. [Google Scholar] [CrossRef]
- Wyse, S.; Dickie, J. Taxonomic Affinity, Habitat and Seed Mass Strongly Predict Seed Desiccation Response: A Boosted Regression Trees Analysis Based on 17 539 Species. Ann. Bot. 2017, 121, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Ņečajeva, J.; Bleidere, M.; Jansone, Z.; Gailīte, A.; Ruņģis, D. Variability of Seed Germination and Dormancy Characteristics and Genetic Analysis of Latvian Avena Fatua Populations. Plants 2021, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Cristaudo, A.; Gresta, F.; Restuccia, A.; Catara, S.; Onofri, A. Germinative Response of Redroot Pigweed (Amaranthus retroflexus L.) to Environmental Conditions: Is There a Seasonal Pattern? Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2016, 150, 583–591. [Google Scholar] [CrossRef]
- Li, R.; Chen, L.; Wu, Y.; Zhang, R.; Baskin, C.C.; Baskin, J.M.; Hu, X. Effects of Cultivar and Maternal Environment on Seed Quality in Vicia Sativa. Front. Plant Sci. 2017, 8, 1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, C.; Baskin, J. When Breaking Seed Dormancy Is a Problem Try a Move-along Experiment. Nativ. Plants J. 2003, 4, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Baskin, C.; Thompson, K.; Baskin, J. Mistakes in Germination Ecology and How to Avoid Them. Seed Sci. Res. 2006, 16, 165–168. [Google Scholar] [CrossRef]
- Roberts, E.H. Predicting the Storage Life of Seeds. Seed Sci. Technol. 1973, 1, 499–514. [Google Scholar]
- Barbedo, C.J.; da Centeno, D.C.; de Ribeiro, R.C.L.F. Do Recalcitrant Seeds Really Exist? Hoehnea 2013, 40, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Sershen; Varghese, B.; Naidoo, C.; Pammenter, N.W. The Use of Plant Stress Biomarkers in Assessing the Effects of Desiccation in Zygotic Embryos from Recalcitrant Seeds: Challenges and Considerations. Plant Biol. 2016, 18, 433–444. [Google Scholar] [CrossRef]
- Plitta-Michalak, B.P.; Naskret-Barciszewska, M.Z.; Kotlarski, S.; Tomaszewski, D.; Tylkowski, T.; Barciszewski, J.; Chmielarz, P.; Michalak, M. Changes in Genomic 5-Methylcytosine Level Mirror the Response of Orthodox (Acer platanoides L.) and Recalcitrant (Acer pseudoplatanus L.) Seeds to Severe Desiccation. Tree Physiol. 2018, 38, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Matilla, A. The Orthodox Dry Seeds Are Alive: A Clear Example of Desiccation Tolerance. Plants 2021, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.P.N.; Salatino, M.L.F.; Barbedo, C.J.; de Figueiredo-Ribeiro, R.C.L.; da Centeno, D.C. Active Metabolism during Desiccation of the Desiccation Tolerant Short-Lived Seeds of Poincianella pluviosa (DC.) L. P. Queiroz. J. Seed Sci. 2022, 44, e202244028. [Google Scholar] [CrossRef]
- Bharuth, V.; Naidoo, C. Responses to Cryopreservation of Recalcitrant Seeds of Ekebergia Capensis from Different Provenances. S. Afr. J. Bot. 2020, 132, 1–14. [Google Scholar] [CrossRef]
- FAO (Ed.) Genebank Standards for Plant Genetic Resources for Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; ISBN 978-92-5-107855-6. [Google Scholar]
- Walters, C. Orthodoxy, Recalcitrance and in-between: Describing Variation in Seed Storage Characteristics Using Threshold Responses to Water Loss. Planta 2015, 242, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An Intermediate Category of Seed Storage Behaviour? I. COFFEE. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Jacobo-Pereira, C.; Muñiz-Castro, M.Á.; Vázquez-García, J.A.; Flores, J.; Muñoz-Urias, A.; Huerta-Martínez, F.M.; Jacobo-Pereira, C.; Muñiz-Castro, M.Á.; Vázquez-García, J.A.; Flores, J.; et al. Seed Integrity, Effect of Temperature and Storage Time on Germination of Populus Luziarum and P. Primaveralepensis, Endangered Subtropical Species from Mexico. Bot. Sci. 2022, 100, 192–203. [Google Scholar] [CrossRef]
- Noor, N.; Reed, B.M.; Yu, X. Seed Storage and Cryoexposure Behavior in Hazelnut (Corylus avellana L. Cv. Barcelona). CryoLetters 1994, 15, 315–322. [Google Scholar]
- Michalak, M.; Plitta, B.P.; Chmielarz, P. Desiccation Sensitivity and Successful Cryopreservation of Oil Seeds of European Hazelnut (Corylus Avellana). Ann. Appl. Biol. 2013, 163, 351–358. [Google Scholar] [CrossRef]
- Pipinis, E.; Stampoulidis, A.; Milios, E.; Kitikidou, K.; Akritidou, S.; Theodoridou, S.; Radoglou, K. Effects of Seed Moisture Content, Stratification and Sowing Date on the Germination of Corylus Avellana Seeds. J. For. Res. 2020, 31, 743–749. [Google Scholar] [CrossRef]
- Harrington, J.F. Thumb Rules of Drying Seeds. Crops Soils 1960, 13, 16–17. [Google Scholar]
- Harrington, J.F. Seed Storage and Longevity. In Insects, and Seed Collection, Storage, Testing, and Certification; Elsevier: Amsterdam, The Netherlands, 1972; pp. 145–245. ISBN 978-0-12-395605-7. [Google Scholar]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. A Comparison of the Low-Moisture-Content Limit to the Logarithmic Relation Between Seed Moisture and Longevity in Twelve Species. Ann. Bot. 1989, 63, 601–611. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. The Low-Moisture-Content Limit to the Negative Logarithmic Relation Between Seed Longevity and Moisture Content in Three Subspecies of Rice. Ann. Bot. 1992, 69, 53–58. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. Moisture Content and the Longevity of Seeds of Phaseolus Vulgaris. Ann. Bot. 1990, 66, 341–348. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H.; Tao, K.-L. Low Moisture Content Limits to Relations Between Seed Longevity and Moisture. Ann. Bot. 1990, 65, 493–504. [Google Scholar] [CrossRef]
- Vertucci, C.W.; Roos, E.E. Theoretical Basis of Protocols for Seed Storage. Plant Physiol. 1990, 94, 1019–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vertucci, C.W.; Roos, E.E. Theoretical Basis of Protocols for Seed Storage II. The Influence of Temperature on Optimal Moisture Levels. Seed Sci. Res. 1993, 3, 201–213. [Google Scholar] [CrossRef]
- Vertucci, C.W.; Roos, E.E.; Crane, J. Theoretical Basis of Protocols for Seed Storage III. Optimum Moisture Contents for Pea Seeds Stored at Different Temperatures. Ann. Bot. 1994, 74, 531–540. [Google Scholar] [CrossRef]
- Pritchard, H.; Wood, C.B.; Hodges, S.S.; Vautier, H.J. 100-Seed Test for Desiccation Tolerance and Germination: A Case Study on Eight Tropical Palm Species. Seed Sci. Technol. 2004, 32, 393–403. [Google Scholar] [CrossRef]
- Mattana, E.; Peguero, B.; Di Sacco, A.; Agramonte, W.; Encarnación Castillo, W.R.; Jiménez, F.; Clase, T.; Pritchard, H.W.; Gómez-Barreiro, P.; Castillo-Lorenzo, E.; et al. Assessing Seed Desiccation Responses of Native Trees in the Caribbean. New For. 2020, 51, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Prediction of Desiccation Sensitivity in Seeds of Woody Species: A Probabilistic Model Based on Two Seed Traits and 104 Species. Ann. Bot. 2006, 97, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Mayrinck, R.C.; Vilela, L.C.; Pereira, T.M.; Rodrigues-Junior, A.G.; Davide, A.C.; Vaz, T.A.A. Seed Desiccation Tolerance/Sensitivity of Tree Species from Brazilian Biodiversity Hotspots: Considerations for Conservation. Trees 2019, 33, 777–785. [Google Scholar] [CrossRef]
- Chau, M.M.; Chambers, T.; Weisenberger, L.; Keir, M.; Kroessig, T.I.; Wolkis, D.; Kam, R.; Yoshinaga, A.Y. Seed Freeze Sensitivity and Ex Situ Longevity of 295 Species in the Native Hawaiian Flora. Am. J. Bot. 2019, 106, 1248–1270. [Google Scholar] [CrossRef] [PubMed]
- Porceddu, M.; Santo, A.; Orrù, M.; Meloni, F.; Ucchesu, M.; Picciau, R.; Sarigu, M.; Cuena Lombraña, A.; Podda, L.; Sau, S.; et al. Seed Conservation Actions for the Preservation of Plant Diversity: The Case of the Sardinian Germplasm Bank (BG-SAR). Plant Sociol. 2017, 54, 111–117. [Google Scholar] [CrossRef]
- Bacchetta, G.; Belletti, P.; Brullo, S.; Cagelli, L.; Carasso, V.; Casas, J.L.; Cervelli, C.; Escribà, M.C.; Fenu, G.; Gorian, F. Manual for the Collection, Study, Ex Situ Processing and Conservation of Germplasm; Agenzia per la Protezione Dell’ambiente e per i Servizi tecnici: Rome, Italy, 2006; ISBN 88-448-0179-5. [Google Scholar]
- Kumar, S.; Jalli (J), R.; Srinivasan, K. Physiological and Biochemical Changes in the Seeds of Karanj (Pongamia Pinnata L) under Different Storage Conditions. Indian J. Agric. Sci. 2011, 81, 423–428. [Google Scholar]
- Magrini, S.; De Vitis, M.; Torelli, D.; Santi, L.; Zucconi, L. Seed Banking of Terrestrial Orchids: Evaluation of Seed Quality in Anacamptis Following 4-Year Dry Storage. Plant Biol. 2019, 21, 544–550. [Google Scholar] [CrossRef]
- Dhyani, A.; Baskin, C.; Nautiyal, B.; Nautiyal, M. Overcoming Root Dormancy and Identifying the Storage Behaviour of Lilium Polyphyllum Seeds. Botany 2019, 97, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Solberg, S.Ø.; Yndgaard, F.; Andreasen, C.; von Bothmer, R.; Loskutov, I.G.; Asdal, Å. Long-Term Storage and Longevity of Orthodox Seeds: A Systematic Review. Front. Plant Sci. 2020, 11, 1007. [Google Scholar] [CrossRef]
- Solberg, S.Ø.; Brodal, G.; von Bothmer, R.; Meen, E.; Yndgaard, F.; Andreasen, C.; Asdal, Å. Seed Germination after 30 Years Storage in Permafrost. Plants 2020, 9, 579. [Google Scholar] [CrossRef]
- Wawrzyniak, M.K.; Michalak, M.; Chmielarz, P. Effect of Different Conditions of Storage on Seed Viability and Seedling Growth of Six European Wild Fruit Woody Plants. Ann. For. Sci. 2020, 77, 58. [Google Scholar] [CrossRef]
- Liu, U.; Cossu, T.A.; Davies, R.M.; Forest, F.; Dickie, J.B.; Breman, E. Conserving Orthodox Seeds of Globally Threatened Plants Ex Situ in the Millennium Seed Bank, Royal Botanic Gardens, Kew, UK: The Status of Seed Collections. Biodivers. Conserv. 2020, 29, 2901–2949. [Google Scholar] [CrossRef]
- Plitta-Michalak, B.P.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P.; Michalak, M. Epigenetic Integrity of Orthodox Seeds Stored under Conventional and Cryogenic Conditions. Forests 2021, 12, 288. [Google Scholar] [CrossRef]
- Desheva, G. The Longevity of Crop Seeds Stored Under Long-Term Condition in the National Gene Bank of Bulgaria. Agric. Polnohospodárstvo 2016, 62, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of Seeds Stored in a Genebank: Species Characteristics. Seed Sci. Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Colville, L.; Bradley, E.L.; Lloyd, A.S.; Pritchard, H.W.; Castle, L.; Kranner, I. Volatile Fingerprints of Seeds of Four Species Indicate the Involvement of Alcoholic Fermentation, Lipid Peroxidation, and Maillard Reactions in Seed Deterioration during Ageing and Desiccation Stress. J. Exp. Bot. 2012, 63, 6519–6530. [Google Scholar] [CrossRef]
- Mira, S.; Hill, L.M.; González-Benito, M.E.; Ibáñez, M.A.; Walters, C. Volatile Emission in Dry Seeds as a Way to Probe Chemical Reactions during Initial Asymptomatic Deterioration. J. Exp. Bot. 2016, 67, 1783–1793. [Google Scholar] [CrossRef]
- Mira, S.; Pirredda, M.; Martín-Sánchez, M.; Marchessi, J.E.; Martín, C. DNA Methylation and Integrity in Aged Seeds and Regenerated Plants. Seed Sci. Res. 2020, 30, 92–100. [Google Scholar] [CrossRef]
- González-Benito, M.E.; Ibáñez, M.Á.; Pirredda, M.; Mira, S.; Martín, C. Application of the MSAP Technique to Evaluate Epigenetic Changes in Plant Conservation. Int. J. Mol. Sci. 2020, 21, 7459. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.P.; Nadarajan, J.; Colville, L. Volatile Signature Indicates Viability of Dormant Orthodox Seeds. Physiol. Plant. 2021, 173, 788–804. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.P.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. DNA Methylation as an Early Indicator of Aging in Stored Seeds of “Exceptional” Species Populus nigra L. Cells 2022, 11, 2080. [Google Scholar] [CrossRef]
- León-Lobos, P.; Ellis, R.H. Seed Storage Behaviour of Fagus Sylvatica and Fagus Crenata. Seed Sci. Res. 2002, 12, 31–37. [Google Scholar] [CrossRef]
- Stavrinides, A.K.; Dussert, S.; Combes, M.-C.; Fock-Bastide, I.; Severac, D.; Minier, J.; Bastos-Siqueira, A.; Demolombe, V.; Hem, S.; Lashermes, P.; et al. Seed Comparative Genomics in Three Coffee Species Identify Desiccation Tolerance Mechanisms in Intermediate Seeds. J. Exp. Bot. 2020, 71, 1418–1433. [Google Scholar] [CrossRef] [PubMed]
- Popova, E.V.; Kim, D.H.; Han, S.H.; Pritchard, H.W.; Lee, J.C. Narrowing of the Critical Hydration Window for Cryopreservation of Salix Caprea Seeds Following Ageing and a Reduction in Vigour. CryoLetters 2012, 33, 220–231. [Google Scholar] [PubMed]
- Suszka, J.; Plitta, B.P.; Michalak, M.; Bujarska-Borkowska, B.; Tylkowski, T.; Chmielarz, P. Optimal Seed Water Content and Storage Temperature for Preservation of Populus nigra L. Germplasm. Ann. For. Sci. 2014, 71, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Michalak, M.; Plitta, B.P.; Tylkowski, T.; Chmielarz, P.; Suszka, J. Desiccation Tolerance and Cryopreservation of Seeds of Black Poplar (Populus nigra L.), a Disappearing Tree Species in Europe. Eur. J. For. Res. 2015, 134, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Bareke, T.; Addi, A.; Jilo, K.; Kumsa, T. Effect of Storage Temperature and Packing Materials on Seed Germination and Seed Storage Behavior of Schefflera Abyssinica. Nusant. Biosci. 2022, 14, 141–147. [Google Scholar] [CrossRef]
- Umarani, R.; Aadhavan, E.K.; Faisal, M.M. Understanding Poor Storage Potential of Recalcitrant Seeds. Curr. Sci. 2015, 108, 2023–2034. [Google Scholar]
- Suszka, B.; Tylkowski, T. Storage of acorns of the English oak (Quercus robur L.) over 1-5 winters. Arbor. Kórn. 1980, 25, 199–229. [Google Scholar]
- Michalak, M.; Plitta-Michalak, B.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Bujarska-Borkowska, B.; Chmielarz, P. Global 5-Methylcytosine Alterations in DNA during Ageing of Quercus Robur Seeds. Ann. Bot. 2015, 116, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Chmielarz, P.; Suszka, J.; Wawrzyniak, M.K. Desiccation Does Not Increase Frost Resistance of Pedunculate Oak (Quercus robur L.) Seeds. Ann. For. Sci. 2022, 79, 3. [Google Scholar] [CrossRef]
- Tylkowski, T. Short-Term Storage of after Ripened Seeds of Acer platanoides L. and A. pseudoplatanus L. Arbor. Kórn. 1989, 34, 135–141. [Google Scholar]
- Lan, Q.; Luo, Y.L.; Ma, S.; Lu, X.; Yang, M.-Z.; Tan, Y.; Jiang, X.; Tan, Y.; Wang, X.; Li, Z. Development and Storage of Recalcitrant Seeds of Hopea Hainanensis. Seed Sci. Technol. 2012, 40, 200–208. [Google Scholar] [CrossRef]
- Pałucka, M.; Hrydziuszko, P. Cryogenic Storage of Plant Matrial in the Kostrzyca Forest Gene Bank. In Current Technologies of Forest Seed Treatment; Muller, J., Kozioł, C., Pałucka, M., Eds.; The Kostrzyca Forest Gene Bank, Milkow, Poland: 2014; ISBN 978-83-88245-12-1.
- Benson, E.E. Cryopreservation Theory. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; pp. 15–32. ISBN 978-0-387-72276-4. [Google Scholar]
- Streczynski, R.; Clark, H.; Whelehan, L.M.; Ang, S.-T.; Hardstaff, L.K.; Funnekotter, B.; Bunn, E.; Offord, C.A.; Sommerville, K.D.; Mancera, R.L. Current Issues in Plant Cryopreservation and Importance for Ex Situ Conservation of Threatened Australian Native Species. Aust. J. Bot. 2019, 67, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Panis, B. Sixty Years of Plant Cryopreservation: From Freezing Hardy Mulberry Twigs to Establishing Reference Crop Collections for Future Generations. Acta Hortic. 2019, 1234, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Reed, B.M. Plant Cryopreservation: A Continuing Requirement for Food and Ecosystem Security. In Vitro Cell. Dev. Biol. Plant 2017, 53, 285–288. [Google Scholar] [CrossRef]
- Wowk, B. Thermodynamic Aspects of Vitrification. Cryobiology 2010, 60, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Wheeler, L.; Stanwood, P.C. Longevity of Cryogenically Stored Seeds. Cryobiology 2004, 48, 229–244. [Google Scholar] [CrossRef]
- Ballesteros, D.; Pence, V.C. Survival and Death of Seeds during Liquid Nitrogen Storage: A Case Study on Seeds with Short Lifespans. CryoLetters 2017, 38, 278–289. [Google Scholar]
- Choudhary, P.; Pramitha, L.; Aggarwal, P.R.; Rana, S.; Vetriventhan, M.; Muthamilarasan, M. Biotechnological Interventions for Improving the Seed Longevity in Cereal Crops: Progress and Prospects. Crit. Rev. Biotechnol. 2022, 1–17. [Google Scholar] [CrossRef]
- Lu, J.; Greene, S.; Reid, S.; Cruz, V.M.V.; Dierig, D.A.; Byrne, P. Phenotypic Changes and DNA Methylation Status in Cryopreserved Seeds of Rye (Secale cereale L.). Cryobiology 2018, 82, 8–14. [Google Scholar] [CrossRef]
- Zevallos, B.; Cejas, I.; Escriba, R.C.; Yabor, L.; Aragón Abreu, C.E.; Gonzalez Olmedo, J.; Engelmann, F.; Martinez-Montero, M.; Lorenzo Feijoo, J. Biochemical Characterization of Ecuadorian Wild Solanum Lycopersicum Mill. Plants Produced from Non-Cryopreserved and Cryopreserved Seeds. CryoLetters 2013, 34, 413–421. [Google Scholar]
- Chmielarz, P. Cryopreservation of Dormant European Ash (Fraxinus Excelsior) Orthodox Seeds. Tree Physiol. 2009, 29, 1279–1285. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Wu, G.R.; Song, X.Y.; Liu, B.L. Cryopreservation of Dormant Prunus armeniaca L. Seeds. Seed Sci. Technol. 2015, 43, 456–466. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.; Chmielarz, P. Desiccation Tolerance and Cryopreservation of Wild Apple (Malus Sylvestris) Seeds. Seed Sci. Technol. 2015, 43, 480–491. [Google Scholar] [CrossRef]
- Reed, B.M.; Schwanke, S.; Shala, R. Pear Seeds Retain Viability after Liquid Nitrogen Immersion. HortScience 2001, 36, 1121–1122. [Google Scholar] [CrossRef] [Green Version]
- Chmielarz, P. Cryopreservation of Dormant Orthodox Seeds of Forest Trees: Mazzard Cherry (Prunus avium L.). Ann. For. Sci. 2009, 66, 405. [Google Scholar] [CrossRef] [Green Version]
- Michalak, M.; Plitta-Michalak, B.; Chmielarz, P. A New Insight in Desiccation Tolerance and Cryopreservation of Mazzard Cherry (Prunus avium L.) Seeds. Cent. Eur. J. Biol. 2015, 10, 354–364. [Google Scholar] [CrossRef]
- Chmielarz, P. Cryopreservation of Dormant Orthodox Seeds of European Hornbeam (Carpinus Betulus). Seed Sci. Technol. 2010, 38, 146–157. [Google Scholar] [CrossRef]
- Normah, M.; Vengadasalam, M. Effects of Moisture-Content on Cryopreservation of Coffea and Vigna Seeds and Embryos. CryoLetters 1992, 13, 199–208. [Google Scholar]
- Hor, Y.L.; Kim, Y.J.; Ugap, A.; Chabrillange, N.; Sinniah, U.R.; Engelmann, F.; Dussert, S. Optimal Hydration Status for Cryopreservation of Intermediate Oily Seeds: Citrus as a Case Study. Ann. Bot. 2005, 95, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Wood, C.; Pritchard, H.; Lindegaard, K. Seed Cryopreservation and Longevity of Two Salix Hybrids. CryoLetters 2003, 24, 17–26. [Google Scholar]
- Ku, J.J.; Han, S.H.; Kim, D.H. Extended Low Temperature and Cryostorage Longevity of Salix Seeds with Desiccation Control. Open Life Sci. 2019, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pence, V.C. Germination, Desiccation and Cryopreservation of Seeds of Populus Deltoides Bartr. In Proceedings of the International Seed Testing Association, Opocno, Czech Republic, 9–11 October 1996. [Google Scholar]
- Walters, C.; Touchell, D.H.; Power, P.; Wesley-Smith, J.; Antolin, M.F. A Cryopreservation Protocol for Embryos of the Endangered Species Zizania Texana. CryoLetters 2002, 23, 291–298. [Google Scholar] [PubMed]
- Hazubska-Przybył, T.; Chmielarz, P.; Michalak, M.; Dering, M.; Bojarczuk, K. Survival and Genetic Stability of Picea Abies Embryogenic Cultures after Cryopreservation Using a Pregrowth-Dehydration Method. Plant Cell Tissue Organ Cult. PCTOC 2013, 113, 303–313. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. Cryostorage of Germplasm of Tropical Recalcitrant-Seeded Species: Approaches and Problems. Int. J. Plant Sci. 2014, 175, 29–39. [Google Scholar] [CrossRef]
- Ballesteros, D.; Sershen; Varghese, B.; Berjak, P.; Pammenter, N.W. Uneven Drying of Zygotic Embryos and Embryonic Axes of Recalcitrant Seeds: Challenges and Considerations for Cryopreservation. Cryobiology 2014, 69, 100–109. [Google Scholar] [CrossRef]
- Xia, K.; Hill, L.M.; Li, D.-Z.; Walters, C. Factors Affecting Stress Tolerance in Recalcitrant Embryonic Axes from Seeds of Four Quercus (Fagaceae) Species Native to the USA or China. Ann. Bot. 2014, 114, 1747–1759. [Google Scholar] [CrossRef]
- Malik, S.; Choudhary, R.; Kaur, S.; Chaudhury, R.; Pritchard, H. Storage Behavior and Cryopreservation of Citrus Cavaleriei, an Endangered, Cold-Resistant Species of Northeast India with Exceptionally Large Seeds. CryoLetters 2020, 41, 281–290. [Google Scholar]
- Gaidamashvili, M.; Khurtsidze, E.; Kutchava, T.; Lambardi, M.; Benelli, C. Efficient Protocol for Improving the Development of Cryopreserved Embryonic Axes of Chestnut (Castanea Sativa Mill.) by Encapsulation-Vitrification. Plants 2021, 10, 231. [Google Scholar] [CrossRef]
- Srivastava, V.; Hajong, S.; Chandora, R.; Agrawal, A. Desiccation and Freezing Tolerance of Recalcitrant Seeds and Embryonic Axes of Prunus Napaulensis (Ser.) Steud.: A Crop Wild Relative of Cherry. Genet. Resour. Crop Evol. 2022, 69, 1571–1583. [Google Scholar] [CrossRef]
- N’Nan, O.; Hocher, V.; Verdeil, J.-L.; Konan, J.-L.; Ballo, K.; Mondeil, F.; Malaurie, B. Cryopreservation by Encapsulation-Dehydration of Plumules of Coconut (Cocos nucifera L.). Cryoletters 2008, 29, 339–350. [Google Scholar]
- Chmielarz, P.; Michalak, M.; Pałucka, M.; Wasilenczyk, U. Successful Cryopreservation of Quercus Robur Plumules. Plant Cell Rep. 2011, 30, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Plitta, B.P.; Michalak, M.; Naskręt-Barciszewska, M.Z.; Barciszewski, J.; Chmielarz, P. DNA Methylation of Quercus robur L. Plumules Following Cryo-Pretreatment and Cryopreservation. Plant Cell Tissue Organ Cult. PCTOC 2014, 117, 31–37. [Google Scholar] [CrossRef]
- Dickie, J.B.; May, K.; Morris, S.V.A.; Titley, S.E. The Effects of Desiccation on Seed Survival in Acer platanoides L. and Acer pseudoplatanus L. Seed Sci. Res. 1991, 1, 149–162. [Google Scholar] [CrossRef]
- Finchsavage, W. Embryo Water Status and Survival in the Recalcitrant Species Quercus-Robur L—Evidence for a Critical Moisture-Content. J. Exp. Bot. 1992, 43, 663–669. [Google Scholar] [CrossRef]
- Tao, D.; Li, P. Classification of Plant-Cell Cryoprotectants. J. Theor. Biol. 1986, 123, 305–310. [Google Scholar] [CrossRef]
- Sakai, A.; Engelmann, F. Vitrification, Encapsulation-Vitrification and Droplet-Vitrification: A Review. Cryoletters 2007, 28, 151–172. [Google Scholar]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of Nucellar Cells of Navel Orange (Citrus-Sinensis Osb Var Brasiliensis Tanaka) by Vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Zhao, M.A.; Zhu, Y.Z.; Dhital, S.P.; Khu, D.M.; Song, Y.S.; Wang, M.H.; Lim, H.T. An Efficient Cryopreservation Procedure for Potato (Solanum tuberosum L.) Utilizing the New Ice Blocking Agent, Supercool X1000. Plant Cell Rep. 2006, 25, 164. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of Asparagus (Asparagus officinalis L.) Embryogenic Suspension Cells and Subsequent Plant Regeneration by Vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Valencia-Quintana, R.; Gómez-Arroyo, S.; Waliszewski, S.M.; Sánchez-Alarcón, J.; Gómez-Olivares, J.L.; Flores-Márquez, A.R.; Cortés-Eslava, J.; Villalobos-Pietrini, R. Evaluation of the Genotoxic Potential of Dimethyl Sulfoxide (DMSO) in Meristematic Cells of the Root of Vicia Faba. Toxicol. Environ. Health Sci. 2012, 4, 154–160. [Google Scholar] [CrossRef]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected Low-Dose Toxicity of the Universal Solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Awan, M.; Buriak, I.; Fleck, R.; Fuller, B.; Goltsev, A.; Kerby, J.; Lowdell, M.; Mericka, P.; Petrenko, A.; Petrenko, Y.; et al. Dimethyl Sulfoxide: A Central Player since the Dawn of Cryobiology, Is Efficacy Balanced by Toxicity? Regen. Med. 2020, 15, 1463–1491. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, E.A.; Hanson, A.; Communications, E. Summary Report World Commission on Forests and Sustainable Development; World Commission on Forests & Sustainable Development: Winnipeg, MB, Canada, 1999. [Google Scholar]
- Li, D.-Z.; Pritchard, H.W. The Science and Economics of Ex Situ Plant Conservation. Trends Plant Sci. 2009, 14, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Hay, F.R.; Probert, R.J. Advances in Seed Conservation of Wild Plant Species: A Review of Recent Research. Conserv. Physiol. 2013, 1, cot030. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.D.; Kumaria, S.; Das, M.C. Development of Cryopreservation Protocol for Aquilaria Malaccensis Lam., a Recalcitrant Seeded Tropical Tree Species. Cryoletters 2019, 40, 18–27. [Google Scholar]
- Nadarajan, J.; Staines, H.J.; Benson, E.E.; Marzalina, M.; Krishnapillay, B.; Harding, K. Optimization of Cryopreservation Protocol for Sterculia Cordata Zygotic Embryos Using Taguchi Experiments. J. Trop. For. Sci. 2006, 18, 222–230. [Google Scholar]
- Luis, Z.G.; Scherwinski-Pereira, J.E. A Simple and Efficient Protocol for the Cryopreservation of Zygotic Embryos of Macaw Palm [Acrocomia Aculeata (Jacq.) Lodd. Ex Mart.], A Tropical Species with A Capacity for Biofuel Production. Cryoletters 2017, 38, 7–16. [Google Scholar]
- Hor, Y.L.; Stanwood, P.C.; Chin, H.F. Effect of Dehydration on Freezing Characteristics and Survival in Liquid Nitrogen of Three Recalcitrant Seeds. Pertanika 1990, 13, 309–314. [Google Scholar]
- Normah, M.N.; Jamilah, M.S.; Serimala, S.D. Viability Studies on Seeds and Embryonic Axes of Lansium Domesticum Corr. Malays. Appl. Biol. 1996, 25, 39–43. [Google Scholar]
- Normah, M.N.; Ramiya, S.D.; Gintangga, M. Desiccation Sensitivity of Recalcitrant Seeds—A Study on Tropical Fruit Species. Seed Sci. Res. 1997, 7, 179–183. [Google Scholar] [CrossRef]
- Groot, S.P.C.; de Groot, L.; Kodde, J.; van Treuren, R. Prolonging the Longevity of Ex Situ Conserved Seeds by Storage under Anoxia. Plant Genet. Resour.-Charact. Util. 2015, 13, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.; Ellis, R. Water and Seed Survival. Ann. Bot. 1989, 63, 39–52. [Google Scholar] [CrossRef]
- Abadia, M.B.; San Martino, S.; Bartosik, R.E. Can Anoxic Atmospheres Protect the Quality of Maize Seeds during Storage? J. Stored Prod. Res. 2022, 96, 101927. [Google Scholar] [CrossRef]
- Han, B.; Fernandez, V.; Pritchard, H.W.; Colville, L. Gaseous Environment Modulates Volatile Emission and Viability Loss during Seed Artificial Ageing. Planta 2021, 253, 106. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, L.; Lu, Q. Exploration of Mechanisms for Internal Deterioration of Wheat Seeds in Postharvest Storage and Nitrogen Atmosphere Control for Properties Protection. Crop Sci. 2018, 58, 823–836. [Google Scholar] [CrossRef]
- Moncini, L.; Simone, G.; Romi, M.; Cai, G.; Guerriero, G.; Whittaker, A.; Benedettelli, S.; Berni, R. Controlled Nitrogen Atmosphere for the Preservation of Functional Molecules during Silos Storage: A Case Study Using Old Italian Wheat Cultivars. J. Stored Prod. Res. 2020, 88, 101638. [Google Scholar] [CrossRef]
- Xia, F.; Cheng, H.; Chen, L.; Zhu, H.; Mao, P.; Wang, M. Influence of Exogenous Ascorbic Acid and Glutathione Priming on Mitochondrial Structural and Functional Systems to Alleviate Aging Damage in Oat Seeds. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.-J.; Chen, M.-X.; Song, T.; Cheng, C.-L.; Tian, Y.; Hu, J.; Zhang, J.-H. Spermidine Enhanced the Antioxidant Capacity of Rice Seeds during Seed Aging. Plant Growth Regul. 2020, 91, 397–406. [Google Scholar] [CrossRef]
- Adetunji, A.E.; Adetunji, T.L.; Varghese, B.; Sershen; Pammenter, N.W. Oxidative Stress, Ageing and Methods of Seed Invigoration: An Overview and Perspectives. Agronomy 2021, 11, 2369. [Google Scholar] [CrossRef]
- Das, G.; Dutta, P. Effect of Nanopriming with Zinc Oxide and Silver Nanoparticles on Storage of Chickpea Seeds and Management of Wilt Disease. J. Agric. Sci. Technol. 2022, 24, 213–226. [Google Scholar]
- Kaur, R.; Chandra, J.; Keshavkant, S. Nanotechnology: An Efficient Approach for Rejuvenation of Aged Seeds. Physiol. Mol. Biol. Plants 2021, 27, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Ciacka, K.; Krasuska, U.; Staszek, P.; Wal, A.; Zak, J.; Gniazdowska, A. Effect of Nitrogen Reactive Compounds on Aging in Seed. Front. Plant Sci. 2020, 11, 1011. [Google Scholar] [CrossRef] [PubMed]
- Ciacka, K.; Staszek, P.; Sobczynska, K.; Krasuska, U.; Gniazdowska, A. Nitric Oxide in Seed Biology. Int. J. Mol. Sci. 2022, 23, 14951. [Google Scholar] [CrossRef] [PubMed]
- Elena Gonzalez-Benito, M.; Kremer, C.; Ibanez, M.A.; Martin, C. Effect of Antioxidants on the Genetic Stability of Cryopreserved Mint Shoot Tips by Encapsulation-Dehydration. Plant Cell Tissue Organ Cult. 2016, 127, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Uchendu, E.E.; Leonard, S.W.; Traber, M.G.; Reed, B.M. Vitamins C and E Improve Regrowth and Reduce Lipid Peroxidation of Blackberry Shoot Tips Following Cryopreservation. Plant Cell Rep. 2009, 29, 25. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Liu, T.; Zhang, D. Exogenous Dehydrin NnRab18 Improves the Arabidopsis Cryopreservation by Affecting ROS Metabolism and Protecting Antioxidase Activities. In Vitro Cell. Dev. Biol.-Plant 2022, 58, 530–539. [Google Scholar] [CrossRef]
- Manuchehrabadi, N.; Gao, Z.; Zhang, J.; Ring, H.L.; Shao, Q.; Liu, F.; McDermott, M.; Fok, A.; Rabin, Y.; Brockbank, K.G.M.; et al. Improved Tissue Cryopreservation Using Inductive Heating of Magnetic Nanoparticles. Sci. Transl. Med. 2017, 9, eaah4586. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, N.M.; Reis, R.L.; Mano, J.F. The Potential of Liquid Marbles for Biomedical Applications: A Critical Review. Adv. Healthc. Mater. 2017, 6, 1700192. [Google Scholar] [CrossRef] [Green Version]
- Saadeldin, I.M.; Khalil, W.A.; Alharbi, M.G.; Lee, S.H. The Current Trends in Using Nanoparticles, Liposomes, and Exosomes for Semen Cryopreservation. Animals 2020, 10, 2281. [Google Scholar] [CrossRef]
- Ren, L.; Deng, S.; Chu, Y.; Zhang, Y.; Zhao, H.; Chen, H.; Zhang, D. Single-Wall Carbon Nanotubes Improve Cell Survival Rate and Reduce Oxidative Injury in Cryopreservation OfAgapanthus Praecoxembryogenic Callus. Plant Methods 2020, 16, 130. [Google Scholar] [CrossRef]
- Kulus, D.; Tymoszuk, A. Gold Nanoparticles Affect the Cryopreservation Efficiency of in Vitro-Derived Shoot Tips of Bleeding Heart. Plant Cell Tissue Organ Cult. 2021, 146, 297–311. [Google Scholar] [CrossRef]
- Walters, C.; Ballesteros, D.; Vertucci, V.A. Structural Mechanics of Seed Deterioration: Standing the Test of Time. Plant Sci. 2010, 179, 565–573. [Google Scholar] [CrossRef]
- Pence, V.C.; Bruns, E.B.; Meyer, A.; Pritchard, H.W.; Westwood, M.; Linsky, J.; Gratzfeld, J.; Helm-Wallace, S.; Liu, U.; Rivers, M.; et al. Gap Analysis of Exceptional Species—Using a Global List of Exceptional Plants to Expand Strategic Ex Situ Conservation Action beyond Conventional Seed Banking. Biol. Conserv. 2022, 266, 109439. [Google Scholar] [CrossRef]
- Mucha, J.; Szymańska, A.K.; Zadworny, M.; Tylkowski, T.; Michalak, M.; Suszka, J. Effect of Seed Storage Temperature on Fine Root Development and Mycorrhizal Colonization of Young Populus Nigra Seedlings. Ann. For. Sci. 2015, 72, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Pirredda, M.; González-Benito, M.E.; Martín, C.; Mira, S. Genetic and Epigenetic Stability in Rye Seeds under Different Storage Conditions: Ageing and Oxygen Effect. Plants 2020, 9, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plitta-Michalak, B.P.; Ramos, A.A.; Pupel, P.; Michalak, M. Oxidative Damage and DNA Repair in Desiccated Recalcitrant Embryonic Axes of Acer pseudoplatanus L. BMC Plant Biol. 2022, 22, 40. [Google Scholar] [CrossRef]
- Rahman, M.; Liu, L.; Barkla, B.J. A Single Seed Protein Extraction Protocol for Characterizing Brassica Seed Storage Proteins. Agronomy 2021, 11, 107. [Google Scholar] [CrossRef]
- Fleming, M.B.; Richards, C.M.; Walters, C. Decline in RNA Integrity of Dry-Stored Soybean Seeds Correlates with Loss of Germination Potential. J. Exp. Bot. 2017, 68, 2219–2230. [Google Scholar] [CrossRef] [Green Version]
- Puchta, M.; Boczkowska, M.; Groszyk, J. Low RIN Value for RNA-Seq Library Construction from Long-Term Stored Seeds: A Case Study of Barley Seeds. Genes 2020, 11, 1190. [Google Scholar] [CrossRef]
- Saighani, K.; Kondo, D.; Sano, N.; Murata, K.; Yamada, T.; Kanekatsu, M. Correlation between Seed Longevity and RNA Integrity in the Embryos of Rice Seeds. Plant Biotechnol. 2021, 38, 277–283. [Google Scholar] [CrossRef]
- Puchta, M.; Groszyk, J.; Małecka, M.; Koter, M.D.; Niedzielski, M.; Rakoczy-Trojanowska, M.; Boczkowska, M. Barley Seeds MiRNome Stability during Long-Term Storage and Aging. Int. J. Mol. Sci. 2021, 22, 4315. [Google Scholar] [CrossRef] [PubMed]
- Tetreault, H.; Fleming, M.; Hill, L.; Dorr, E.; Yeater, K.; Richards, C.; Walters, C. A Power Analysis for Detecting Aging of Dry-Stored Soybean Seeds: Germination versus RNA Integrity Assessments. Crop Sci. 2022. [Google Scholar] [CrossRef]
- Arif, M.A.R.; Afzal, I.; Börner, A. Genetic Aspects and Molecular Causes of Seed Longevity in Plants—A Review. Plants 2022, 11, 598. [Google Scholar] [CrossRef]
- Ince, A.G.; Karaca, M. Tissue and/or Developmental Stage Specific Methylation of NrDNA in Capsicum Annuum. J. Plant Res. 2021, 134, 841–855. [Google Scholar] [CrossRef]
- Yalamalle, V.; Ithape, D.; Kumar, A.; Bhagat, K.; Ghosh, S.; Singh, M. Seed Treatment with 5-Azacytidine Reduces Ageing-Induced Damage in Onion Seeds. Seed Sci. Technol. 2020, 48, 407–412. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. Seeds and the Art of Genome Maintenance. Front. Plant Sci. 2019, 10, 706. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trusiak, M.; Plitta-Michalak, B.P.; Michalak, M. Choosing the Right Path for the Successful Storage of Seeds. Plants 2023, 12, 72. https://doi.org/10.3390/plants12010072
Trusiak M, Plitta-Michalak BP, Michalak M. Choosing the Right Path for the Successful Storage of Seeds. Plants. 2023; 12(1):72. https://doi.org/10.3390/plants12010072
Chicago/Turabian StyleTrusiak, Magdalena, Beata Patrycja Plitta-Michalak, and Marcin Michalak. 2023. "Choosing the Right Path for the Successful Storage of Seeds" Plants 12, no. 1: 72. https://doi.org/10.3390/plants12010072
APA StyleTrusiak, M., Plitta-Michalak, B. P., & Michalak, M. (2023). Choosing the Right Path for the Successful Storage of Seeds. Plants, 12(1), 72. https://doi.org/10.3390/plants12010072