

Terrestrial and Floating Aquatic Plants Differ in Acclimation to Light Environment
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Acclimation of Photosynthesis and Photoprotection to Light Environment in Land Plants
2.1. Acclimation to Sun-Exposed versus Deeply Shaded Natural Environments
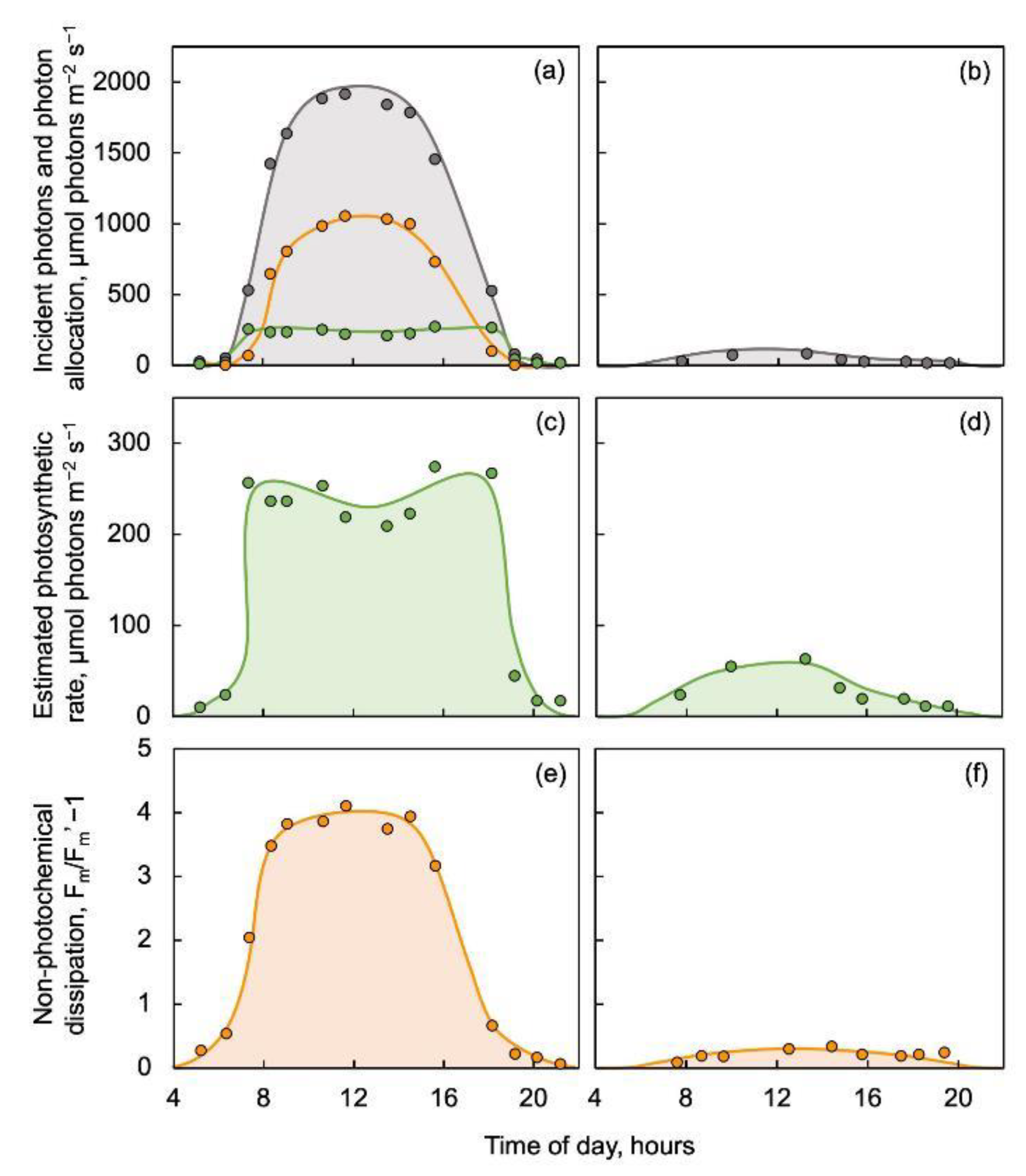
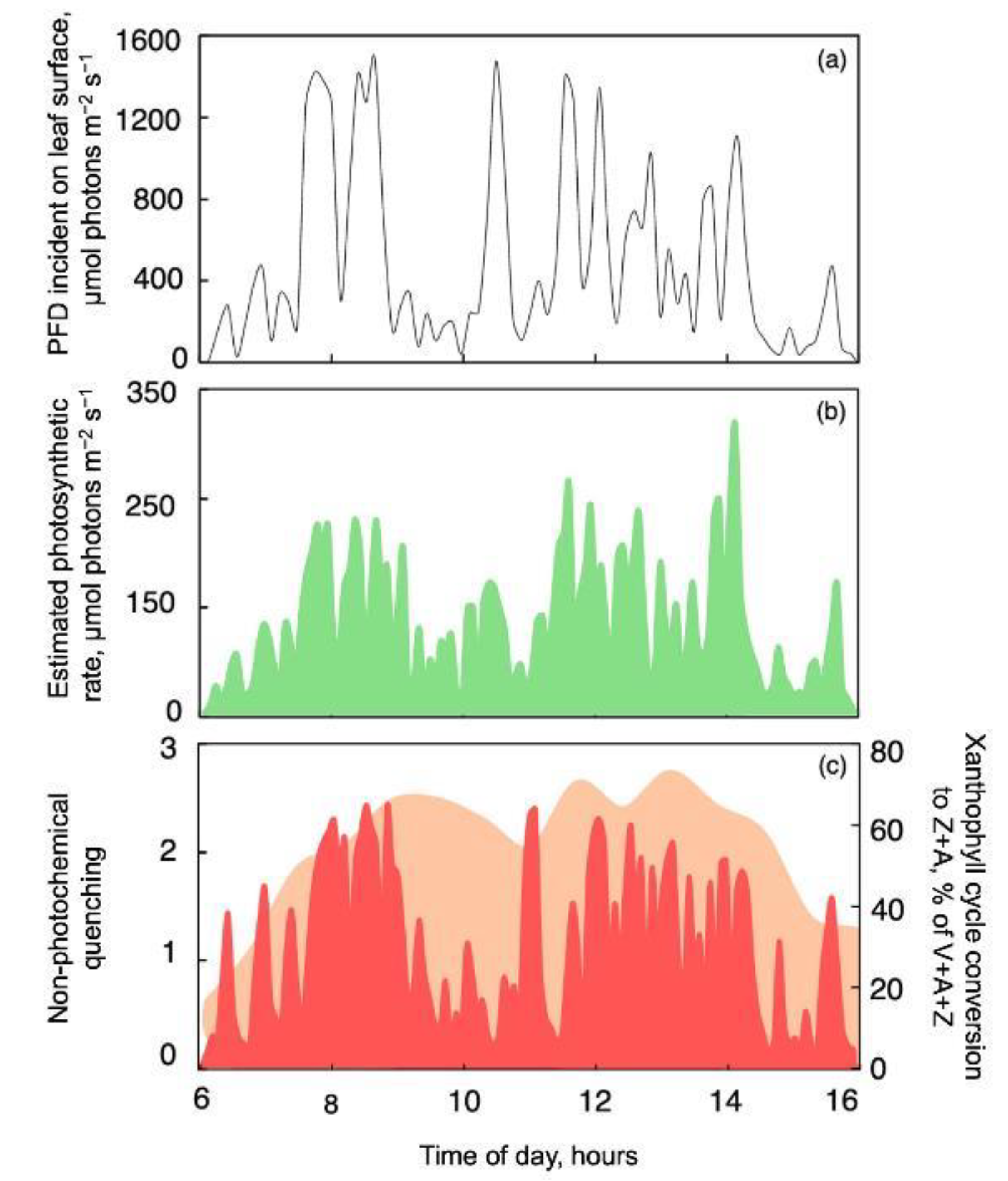
2.2. Acclimation to Shaded Light Environments with Frequent Sunflecks
3. Unusual Features That Afford Flexibility in Lemnaceae
3.1. Rapid Growth
3.2. Constitutively Open Stomates and Facile Gas Flow
3.3. Effective Convective Cooling
3.4. Unique Combination of Annual and Evergreen Terrestrial Plant Features
4. Comparison of Capacity of Photosynthesis and Photosynthesis-Related Processes across Growth Light Intensity in Floating Aquatic Plants versus Land Plants
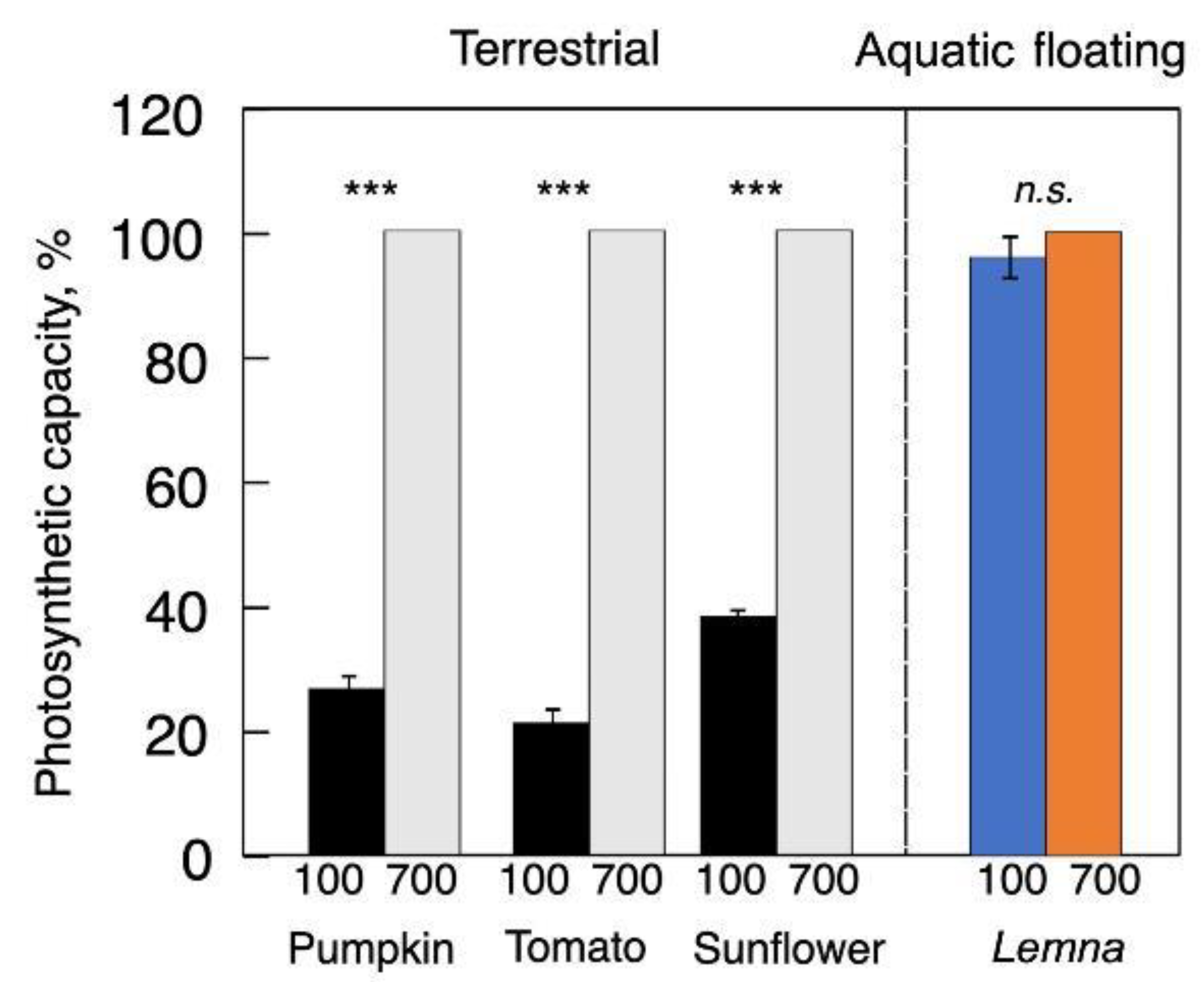
4.1. Maximal Photosynthetic Capacity

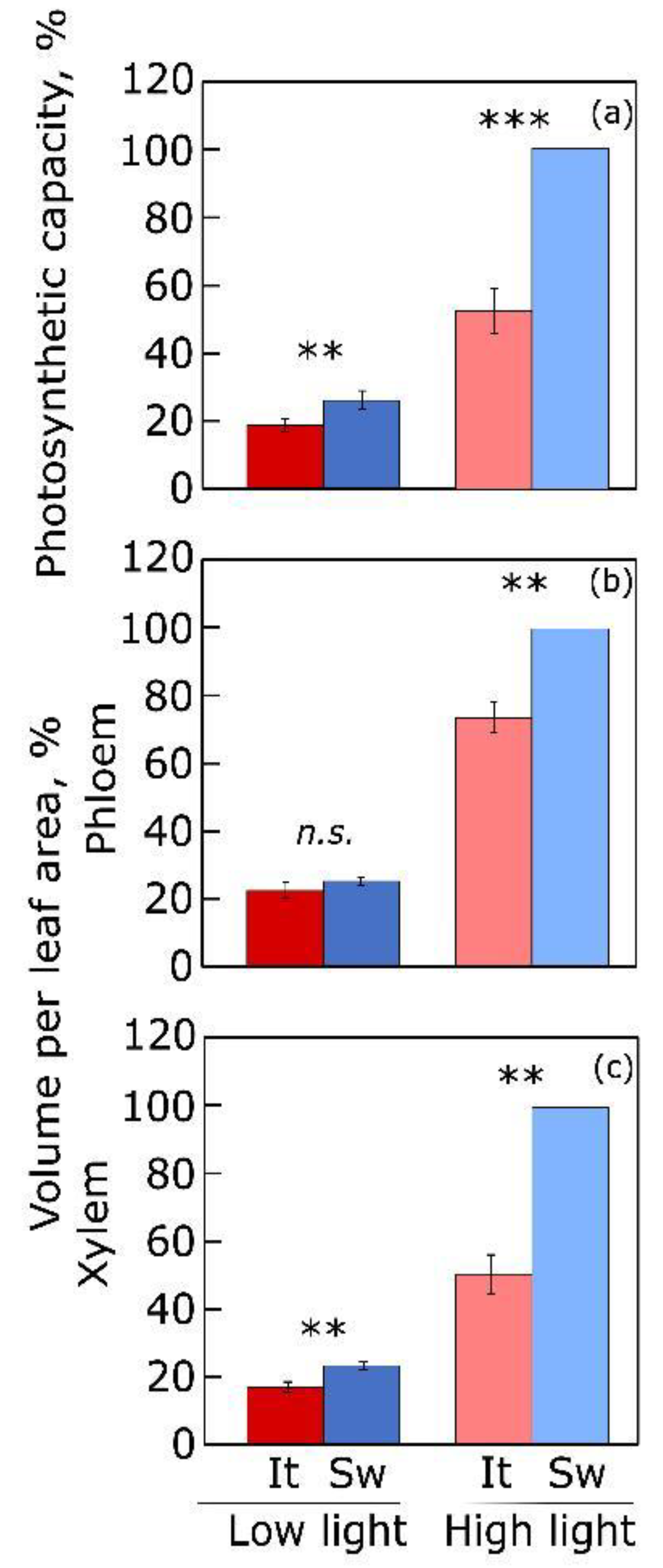
4.2. Vascular Infrastructure for Sugar and Water Transport
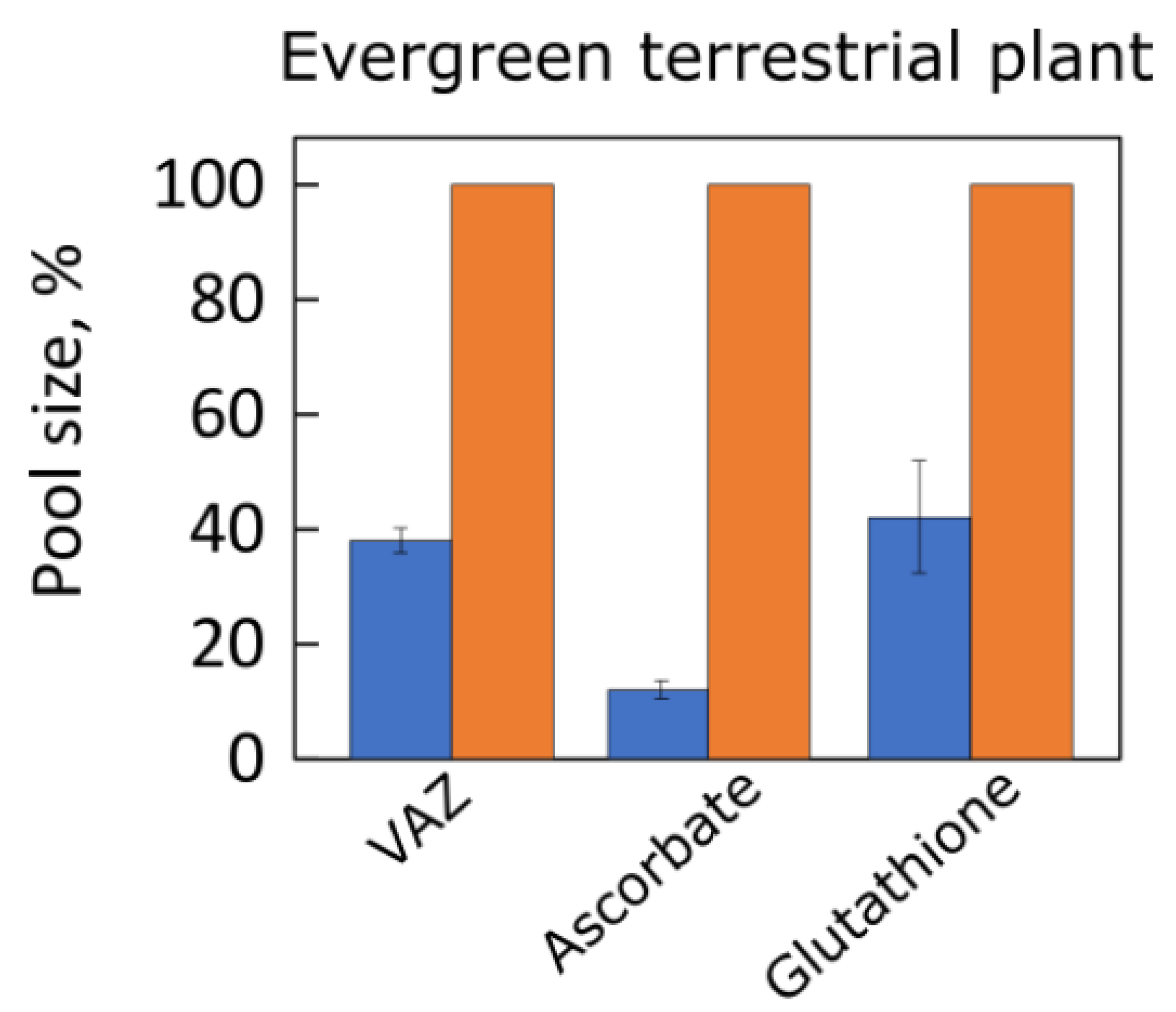
4.3. Antioxidants
5. Comparison of How Quickly Substantial Additional Zeaxanthin Is Formed upon Sudden Transfer from Low to High PFD in Land Plants versus Floating Aquatic Plants
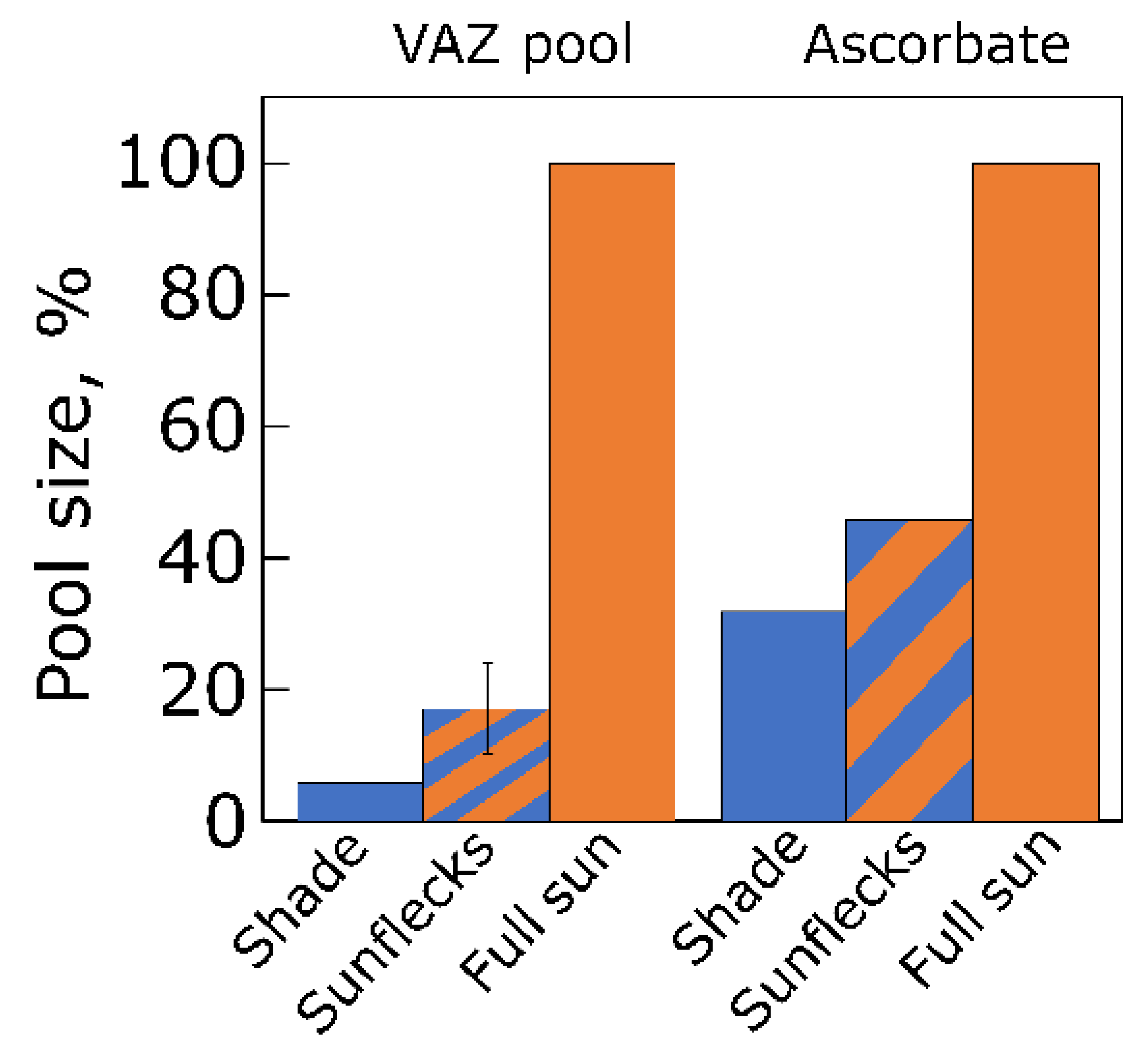
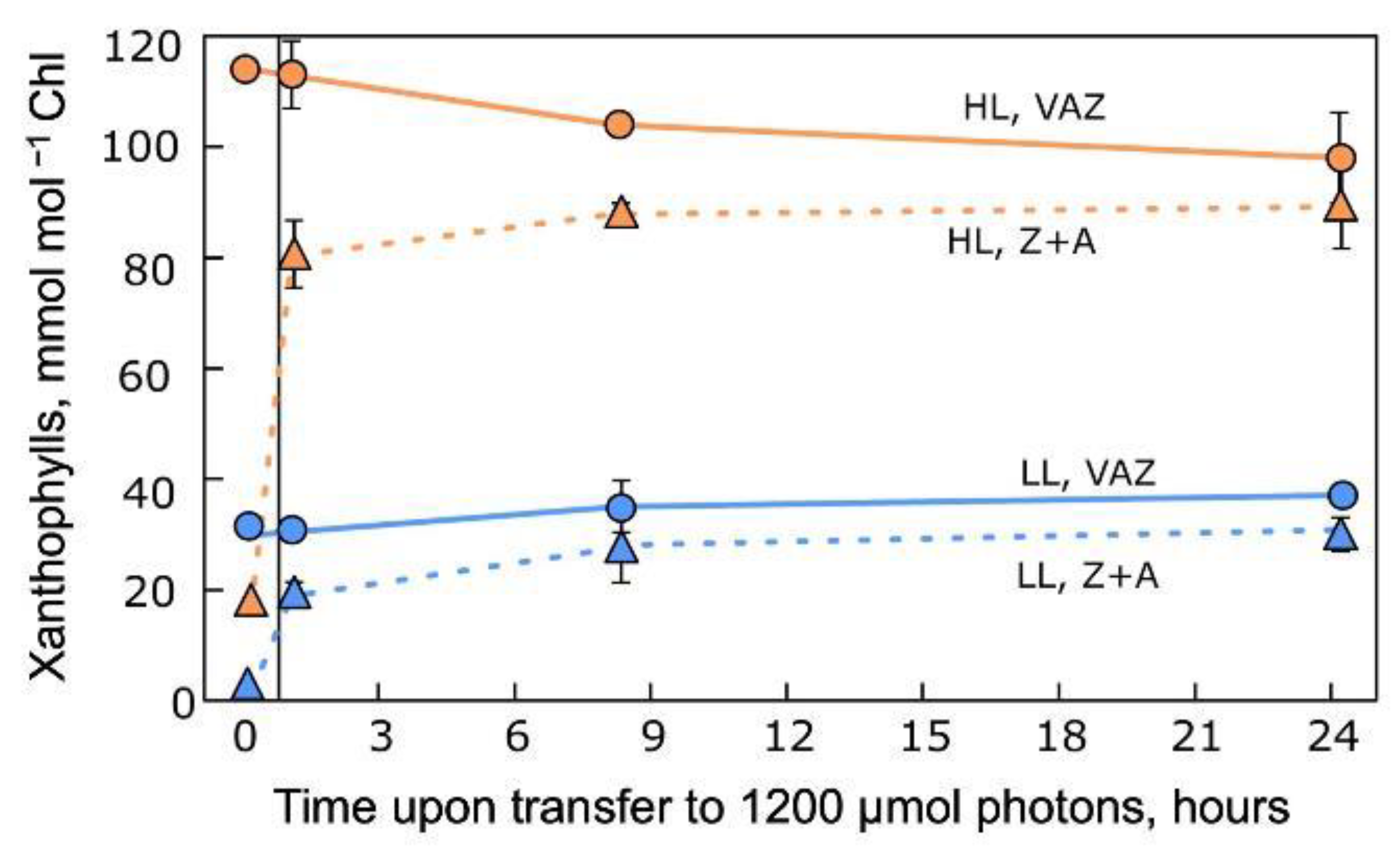
5.1. Changes in Xanthophyll Cycle Pool Size and Conversion in Terrestrial Plants upon Transfer
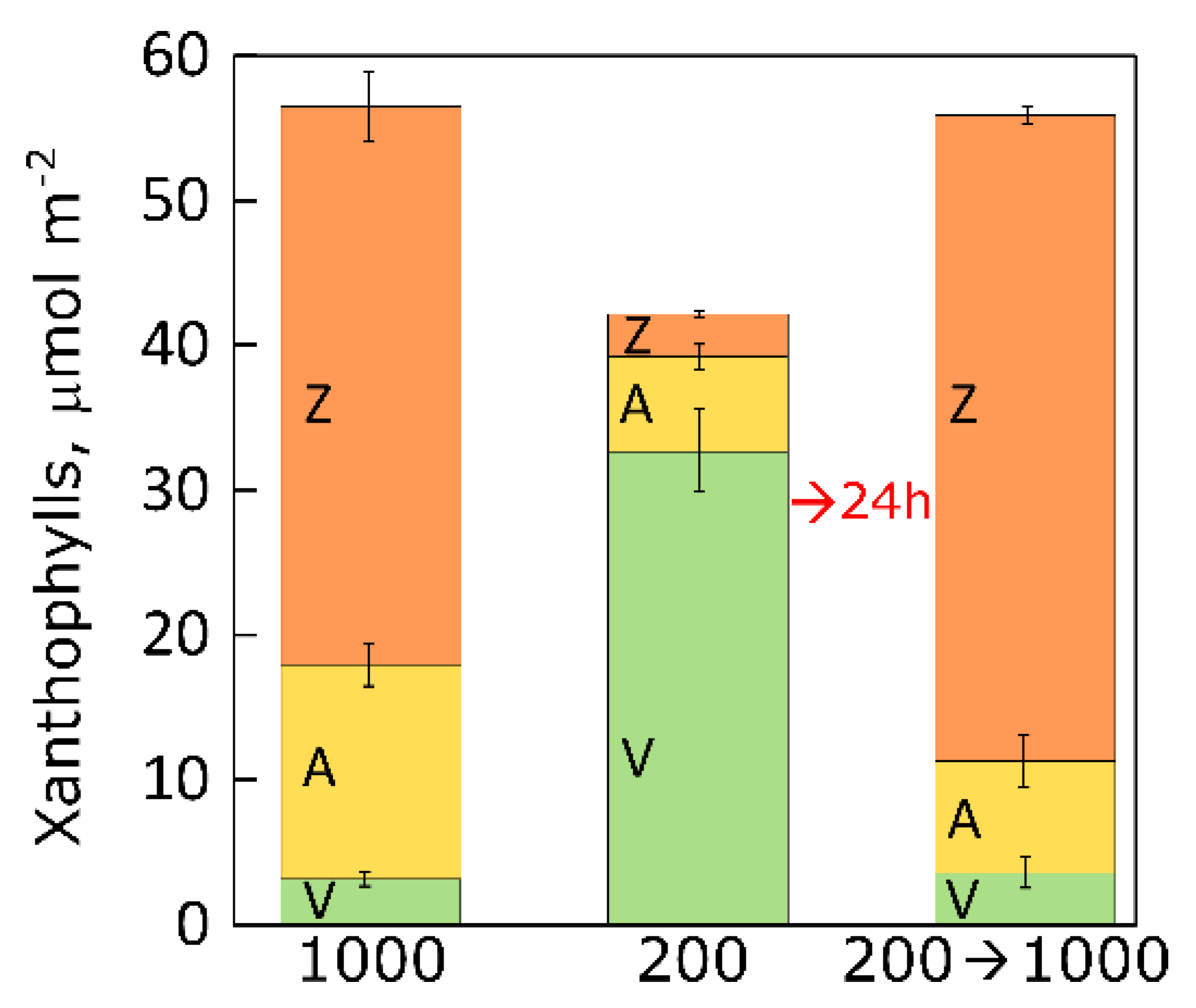
5.2. Changes in Xanthophyll Cycle Pool and Conversion in Lemna upon Transfer
6. Acclimation to Other Environmental Factors in Land Plants and Lemnaceae
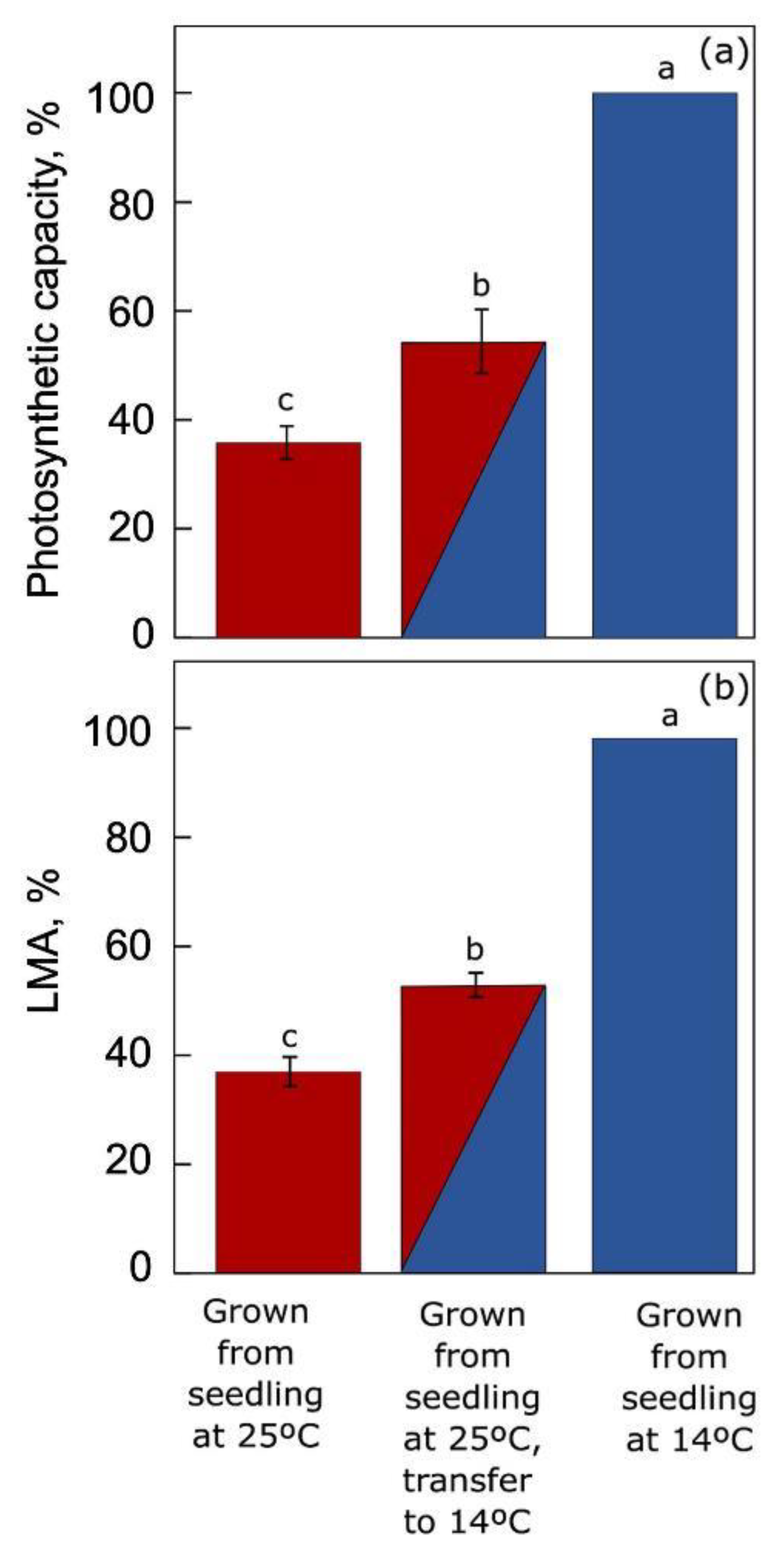
6.1. Temperature Acclimation in Land Plants
6.2. Response to Low or High Nutrient Supply in Lemnaceae
6.3. Maintenance of Light-Harvesting Capacity in Lemnaceae despite Growth Arrest
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Logan, B.A.; Demmig-Adams, B.; Rosenstiel, T.N.; Adams, W.W., III. Effect of nitrogen limitation on foliar antioxidants in relationship to other metabolic characteristics. Planta 1999, 209, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Amiard, V.; Mueh, K.E.; Demmig-Adams, B.; Ebbert, V.; Turgeon, R.; Adams, W.W., III. Anatomical and photosynthetic acclimation to the light environment in species with differing mechanisms of phloem loading. Proc. Natl. Acad. Sci. USA 2005, 102, 12968–12973. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W., III; Stewart, J.J.; Demmig-Adams, B. Photosynthetic modulation in response to plant activity and environment. In The Leaf: A Platform for Performing Photosynthesis; Adams, W.W., III, Terashima, I., Eds.; Advances in Photosynthesis and Respiration; Springer: Cham, Switzerland, 2018; Volume 44, pp. 493–563. [Google Scholar]
- Adams, W.W., III; Stewart, J.J.; Cohu, C.M.; Muller, O.; Demmig-Adams, B. Habitat temperature and precipitation of Arabidopsis thaliana ecotypes determine the response of foliar vasculature, photosynthesis, and transpiration to growth temperature. Front. Plant Sci. 2016, 7, 1026. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Allen, M.T.; Pearcy, R.W. Photosynthetic responses to dynamic light under field conditions in six tropical rainforest shrubs occuring along a light gradient. Oecologia 1997, 111, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W., III; Demmig-Adams, B.; Logan, B.A.; Barker, D.H.; Osmond, C.B. Rapid changes in xanthophyll cycle-dependent energy dissipation and photosystem II efficiency in two vines, Stephania japonica and Smilax australis, growing in the understory of an open Eucalyptus forest. Plant Cell Environ. 1999, 22, 125–136. [Google Scholar] [CrossRef]
- Way, D.A.; Pearcy, R.W. Sunflecks in trees and forests: From photosynthetic physiology to global change biology. Tree Physiol. 2012, 32, 1066–1081. [Google Scholar] [CrossRef]
- Burgess, A.J.; Durand, M.; Gibbs, J.A.; Retkute, R.; Robson, T.M.; Murchie, E.H. The effect of canopy architecture on the patterning of “windflecks” within a wheat canopy. Plant Cell Environ. 2021, 44, 3524–3537. [Google Scholar] [CrossRef]
- Durand, M.; Matule, B.; Burgess, A.J.; Robson, T.M. Sunfleck properties from time series of fluctuating light. Agric. For. Meteorol. 2021, 308, 108554. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.R.M.; Matthews, J.S.A.; McAusland, L.; Blatt, M.R.; Griffiths, H.; Lawson, T. Temporal dynamics of stomatal behavior: Modeling and implications for photosynthesis and water use. Plant Physiol. 2017, 174, 603–613. [Google Scholar] [CrossRef]
- Naidu, S.L.; DeLucia, E.H. Acclimation of shade-developed leaves on saplings exposed to late-season canopy gaps. Tree Physiol. 1997, 17, 367–376. [Google Scholar] [CrossRef]
- Pray, C.L.; Nowlin, W.H.; Vanni, M.J. Deposition and decomposition of periodical cicadas (Homoptera: Cicadidae: Magicicada) in woodland aquatic ecosystems. J. N. Am. Benthol. Soc. 2009, 28, 181–195. [Google Scholar] [CrossRef]
- Weber, M.J.; Brown, M.L. Effects of resource pulses on nutrient availability, ecosystem productivity, and temporal variability following a stochastic disturbance in eutrophic glacial lakes. Hydrobiologia 2016, 771, 165–177. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Peng, W.; Wu, N.; Qu, X.; Yu, Y.; Zhang, Y.; Yang, C. The effects of flood pulse on multiple aquatic organisms in a seasonal shallow lake. Aquat. Ecol. 2021, 55, 379–399. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.; Xiang, L.; Zhang, J.; Xu, Y.; Jeppesen, E. Influence of Lateglacial meltwater influx from alpine glaciers on the aquatic ecosystem of Kanas Lake in the Altai Mountains, China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 604, 111214. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Stewart, J.J.; Adams, W.W., III. Environmental regulation of intrinsic photosynthetic capacity: An integrated view. Curr. Opin. Plant Biol. 2017, 37, 34–41. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Cohu, C.M.; Muller, O.; Adams, W.W., III. Modulation of photosynthetic energy conversion efficiency in nature: From seconds to seasons. Photosynth. Res. 2012, 113, 75–88. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Stewart, J.J.; López-Pozo, M.; Polutchko, S.K.; Adams, W.W., III. Zeaxanthin, a molecule for photoprotection in many different environments. Molecules 2020, 25, 5825. [Google Scholar] [CrossRef]
- Cun, Z.; Xu, X.Z.; Zhang, J.Y.; Shuang, S.P.; Wu, H.M.; An, T.X.; Chen, J.-W. Responses of photosystem to long-term light stress in a typically shade-tolerant species Panax notoginseng. Front. Plant Sci. 2023, 13, 1095726. [Google Scholar] [CrossRef]
- Björkman, O. Responses to Different Quantum Flux Densities. In Physiological Plant Ecology I; Blackwell Science: Oxford, MI, USA, 1981. [Google Scholar]
- Demmig-Adams, B.; Polutchko, S.K.; Stewart, J.J.; Adams, W.W., III. History of excess-light exposure modulates extent and kinetics of fast-acting non-photochemical energy dissipation. Plant Physiol. Rep. 2022, 27, 560–572. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Polutchko, S.K.; Adams, W.W., III. Structure-function-environment relationship of the isomers zeaxanthin and lutein. Photochem 2022, 2, 308–325. [Google Scholar] [CrossRef]
- Parker, G.G.; Fitzjarrald, D.R.; Gonçalves Sampaio, I.C. Consequences of environmental heterogeneity for the photosynthetic light environment of a tropical forest. Agric. For. Meteorol. 2019, 278, 107661. [Google Scholar] [CrossRef]
- Campany, C.E.; Tjoelker, M.G.; von Caemmerer, S.; Duursma, R.A. Coupled response of stomatal and mesophyll conductance to light enhances photosynthesis of shade leaves under sunflecks. Plant Cell Environ. 2016, 39, 2762–2773. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, R.; Koyama, K. Photosynthetic and morphological acclimation to high and low light environments in Petasites japonicus subsp. Giganteus. Forests 2020, 11, 1365. [Google Scholar] [CrossRef]
- Morales, A.; Kaiser, E. Photosynthetic acclimation to fluctuating irradiance in plants. Front. Plant Sci. 2020, 11, 268. [Google Scholar] [CrossRef]
- Landolt, E. Biosystematic investigations in the family of duckweeds (Lemnaceae), II: The family of Lemnaceae: A monographic study 1. Veröffentlichungen des Geobotanischen Institutes der ETH 1986, 95, 638. [Google Scholar]
- Mateo-Elizalde, C.; Lynn, J.; Ernst, E.; Martienssen, R. Duckweeds. Curr. Biol. 2023, 33, 89–91. [Google Scholar] [CrossRef]
- Skillman, J.B.; Strain, B.R.; Osmond, C.B. Contrasting patterns of photosynthetic acclimation and photoinhibition in two evergreen herbs from a winter deciduous forest. Oecologia 1996, 107, 446–455. [Google Scholar] [CrossRef]
- Klich, M.G.; Mújica, M.B.; Fernández, O.A. Stomatal morphology and ontogeny in Spirodela intermedia W. Koch. Aquat. Bot. 1986, 26, 155–164. [Google Scholar] [CrossRef]
- Bauer, R.; Huber, W.; Sankhla, N. Effect of abscisic acid on photosynthesis in Lemna minor L. Zeitschrift für Pflanzenphysiologie 1976, 77, 237–246. [Google Scholar] [CrossRef]
- Smith, J.M.H. The Effect of abscisic acid on growth, photosynthetic rate and carbohydrate metabolism in Lemna minor L. New Phytol. 1976, 76, 11–20. [Google Scholar]
- White, S.L.; Wise, R.R. Anatomy and ultrastructure of Wolffia columbiana and Wolffia borealis, two nonvascular aquatic angiosperms. Int. J. Plant Sci. 1998, 159, 297–304. [Google Scholar] [CrossRef]
- Kwak, M.; Kim, I. Turion as dormant structure in Spirodela polyrhiza. Korean J. Microsc. 2008, 38, 307–314. [Google Scholar]
- Jones, D.H.; Atkinson, B.S.; Ware, A.; Sturrock, C.J.; Bishopp, A.; Wells, D.M. Preparation, scanning and analysis of duckweed using X-ray computed microtomography. Front. Plant Sci. 2021, 11, 617830. [Google Scholar] [CrossRef]
- Borisjuk, N.; Peterson, A.A.; Lv, J.; Qu, G.; Luo, Q.; Shi, L.; Chen, G.; Kishchenko, O.; Zhou, Y.; Shi, J. Structural and biochemical properties of duckweed surface cuticle. Front. Chem. 2018, 6, 317. [Google Scholar] [CrossRef]
- Shrestha, S.; Mahat, J.; Shrestha, J.; Madhav, K.C.; Paudel, K. Influence of high-temperature stress on rice growth and development. A review. Heliyon 2022, 8, e12651. [Google Scholar] [CrossRef]
- Okamoto, A.; Koyama, K.; Bhusal, N. Diurnal change of the photosynthetic light-response curve of Buckbean (Menyanthes trifoliata), an emergent aquatic plant. Plants 2022, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J.; Adams, W.W., III; López-Pozo, M.; Doherty Garcia, N.; McNamara, M.; Escobar, C.M.; Demmig-Adams, B. Features of the duckweed lemna that support rapid growth under extremes of light intensity. Cells 2021, 10, 1481. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; López-Pozo, M.; Polutchko, S.K.; Fourounjian, P.; Stewart, J.J.; Zenir, M.C.; Adams, W.W. III. Growth and nutritional quality of Lemnaceae viewed comparatively in an ecological and evolutionary context. Plants 2022, 11, 145. [Google Scholar] [CrossRef]
- Campany, C.E.; Medlyn, B.E.; Duursma, R.A. Reduced growth due to belowground sink limitation is not fully explained by reduced photosynthesis. Tree Physiol. 2017, 37, 1042–1054. [Google Scholar] [CrossRef]
- Adams, W.W., III; Muller, O.; Cohu, C.M.; Demmig-Adams, B. May photoinhibition be a consequence, rather than a cause, of limited plant productivity? Photosyn. Res. 2013, 117, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Vicente, R.; Bolger, A.M.; Martínez-Carrasco, R.; Pérez, P.; Gutiérrez, E.; Usadel, B.; Morcuende, R. De novo transcriptome analysis of durum wheat flag leaves provides new insights into the regulatory response to elevated CO2 and high temperature. Front. Plant Sci. 2019, 10, 1605. [Google Scholar] [CrossRef] [PubMed]
- Jauregui, I.; Pozueta-Romero, J.; Córdoba, J.; Avice, J.C.; Aparicio-Tejo, P.M.; Baroja-Fernández, E.; Aranjuelo, I. Unraveling the role of transient starch in the response of Arabidopsis to elevated CO2 under long-day conditions. Environ. Exp. Bot. 2018, 155, 158–164. [Google Scholar] [CrossRef]
- Nieuwland, M.; Geerdink, P.; Engelen-Smit, N.P.E.; van der Meer, I.M.; America, A.H.P.; Mes, J.J.; Kootstra, A.M.J.; Henket, J.T.M.M.; Mulder, W.J. Isolation and gelling properties of duckweed protein concentrate. ACS Food Sci. Technol. 2021, 1, 908–916. [Google Scholar] [CrossRef]
- Polutchko, S.K.; Adams, W.W., III; Escobar, C.M.; Demmig-Adams, B. Conquering space with crops that produce ample oxygen and antioxidants. Oxygen 2022, 2, 211–226. [Google Scholar] [CrossRef]
- Stewart, J.J.; Adams, W.W., III; Escobar, C.M.; López-Pozo, M.; Demmig-Adams, B. Growth and essential carotenoid micronutrients in Lemna gibba as a function of growth light intensity. Front. Plant Sci. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Martindale, W.; Bowes, G. The effects of irradiance and CO2 on the activity and activation of ribulose-1,5-bisphosphate carboxylase/oxygenase in the aquatic plant Spirodela polyrhiza. J. Exp. Bot. 1996, 47, 781–784. [Google Scholar] [CrossRef]
- Oguchi, R.; Hikosaka, K.; Hirose, T. Leaf anatomy as a constraint for photosynthetic acclimation: Differential responses in leaf anatomy to increasing growth irradiance among three deciduous trees. Plant Cell Environ. 2005, 28, 916–927. [Google Scholar] [CrossRef]
- Oguchi, R.; Hikosaka, K.; Hirose, T. Does the photosynthetic light-acclimation need change in leaf anatomy? Plant Cell Environ. 2003, 26, 505–512. [Google Scholar] [CrossRef]
- Sims, D.A.; Pearcy, R.W. Response of leaf anatomy and photosynthetic capacity in Alocasia macrorrhiza (Araceae) to a transfer from low to high light. Am. J. Bot. 1992, 79, 449–455. [Google Scholar] [CrossRef]
- Polutchko, S.K.; Stewart, J.J.; Adams, W.W., III; Demmig-Adams, B. Photosynthesis and foliar vascular adjustments to growth light intensity in summer annual species with symplastic and apoplastic phloem loading. J. Plant Physiol. 2021, 267, 153532. [Google Scholar] [CrossRef]
- Adams, W.W., III; Stewart, J.J.; Polutchko, S.K.; Demmig-Adams, B. Leaf vasculature and the upper limit of photosynthesis. In The Leaf: A Platform for Performing Photosynthesis in Advances in Photosynthesis and Respiration; Adams, W.W., III, Terashima, I., Eds.; Springer: Cham, Switzerland, 2018; Volume 44, pp. 27–54. [Google Scholar]
- Cohu, C.M.; Muller, O.; Stewart, J.J.; Demmig-Adams, B.; Adams, W.W., III. Association between minor loading vein architecture and light- and CO2-saturated rates of photosynthetic oxygen evolution among Arabidopsis thaliana ecotypes from different latitudes. Front. Plant Sci. 2013, 22, 264. [Google Scholar] [CrossRef]
- Stewart, J.J.; Polutchko, S.K.; Adams, W.W., III; Demmig-Adams, B. Acclimation of Swedish and Italian ecotypes of Arabidopsis thaliana to light intensity. Photosynth. Res. 2017, 134, 215–229. [Google Scholar] [CrossRef]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.J. How fast can angiosperms grow? Species and clonal diversity of growth rates in the genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 204. [Google Scholar] [CrossRef]
- Cedergreen, N.; Madsen, T.V. Nitrogen uptake by the floating macrophyte Lemna minor. New Phytol. 2002, 155, 285–292. [Google Scholar] [CrossRef]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell. 2021, 33, 3207–3234. [Google Scholar] [CrossRef]
- Kim, I.S. Development of the root system in Spirodela polyrhiza (L.) Schleiden (Lemnaceae). J. Plant Biol. 2007, 50, 540–547. [Google Scholar] [CrossRef]
- Grace, S.C.; Logan, B.A. Acclimation of foliar antioxidant systems to growth irradiance in three broad-leaved evergreen species. Plant Physiol. 1996, 112, 1631–1640. [Google Scholar] [CrossRef]
- Müller-Moulé, P.; Havaux, M.; Niyogi, K.K. Zeaxanthin deficiency enhances the high light sensitivity of an ascorbate-deficient mutant of Arabidopsis. Plant Physiol. 2003, 133, 748–760. [Google Scholar] [CrossRef]
- Neubauer, C.; Yamamoto, H.Y. Membrane barriers and Mehler-peroxidase reaction limit the ascorbate available for violaxanthin de-epoxidase activity in intact chloroplasts. Photosynth. Res. 1994, 39, 137–147. [Google Scholar] [CrossRef]
- Müller-Schüssele, S.J.; Wang, R.; Gütle, D.D.; Romer, J.; Rodriguez-Franco, M.; Scholz, M.; Buchert, F.; Lüth, V.M.; Kopriva, S.; Dörmann, P.; et al. Chloroplasts require glutathione reductase to balance reactive oxygen species and maintain efficient photosynthesis. Plant J. 2020, 103, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Appenroth, K.J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional value of duckweeds (Lemnaceae) as human food. Food Chem. 2017, 217, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Natesh, H.N.; Abbey, L.; Asiedu, S.K. An overview of nutritional and anti nutritional factors in green leafy vegetables. Hortic. Int. J. 2017, 1, 58–65. [Google Scholar] [CrossRef]
- Diotallevi, C.; Angeli, A.; Vrhovsek, U.; Gobbetti, M.; Shai, I.; Lapidot, M.; Tuohy, K. Measuring phenolic compounds in Mankai: A novel polyphenol and amino rich plant protein source. Proc. Nutr. Soc. 2020, 79, E434. [Google Scholar] [CrossRef]
- Yaskolka Meir, A.; Tuohy, K.; von Bergen, M.; Krajmalnik-Brown, R.; Heinig, U.; Zelicha, H.; Tsaban, G.; Rinott, E.; Kaplan, A.; Aharoni, A.; et al. The metabolomic-gut-clinical axis of mankai plant-derived dietary polyphenols. Nutrients 2021, 13, 1866. [Google Scholar] [CrossRef]
- Hu, Z.; Fang, Y.; Yi, Z.; Tian, X.; Li, J.; Jin, Y.; He, K.; Liu, P.; Du, A.; Huang, Y.; et al. Determining the nutritional value and antioxidant capacity of duckweed (Wolffia arrhiza) under artificial conditions. LWT 2022, 153, 112477. [Google Scholar] [CrossRef]
- Pagliuso, D.; Palacios Jara, C.E.; Grandis, A.; Lam, E.; Pena Ferreira, M.J.; Buckeridge, M.S. Flavonoids from duckweeds: Potential applications in the human diet. RSC Adv. 2020, 10, 44981–44988. [Google Scholar] [CrossRef]
- Demmig-Adams, B. Survey of thermal energy dissipation and pigment composition in sun and shade leaves. Plant Cell Physiol. 1998, 39, 474–482. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Winter, K.; Winkelmann, E.; Krüger, A.; Czygan, F.C. Photosynthetic characteristics and the ratios of chlorophyll, β-carotene, and the components of the xanthophyll cycle upon a sudden increase in growth light regime in several plant species. Bot. Acta 1989, 102, 319–325. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Moeller, D.L.; Logan, B.A.; Adams, W.W., III. Positive correlation between levels of retained zeaxanthin + antheraxanthin and degree of photoinhibition in shade leaves of Schefflera arboricola (Hayata) Merrill. Planta 1998, 205, 367–374. [Google Scholar] [CrossRef]
- Brodribb, T.J.; McAdam, S.A.M.; Carins Murphy, M.R. Xylem and stomata, coordinated through time and space. Plant Cell Environ. 2017, 40, 872–880. [Google Scholar] [CrossRef]
- Cohu, C.M.; Lombardi, E.; Adams, W.W., III; Demmig-Adams, B. Increased nutritional quality of plants for long-duration spaceflight missions through choice of plant variety and manipulation of growth conditions. Acta Astronaut. 2014, 94, 799–806. [Google Scholar] [CrossRef]
- Stewart, J.J.; Demmig-Adams, B.; Cohu, C.M.; Wenzl, C.A.; Muller, O.; Adams, W.W., III. Growth temperature impact on leaf form and function in Arabidopsis thaliana ecotypes from northern and southern Europe. Plant Cell Environ. 2016, 39, 1549–1558. [Google Scholar] [CrossRef]
- Duong, T.P.; Tiedje, J.M. Nitrogen fixation by naturally occurring duckweed–cyanobacterial associations. Can. J. Microbiol. 1985, 31, 327–330. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Polutchko, S.K.; Stewart, J.J.; Mcnamara, M.; Garcia, N.D.; López-Pozo, M.; Adams, W.W., III; Demmig-Adams, B. Lemna as a sustainable, highly nutritious crop: Nutrient production in different light environments. Nutraceuticals 2022, 2, 350–364. [Google Scholar] [CrossRef]
- Roubeau Dumont, E.; Larue, C.; Pujol, B.; Lamaze, T.; Elger, A. Environmental variations mediate duckweed (Lemna minor L.) sensitivity to copper exposure through phenotypic plasticity. Environ. Sci. Pollut. Res. 2019, 26, 14106–14115. [Google Scholar] [CrossRef]
- Harper, J.E. Soil and symbiotic nitrogen requirements for optimum soybean production. Crop. Sci. 1974, 14, 255–260. [Google Scholar] [CrossRef]
- Driever, S.M.; Van Nes, E.H.; Roijackers, R.M.M. Growth limitation of Lemna minor due to high plant density. Aquat. Bot. 2005, 81, 245–251. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 4 | 7 | 10 | 14 | |
|---|---|---|---|---|---|
| Photosynthesis per area (µmol O2 m−2 s−1) | 40.3 ± 5.6 | 59.8 ± 10.8 | 71.1 ± 4.3 | 82.9 ± 7.4 | 100.4 ± 13.5 |
| Dry mass (g m−2) | 26.0 ± 1.1 | 39.1 ± 2.0 | 50.4 ± 2.0 | 62.5 ± 2.7 | 58.7 ± 2.5 |
| Photosynthesis per dry mass (µmol O2 g−1 s−1) | 1.6 ± 0.2 | 1.4 ± 0.1 | 1.4 ± 0.1 | 1.3 ± 0.1 | 1.7 ± 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Pozo, M.; Adams, W.W., III; Polutchko, S.K.; Demmig-Adams, B. Terrestrial and Floating Aquatic Plants Differ in Acclimation to Light Environment. Plants 2023, 12, 1928. https://doi.org/10.3390/plants12101928
López-Pozo M, Adams WW III, Polutchko SK, Demmig-Adams B. Terrestrial and Floating Aquatic Plants Differ in Acclimation to Light Environment. Plants. 2023; 12(10):1928. https://doi.org/10.3390/plants12101928
Chicago/Turabian StyleLópez-Pozo, Marina, William W. Adams, III, Stephanie K. Polutchko, and Barbara Demmig-Adams. 2023. "Terrestrial and Floating Aquatic Plants Differ in Acclimation to Light Environment" Plants 12, no. 10: 1928. https://doi.org/10.3390/plants12101928
APA StyleLópez-Pozo, M., Adams, W. W., III, Polutchko, S. K., & Demmig-Adams, B. (2023). Terrestrial and Floating Aquatic Plants Differ in Acclimation to Light Environment. Plants, 12(10), 1928. https://doi.org/10.3390/plants12101928