

Arnica montana L.: Doesn’t Origin Matter? †


Abstract
:1. Introduction
2. Botanical and Phytochemical Evidence
2.1. Botany of A. montana and Possible Segregation in Two Subspecies
2.2. Chemistry of A. montana and Existence of Two Chemotypes
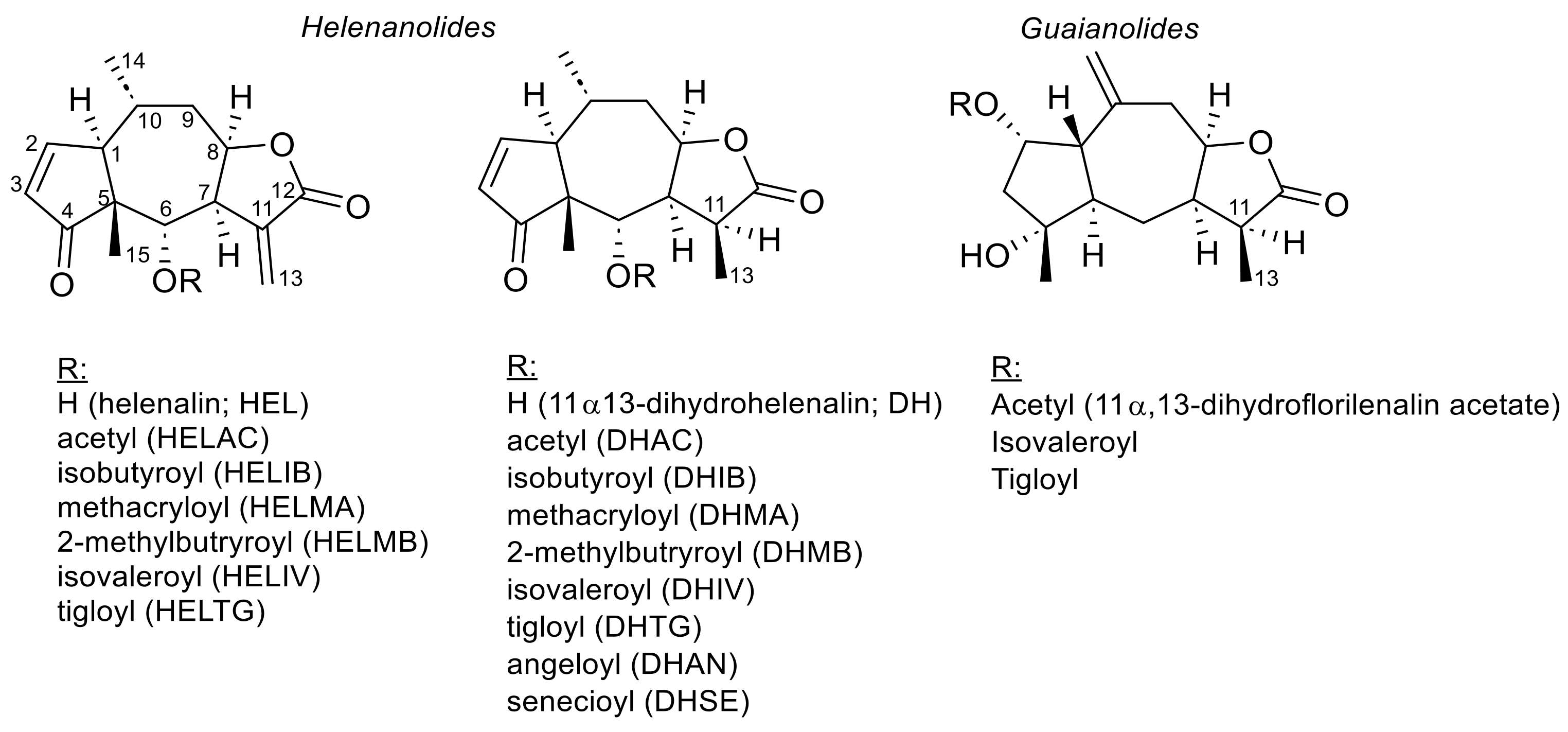
2.2.1. Chemical Constituents
2.2.2. Quantitative Analysis of STLs
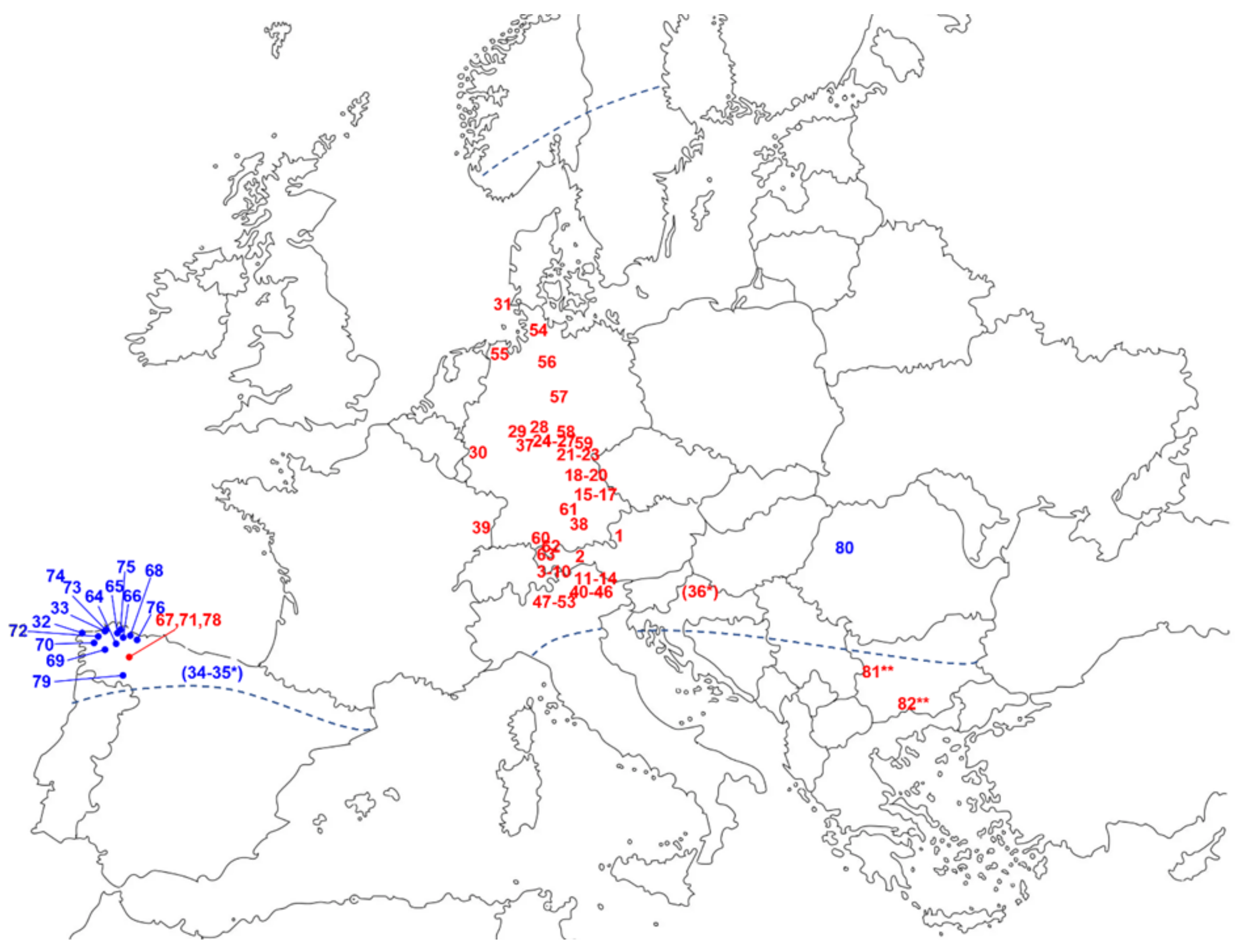
2.2.3. Different STL Chemotypes of A. montana Occur in Distinct Regions of Europe


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Region | Country | Location | Total STL (%) | HEL-Der. (% of Total) | DH-Der. (% of Total) | RHD | Ref. |
|---|---|---|---|---|---|---|---|---|
| 1 | Alps | AT | Lungau | 0.7 | 84 | 16 | 5.3 | [25] |
| 2 | Alps | AT | Ötztal | 0.2 | 88 | 12 | 7.3 | [25] |
| 3 | Alps | AT | Silvretta, Fimbertal | 0.3 | 84 | 16 | 5.3 | [25] |
| 4 | Alps | CH | Engadin, Alpe Laretz | 0.5 | 51 | 49 | 1.0 | [25] |
| 5 | Alps | CH | Engadin, Tuoi-Hütte | 0.6 | 52 | 48 | 1.1 | [25] |
| 6 | Alps | CH | Engadin, Jantal-Hütte | 0.7 | 59 | 41 | 1.4 | [25] |
| 7 | Alps | CH | Engadin, Wiesbdadener Hütte | 0.7 | 57 | 43 | 1.3 | [25] |
| 8 | Alps | CH | Engadin, Ftan Pitschen | 0.3 | 74 | 26 | 2.8 | [25] |
| 9 | Alps | CH | Engadin, Scuol | 0.5 | 72 | 28 | 2.6 | [25] |
| 10 | Alps | CH | Engadin, Prada da Tuoi | 0.4 | 78 | 22 | 3.5 | [25] |
| 11 | Alps | IT | Regensburger Hütte | 0.5 | 84 | 16 | 5.3 | [25] |
| 12 | Alps | IT | Wolkenstein Campinoi | 0.6 | 88 | 12 | 7.3 | [25] |
| 13 | Alps | IT | Ref. Fermada | 0.6 | 91 | 9 | 10.1 | [25] |
| 14 | Alps | IT | Grödner Joch | 0.5 | 93 | 7 | 13.3 | [25] |
| 15 | Bayer. Wald | DE | Haidmühle | 0.6 | 88 | 12 | 7.3 | [25] |
| 16 | Bayer. Wald | DE | Hinterfirmiansreuth | 0.4 | 83 | 17 | 4.9 | [25] |
| 17 | Bayer. Wald | DE | St. Engelmar | 0.2 | 87 | 13 | 6.7 | [25] |
| 18 | Oberpf. Wald | DE | Gibacht b. Furth i.W. | 0.5 | 81 | 19 | 4.3 | [25] |
| 19 | Oberpf. Wald | DE | Schönau | 0.3 | 85 | 15 | 5.7 | [25] |
| 20 | Oberpf. Wald | DE | Silberhütte | 0.3 | 69 | 31 | 2.2 | [25] |
| 21 | Fichtelgeb. | DE | Kirchlamitz | 0.6 | 83 | 17 | 4.9 | [25] |
| 22 | Fichtelgeb. | DE | Weißenstadt | 0.7 | 79 | 21 | 3.8 | [25] |
| 23 | Fichtelgeb. | DE | Tröstan | 0.4 | 76 | 24 | 3.2 | [25] |
| 24 | Rhön | DE | Wüstensachsen | 0.6 | 80 | 20 | 4.0 | [25] |
| 25 | Rhön | DE | Wasserkuppe | 0.4 | 77 | 23 | 3.3 | [25] |
| 26 | Rhön | DE | Grabenhöfchen | 0.5 | 72 | 28 | 2.6 | [25] |
| 27 | Rhön | DE | Heidelstein | 0.3 | 76 | 24 | 3.2 | [25] |
| 28 | Meißner | DE | Meißnerhaus | 0.4 | 82 | 18 | 4.6 | [25] |
| 29 | Westerwald | DE | Rennerod | 0.2 | 87 | 13 | 6.7 | [25] |
| 30 | Eifel | DE | Baasern | 0.4 | 82 | 18 | 4.6 | [25] |
| 31 | North Sea Island | DE | Sylt | 0.3 | 91 | 9 | 10.1 | [25] |
| 32 | Galicia | ES | Berdoias/Muxia | 0.6 | 12 | 88 | 0.1 | [25] |
| 33 | Commercial * | ES | Herborista Mordage, La Coruna, Spanien | 0.4 | 15 | 85 | 0.2 | [25] |
| 34 | Commercial * | ES | Caelo GmbH, Hilden; (origin Spain) 1986 | 0.7 | 0 | 100 | 0.0 | [25] |
| 35 | Commercial * | ES | Caelo GmbH, Hilden; (origin Spain) 1990 | 1.0 | 2 | 98 | 0.0 | [25] |
| 36 | Commercial * | (YU) | Caelo GmbH, Hilden; (origin: former Yugoslavia) | 0.6 | 76 | 24 | 3.2 | [25] |
| 37 | Cultivated | DE | Exp. Station Rauischholzhausen | 0.4 | 85 | 15 | 5.7 | [25] |
| 38 | Cultivated | DE | Bayer. Landesanstalt f. Bodenkultur und Pflanzenbau, Freising | 0.9 | 78 | 22 | 3.5 | [25] |
| 39 | Cultivated | FR | Agricultural research institute Colmar | 0.9 | 80 | 20 | 4.0 | [25] |
| 40 | Alps | IT | Baito Casere Vece | 1.2 | 76 | 24 | 3.2 | [31] |
| 41 | Alps | IT | Malga Caserine di Dentro | 1.3 | 65 | 35 | 1.9 | [31] |
| 42 | Alps | IT | Malga Casina | 0.9 | 80 | 21 | 3.9 | [31] |
| 43 | Alps | IT | Malga Fregasoga | 2.3 | 71 | 29 | 2.4 | [31] |
| 44 | Alps | IT | Malga Juribello | 1.0 | 84 | 16 | 5.2 | [31] |
| 45 | Alps | IT | Malga Ora | 1.1 | 55 | 45 | 1.2 | [31] |
| 46 | Alps | IT | Malga Sass | 0.5 | 71 | 29 | 2.5 | [31] |
| 47 | Alps | IT | Malga Vericolo | 0.8 | 67 | 34 | 2.0 | [31] |
| 48 | Alps | IT | Monte Bondone | 0.7 | 63 | 38 | 1.7 | [31] |
| 49 | Alps | IT | Monte Peller | 0.9 | 72 | 28 | 2.5 | [31] |
| 50 | Alps | IT | Passo Campelli | 0.8 | 67 | 33 | 2.0 | [31] |
| 51 | Alps | IT | Passo Manghen | 1.8 | 68 | 32 | 2.1 | [31] |
| 52 | Alps | IT | Rifugio Bedole | 1.2 | 66 | 34 | 1.9 | [31] |
| 53 | Alps | IT | Rifugio Camini | 1.5 | 88 | 12 | 7.3 | [31] |
| 54 | Schleswig-Holstein | DE | Aukrug | 0.9 | n.a. | n.a. | 8.1 | [32] |
| 55 | Lower Saxony | DE | Tergast | 0.6 | n.a. | n.a. | 4.7 | [32] |
| 56 | Lower Saxony | DE | Unterlüß | 0.8 | n.a. | n.a. | 5.8 | [32] |
| 57 | Lower Saxony | DE | Braunlage | 0.7 | n.a. | n.a. | 8.9 | [32] |
| 58 | Thuringia | DE | Vesser | 0.7 | n.a. | n.a. | 10.6 | [32] |
| 59 | Bavaria | DE | Teuschnitz | 0.6 | n.a. | n.a. | 8.5 | [32] |
| 60 | Bavaria | DE | Immenstadt | 1.1 | n.a. | n.a. | 2.8 | [32] |
| 61 | Bavaria | DE | Schnellers | 1.1 | n.a. | n.a. | 2.5 | [32] |
| 62 | Bavaria | DE | Sonthofen | 0.9 | n.a. | n.a. | 3.9 | [32] |
| 63 | Bavaria | DE | Fellhorn | 0.7 | n.a. | n.a. | 4.7 | [32] |
| 64 | Galicia | ES | A Balsa | 1.8 | 10 | 90 | 0.1 | [35] |
| 65 | Galicia | ES | Aborbó | 1.5 | 11 | 89 | 0.1 | [35] |
| 66 | Galicia | ES | Aldixe | 1.6 | 4 | 96 | 0.0 | [35] |
| 67 | Galicia | ES | Alto do Couto | 1.4 | 72 | 28 | 2.6 | [35] |
| 68 | Galicia | ES | Campo do Oso | 1.8 | 10 | 90 | 0.1 | [35] |
| 69 | Galicia | ES | Chaos | 1.4 | 21 | 79 | 0.3 | [35] |
| 70 | Galicia | ES | Cruz da Golada | 1.2 | 6 | 94 | 0.1 | [35] |
| 71 | Galicia | ES | Formigueiros | 1.2 | 57 | 43 | 1.3 | [35] |
| 72 | Galicia | ES | Meira | 1.7 | 9 | 91 | 0.1 | [35] |
| 73 | Galicia | ES | Penchaina | 1.9 | 5 | 95 | 0.1 | [35] |
| 74 | Galicia | ES | Ponte Pedrido | 1.7 | 4 | 96 | 0.0 | [35] |
| 75 | Galicia | ES | Ponte Xestido | 1.5 | 12 | 88 | 0.1 | [35] |
| 76 | Galicia | ES | Reibocha | 1.7 | 4 | 96 | 0.0 | [35] |
| 77 | Galicia | ES | Sto Tomé | 1.5 | 25 | 75 | 0.3 | [35] |
| 78 | Galicia | ES | Sufoio | 1.1 | 48 | 52 | 0.9 a | [35] |
| 79 | Galicia | ES | Valdin | 1.3 | 7 | 93 | 0.1 | [35] |
| 80a | Carpathians | RO | Apuseni mountains/silic | 0.8 | 26 | 74 | 0.4 | [34] |
| 80b | Carpathians | RO | Apuseni mountains/calc | 1.2 | 29 | 71 | 0.4 | [34] |
| 81a | Cultivated ** | BG/DE | Rhodopes mt.; German seeds | 0.2 | 72 | 28 | 2.6 | [36] |
| 81b | Cultivated ** | BG/UA | Rhodopes mt.; Ukrainian seeds | 0.3 | 75 | 25 | 3.0 | [36] |
| 81c | Cultivated ** | BG/AT | Rhodopes mt.; Austrian seeds | 0.4 | 76 | 24 | 3.2 | [36] |
| 82a | Cultivated ** | BG/DE | Vitosha mt.; German seeds | 0.4 | 82 | 18 | 4.4 | [36] |
| 82b | Cultivated ** | BG/UA | Vitosha mt.; Ukrainian seeds | 0.5 | 77 | 23 | 3.3 | [36] |
2.2.4. The Occurrence of Different STL Chemotypes Is Not Confined to Different Populations
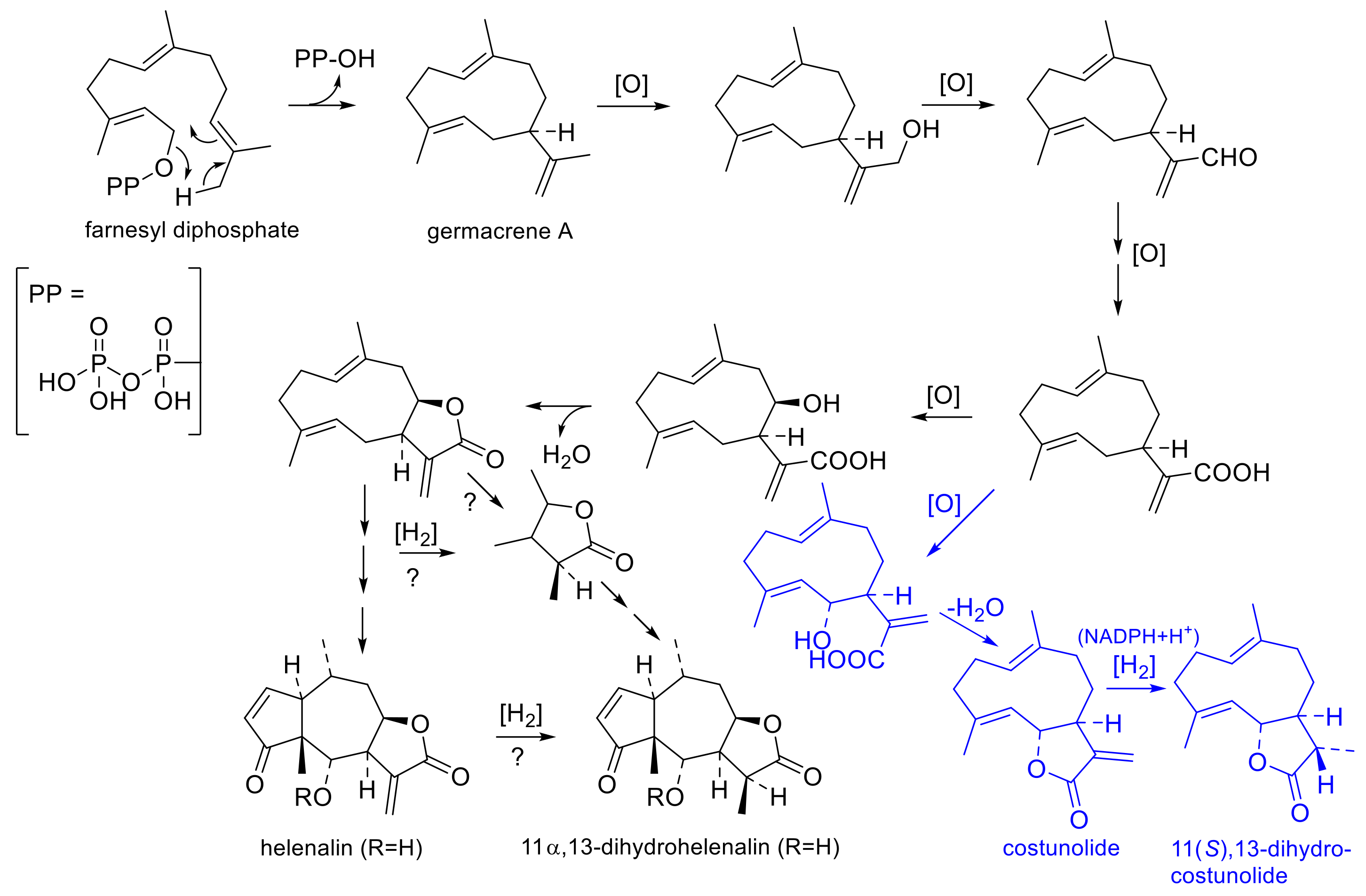
2.2.5. Biogenetic Origin of Different STL Chemotypes
3. Pharmacological Evidence
3.1. Wanted Pharmacological Effect: Anti-Inflammatory Activity—Implications for Drug Efficacy
3.2. Unwanted Effects—Sensitizing and Acute Irritant Potential—Implications on Drug Safety
3.2.1. Contact Allergenic Potential
3.2.2. Acute Skin Irritant Potential
4. Conclusions and Future Directions
5. Materials and Methods
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willuhn, G. Arnica montana L.–Porträt einer Arzneipflanze. Pharm. Ztg. 1991, 136, 2453–2468. [Google Scholar]
- Maguire, B. A monograph of the genus Arnica. Brittonia 1943, 4, 386–510. [Google Scholar]
- Merfort, I. Arnica. In Hager’s Handbuch der Pharmazeutischen Praxis; Hänsel, R., Keller, K., Rimpler, H., Schneider, G., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1992; Volume 4, pp. 342–357. [Google Scholar]
- European Directorate for the Quality of Medicines & HealthCare (EDQM). Arnicae flos. In European Pharmacopoeia, 10th ed.; Council of Europe: Strasbourg, France, 2020. [Google Scholar]
- Dragendorff, G. Die Heilpflanzen der Verschiedenen Völker und Zeiten. Ihre Anwendung, Wesentlichen Bestandteile und Geschichte; Ferdinand Enke Verlag: Stuttgart, Germany, 1888. [Google Scholar]
- Tschirch, A. Handbuch der Pharmakognosie; Verlag Chr. H. Tauchnitz: Leipzig, Germany, 1917; Volume 2, pp. 1174–1184. [Google Scholar]
- Shulha, O. Arnica montana Flower Laboratory Guidance Document; ABC-AHP-NCNPR Botanical Adulterants Prevention Program: Austin, TX, USA, 2022; Available online: https://www.researchgate.net/publication/369033335_Arnica_montana_Flower_Laboratory_Guidance_Document#fullTextFileContent (accessed on 5 September 2023).
- Willuhn, G. Arnicae flos. In Herbal Drugs and Phytopharmaceuticals: A Handbook for Practice on a Scientific Basis, 3rd ed.; Wichtl, M., Ed.; Medpharm: Stuttgart, Germany; CRC Press: Boca Raton, FL, USA, 2004; pp. 54–59. [Google Scholar]
- European Nature Information System. Arnica montana L. Available online: https://eunis.eea.europa.eu/species/Arnica%20montana (accessed on 25 September 2023).
- Von Raison, J.; Heilmann, J.; Merfort, I.; Schmidt, T.J.; Brock, F.E.; Leven, W.; Bomme, U.; Bauer, R. Arnika–Arzneipflanze mit Tradition und Zukunft. Z. Phytother. 2000, 231, 39–54. [Google Scholar]
- European Directorate for the Quality of Medicines & HealthCare (EDQM). Arnicae Tinctura. In European Pharmacopoeia, 10th ed.; Council of Europe: Strasbourg, France, 2020. [Google Scholar]
- Willuhn, G. Arnica flowers: Pharmacology, toxicology, and analysis of the sesquiterpene lactones—Their main active sub-stances. In Phytomedicines of Europe—ACS Symposium Series 691; Lawson, L.D., Bauer, R., Eds.; American Chemical Society: Washington, DC, USA, 1998; Volume 691, pp. 118–132. ISBN 0-8412-3559-7. [Google Scholar]
- Merfort, I. Arnika: Neue Erkenntnisse zum Wirkungsmechanismus einer traditionellen Heilpflanze. Forsch. Komplementarmedizin Klass. Naturheilkunde 2003, 10 (Suppl. S1), 45–48. [Google Scholar] [CrossRef] [PubMed]
- Commission E Monograph 00036 Arnicae flos/Arnikablueten. BAnz Nr. 228. Bundesanzeiger (German Federal Gazette), 5 December 1984. [Google Scholar]
- European Scientific Cooperative on Phytotherapy (ESCOP). Arnicae flos–Arnica Flower. ESCOP Monographs Online Series. 2019. Available online: https://escop.com/arnicae-flos-arnica-flower-escop-2019online-consultation/ (accessed on 5 September 2023).
- European Medicines Agency (EMA). Assessment Report on Arnica montana L., Flos. Available online: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-arnica-montana-l-flos_en.pdf (accessed on 5 September 2023).
- European Medicines Agency (EMA). Community Herbal Monograph on Arnica montana L., Flos. Available online: https://www.ema.europa.eu/en/documents/herbal-monograph/final-community-herbal-monograph-arnica-montana-l-flos_en.pdf (accessed on 5 September 2023).
- Bolos y Vayreda, A. El Arnica montana L. en la Península Ibérica. Farmacogn. An. Inst. Farmacogn. José Celestino Mutis 1947, 4, 145–151. [Google Scholar]
- Romero, R.; Real, C.; Rodríguez-Guitián, M.A.; Barros, M.R.; Rigueiro, A.; González-Hernández, M.P. Estudio de la variabi-lidad biométrica de Arnica montana L. (Asteraceae) en el extremo occidental cantábrico (NW Ibérico). In Proceedings of the IX Colloqui Internacional de Botànica Pirenaico-Cantàbrica, Ordino, Andorra, 7–9 July 2010; pp. 379–388. Available online: https://www.researchgate.net/publication/258256803 (accessed on 5 September 2023).
- Schmiderer, C.; Torres-Londono, P.; Lutz-Röder, A.; Duwe, V.K.; Novak, J. Arnica montana subsp. atlantica: Really a subspecies? Genet. Resour. Crop Evol. 2018, 65, 1829–1838. [Google Scholar] [CrossRef]
- Vera, M.; Romero, R.; Rodríguez-Guitián, M.A.; Barros, R.M.; Real, C.; Bouza, C. Phylogeography and genetic variability of the Arnica montana chemotypes in NW Iberian Peninsula. Silvae Genet. 2015, 63, 293–300. [Google Scholar] [CrossRef]
- Vera, M.; Mora, G.; Rodríguez-Guitián, M.A.; Blanco, A.; Casanova, A.; Real, C.; Romero, R.; Bouza, C. Living at the edge: Population differentiation in endangered Arnica montana from NW Iberian Peninsula. Plant Syst. Evol. 2020, 306, 44. [Google Scholar] [CrossRef]
- Bouza, C.; Lorenzo, I.; Rodríguez-Guitián, M.A.; Casanova, A.; Real, C.; Romero, R.; Vera, M. Genetic survey extension of the threatened Iberian Arnica montana L. revealed the presence of divergent plastid lineages and highly structured populations in northern Spain. Genet. Resour. Crop Evol. 2023, 70, 1677–1689. [Google Scholar] [CrossRef]
- Willuhn, G.; Merfort, I.; Paßreiter, C.M.; Schmidt, T.J. Chemistry and Systematics of the Genus Arnica. In Advances in Compositae Systematics; Hind, D.J.N., Jeffrey, C., Pope, G.V., Eds.; Royal Botanic Gardens: Kew, UK, 1994; pp. 167–195. [Google Scholar]
- Willuhn, G.; Leven, W.; Luley, C. Arnikablüten DAB10–Untersuchungen zur Qualitativen und Quantitativen Variabilität des Sesquiterpenlactongehaltes der Offizinellen Arnikadrogen. Dtsch. Apoth. Ztg. 1994, 134, 4077–4085. [Google Scholar]
- Willuhn, G.; Leven, W. Zur qualitativen und Quantitativen Analyse der Sesquiterpenlactone von Arnikablüten DAB 9. Pharm. Ztg. Wiss. 1991, 136, 32–39. [Google Scholar]
- Kos, O.; Lindenmeyer, M.T.; Tubaro, A.; Sosa, S.; Merfort, I. New Sesquiterpene Lactones from Arnica Tincture Prepared from Fresh Flowerheads of Arnica montana. Planta Med. 2005, 71, 1044–1052. [Google Scholar] [CrossRef]
- Paßreiter, C.; Willuhn, G.; Röder, E. Tussilagine and Isotussilagine: Two Pyrrolizidine Alkaloids in the Genus Arnica. Planta Med. 1992, 58, 556–557. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Paßreiter, C.M.; Wendisch, D.; Willuhn, G. Diterpenes from Arnica angustifolia. Phytochemistry 1995, 40, 1213–1218. [Google Scholar] [CrossRef]
- Schmidt, T.J.; von Raison, J.; Willuhn, G. New Triterpene Esters from Flowerheads of Arnica lonchophylla. Planta Med. 2004, 70, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Clauser, M.; Aiello, N.; Scartezzini, F.; Innocenti, G.; Dall’Acqua, S. Differences in the Chemical Composition of Arnica montana Flowers from Wild Populations of North Italy. Nat. Prod. Commun. 2014, 9, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Seemann, A.; Wallner, T.; Poschlod, P.; Heilmann, J. Variation of Sesquiterpene Lactone Contents in Different Arnica montana Populations: Influence of Ecological Parameters. Planta Med. 2010, 76, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Spitaler, R.; Schlorhaufer, P.D.; Ellmerer, E.P.; Merfort, I.; Bortenschlager, S.; Stuppner, H.; Zidorn, C. Altitudinal variation of secondary metabolite profiles in flowering heads of Arnica montana cv. ARBO. Phytochemistry 2006, 67, 409–417. [Google Scholar] [CrossRef]
- Greinwald, A.; Hartmann, M.; Heilmann, J.; Heinrich, M.; Luick, R.; Reif, A. Soil and Vegetation Drive Sesquiterpene Lactone Content and Profile in Arnica montana L. Flower Heads from Apuseni-Mountains, Romania. Front. Plant Sci. 2022, 13, 813939. [Google Scholar] [CrossRef]
- Perry, N.B.; Burgess, E.J.; Guitián, M.A.R.; Franco, R.R.; Mosquera, E.L.; Smallfield, B.M.; Joyce, N.I.; Littlejohn, R.P. Sesquiterpene Lactones in Arnica montana: Helenalin and Dihydrohelenalin Chemotypes in Spain. Planta Med. 2009, 75, 660–666. [Google Scholar] [CrossRef]
- Todorova, M.; Trendafilova, A.; Vitkova, A.; Petrova, M.; Zayova, E.; Antonova, D. Developmental and Environmental Effects on Sesquiterpene Lactones in Cultivated Arnica montana L. Chem. Biodivers. 2016, 13, 976–981. [Google Scholar] [CrossRef]
- Douglas, J.A.; Smallfield, B.M.; Burgess, E.J.; Perry, N.B.; Anderson, R.E.; Douglas, M.H.; Glennie, V.l.A. Sesquiterpene Lactones in Arnica montana: A Rapid Analytical Method and the Effects of Flower Maturity and Simulated Mechanical Harvesting on Quality and Yield. Planta Med. 2004, 70, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Bomme, U.; Alfermann, A. Sesquiterpene Lactone Content in Leaves of in vitro and Field Cultivated Arnica montana. Planta Med. 1998, 64, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Parafiniuk, A.; Kromer, K.; Fleszar, M.G.; Kreitschitz, A.; Wisniewski, J.; Gamian, A. Localization of Sesquiterpene Lactones Biosynthesis in Flowers of Arnica Taxa. Molecules 2023, 28, 4379. [Google Scholar] [CrossRef] [PubMed]
- Geissman, T. The Biogenesis of Sesquiterpene Lactones of the Compositae. Recent Adv. Phytochem. 1973, 6, 65–95. [Google Scholar] [CrossRef]
- Fischer, N.H. Sesquiterpene Lactones: Biogenesis and Biomimetic Transformations. In Biochemistry of the Mevalonic Acid Pathway to Terpenoids; Towers, G.H.N., Stafford, H.A., Eds.; Plenum Press: New York, NY, USA, 1990; Chapter 4; pp. 161–201. [Google Scholar] [CrossRef]
- De Kraker, J.-W.; Franssen, M.C.; Joerink, M.; de Groot, A.; Bouwmeester, H.J. Biosynthesis of Costunolide, Dihydrocostunolide, and Leucodin. Demonstration of Cytochrome P450-Catalyzed Formation of the Lactone Ring Present in Sesquiterpene Lactones of Chicory. Plant Physiol. 2002, 129, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Willuhn, G. Arnika-Kontaktdermatitis und die sie Verursachenden Kontaktallergene. Dtsch. Apoth. Ztg. 1986, 126, 2038–2044. [Google Scholar]
- Hall, I.; Starnes, C.O., Jr.; Lee, K.; Waddell, T. Mode of action of sesquiterpene lactones as anti-inflammatory agents. J. Pharm. Sci. 1980, 69, 537–543. [Google Scholar] [CrossRef]
- Hall, I.; Lee, K.; Starnes, C.; Sumida, Y.; Wu, R.; Waddell, T.; Cochran, J.; Gerhart, K. Anti-Inflammatory Activity of Sesquiterpene Lactones and Related Compounds. J. Pharm. Sci. 1979, 68, 537–542. [Google Scholar] [CrossRef]
- Picman, A.K. Biological Activities of Sesquiterpene Lactones. Biochem. Syst. Ecol. 1986, 14, 255–281. [Google Scholar] [CrossRef]
- Schmidt, T.J. Toxic Activities of Sesquiterpene Lactones: Structural and Biochemical Aspects. Curr. Org. Chem. 1999, 3, 577–605. [Google Scholar]
- Schmidt, T.J. Structure-activity relationships of sesquiterpene lactones. In Studies in Natural Products Chemistry; Rahman, A.-U., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 33, pp. 309–392. [Google Scholar] [CrossRef]
- Lyß, G.; Schmidt, T.J.; Merfort, I.; Pahl, H.L. Helenalin, an Anti-Inflammatory Sesquiterpene Lactone from Arnica, Selectively Inhibits Transcription Factor NF-κB. Biol. Chem. 1997, 378, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Lyß, G.; Knorre, A.; Schmidt, T.J.; Pahl, H.L.; Merfort, I. The Anti-inflammatory Sesquiterpene Lactone Helenalin Inhibits the Transcription Factor NF-κB by Directly Targeting p65. J. Biol. Chem. 1998, 273, 33508–33516. [Google Scholar] [CrossRef] [PubMed]
- Lyß, G.; Schmidt, T.J.; Pahl, H.L.; Merfort, I. Studies for the anti-inflammatory activity of Arnica tincture using the transcription factor NF-κB as molecular target. Pharm. Pharmacol. Lett. 1999, 9, 5–8. [Google Scholar]
- Rüngeler, P.; Castro, V.; Mora, G.; Gören, N.; Vichnewski, W.; Pahl, H.L.; Merfort, I.; Schmidt, T.J. Inhibition of transcription factor NF-κB by sesquiterpene lactones: A proposed molecular mechanism of action. Bioorg. Med. Chem. 1999, 7, 2343–2352. [Google Scholar] [CrossRef] [PubMed]
- García-Piñeres, A.J.; Castro, V.; Mora, G.; Schmidt, T.J.; Strunck, E.; Pahl, H.L.; Merfort, I. Cysteine 38 in p65/NF-κB Plays a Crucial Role in DNA Binding Inhibition by Sesquiterpene Lactones. J. Biol. Chem. 2001, 276, 39713–39720. [Google Scholar] [CrossRef] [PubMed]
- Klaas, C.A.; Wagner, G.; Laufer, S.; Sosa, S.; Della Loggia, R.; Bomme, U.; Pahl, H.L.; Merfort, I. Studies on the Anti-Inflammatory Activity of Phytopharmaceuticals Prepared from Arnica Flowers. Planta Med. 2002, 68, 385–391. [Google Scholar] [CrossRef]
- Berges, C.; Fuchs, D.; Opelz, G.; Daniel, V.; Naujokat, C. Helenalin suppresses essential immune functions of activated CD4+ T cells by multiple mechanisms. Mol. Immunol. 2009, 46, 2892–2901. [Google Scholar] [CrossRef]
- Jäger, C.; Hrenn, A.; Zwingmann, J.; Suter, A.; Merfort, I. Phytomedicines Prepared from Arnica Flowers Inhibit the Transcription Factors AP-1 and NF-κB and Modulate the Activity of MMP1 and MMP13 in Human and Bovine Chondrocytes. Planta Med. 2009, 75, 1319–1325. [Google Scholar] [CrossRef]
- Tornhamre, S.; Schmidt, T.J.; Näsman-Glaser, B.; Ericsson, I.; Lindgren, J. Inhibitory effects of helenalin and related compounds on 5-lipoxygenase and leukotriene C4 synthase in human blood cells. Biochem. Pharmacol. 2001, 62, 903–911. [Google Scholar] [CrossRef]
- Schmidt, T.J.; Klempnauer, K.-H. Natural Products with Antitumor Potential Targeting the MYB-C/EBPβ-p300 Transcription Module. Molecules 2022, 27, 2077. [Google Scholar] [CrossRef] [PubMed]
- Bujnicki, T.; Wilczek, C.; Schomburg, C.; Feldmann, F.; Schlenke, P.; Müller-Tidow, C.; Schmidt, T.J.; Klempnauer, K.-H. Inhibition of Myb-dependent gene expression by the sesquiterpene lactone mexicanin-I. Leukemia 2012, 26, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, C.; Schuehly, W.; Da Costa, F.B.; Klempnauer, K.-H.; Schmidt, T.J. Natural sesquiterpene lactones as inhibitors of Myb-dependent gene expression: Structure–activity relationships. Eur. J. Med. Chem. 2013, 63, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Jakobs, A.; Uttarkar, S.; Schomburg, C.; Steinmann, S.; Coulibaly, A.; Schlenke, P.; Berdel, W.E.; Müller-Tidow, C.; Schmidt, T.J.; Klempnauer, K.-H. An isoform-specific C/EBPβ inhibitor targets acute myeloid leukemia cells. Leukemia 2016, 30, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Jakobs, A.; Steinmann, S.; Henrich, S.M.; Schmidt, T.J.; Klempnauer, K.-H. Helenalin Acetate, a Natural Sesquiterpene Lactone with Anti-inflammatory and Anti-cancer Activity, Disrupts the Cooperation of CCAAT Box/Enhancer-binding Protein β (C/EBPβ) and Co-activator p300. J. Biol. Chem. 2016, 291, 26098–26108. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Isshiki, H.; Sugita, T.; Tanabe, O.; Kinoshita, S.; Nishio, Y.; Nakajima, T.; Hirano, T.; Kishimoto, T. A nuclear factor for IL-6 expression (NF-IL6) is a member of a C/EBP family. EMBO J. 1990, 9, 1897–1906. [Google Scholar] [CrossRef]
- Le Coz, J.; Ducombs, G. Plants and plant products. In Contact Dermatitis, 4th ed.; Frosch, P.J., Menné, T., Lepottevin, J.-P., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 751–800. [Google Scholar]
- Warshaw, E.M.; Zug, K.A. Sesquiterpene lactone allergy. Am. J. Contact Dermat. 1996, 7, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Hausen, B.M.; Vieluf, I.K. Allergiepflanzen Handbuch und Atlas, 2nd ed.; Nikol-Verlag: Hamburg, Germany, 1997; pp. 74–77. [Google Scholar]
- Jacob, M.; Brinkmann, J.; Schmidt, T.J. Sesquiterpene lactone mix as a diagnostic tool for Asteraceae allergic contact dermatitis: Chemical explanation for its poor performance and Sesquiterpene lactone mix II as a proposed improvement. Contact Dermat. 2012, 66, 233–240. [Google Scholar] [CrossRef]
- Schmidt, T.J. Helenanolide-type sesquiterpene lactones—III. Rates and stereochemistry in the reaction of helenalin and related helenanolides with sulfhydryl containing biomolecules. Bioorg. Med. Chem. 1997, 5, 645–653. [Google Scholar] [CrossRef]
- Hausen, B.M.; Schmalle, H.W. Structure-activity aspects of 4 allergenic sesquiterpene lactones tacking the exocyclic α-methylene at the lactone ring. Contact Dermat. 1985, 13, 329–332. [Google Scholar] [CrossRef]
- Lass, C.; Vocanson, M.; Wagner, S.; Schempp, C.M.; Nicolas, J.-F.; Merfort, I.; Martin, S.F. Anti-inflammatory and immune-regulatory mechanisms prevent contact hypersensitivity to Arnica montana L. Exp. Dermatol. 2008, 17, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Lass, C.; Merfort, I.; Martin, S.F. In vitro and in vivo analysis of pro- and anti-inflammatory effects of weak and strong contact allergens. Exp. Dermatol. 2010, 19, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Jocher, A.; Nist, G.; Weiss, J.M.; Wetzel, D.; Merfort, I.; Jakob, T.; Schempp, C.M. Allergenic potential of Arnica-containing formulations in Arnica-allergic patients. Contact Dermat. 2009, 61, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Passreiter, C.M.; Florack, M.; Willuhn, G.; Goerz, G. Allergic contact dermatitis from Asteraceae. Identification of an 8,9-epoxythymol diester as a contact allergen in Arnica sachalinensis. Dermatosen Beruf Umw. 1988, 36, 79–82. [Google Scholar]
- Elissalde, M.H.; Ivie, G.W.; Rowe, L.D.; Elissalde, G.S. Considerations of the structure of sesquiterpene lactones on biological activity: Influence of the α-methylene-γ-lactone moiety on mast cell degranulation. Am. J. Vet. Res. 1983, 44, 1894–1897. [Google Scholar] [PubMed]
- Wu, J.; Chun, Y.; Ebizuka, Y.; Sankawa, U. Biologically active constituents of Centipeda minima: Isolation of a new plenolin ester and the antiallergy activity of sesquiterpene lactones. Chem. Pharm. Bull. 1985, 33, 4091–4094. [Google Scholar] [CrossRef]
- Wu, J.-B.; Chun, Y.-T.; Ebizuka, Y.; Sankawa, U. Biologically Active Constituents of Centipedaminima: Sesquiterpenes of Potential Anti-allergy Activity. Chem. Pharm. Bull. 1991, 39, 3272–3275. [Google Scholar] [CrossRef]
- Jürgens, F.M.; Robledo, S.M.; Schmidt, T.J. Evaluation of Pharmacokinetic and Toxicological Parameters of Arnica Tincture after Dermal Application In Vivo. Pharmaceutics 2022, 14, 2379. [Google Scholar] [CrossRef]
- Jürgens, F.M.; Herrmann, F.C.; Robledo, S.M.; Schmidt, T.J. Dermal Absorption of Sesquiterpene Lactones from Arnica Tincture. Pharmaceutics 2022, 14, 742. [Google Scholar] [CrossRef]
- PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 27 July 2023).
- SciFinder. Available online: https://scifinder-n.cas.org/ (accessed on 27 July 2023).



| STL | Kf [26] | 1/Kf | SCF [4,11] |
|---|---|---|---|
| DH | 0.80 | 1.250 | |
| DHAC | 0.85 | 1.176 | |
| DHTG 1 | 0.88 | 1.136 | 1.187 |
| DHIV | 0.87 | 1.149 | |
| HEL | 2.20 | 0.455 | |
| HELAC | 1.23 | 0.813 | |
| HELTG 1 | 1.15 | 0.870 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, T.J. Arnica montana L.: Doesn’t Origin Matter? Plants 2023, 12, 3532. https://doi.org/10.3390/plants12203532
Schmidt TJ. Arnica montana L.: Doesn’t Origin Matter? Plants. 2023; 12(20):3532. https://doi.org/10.3390/plants12203532
Chicago/Turabian StyleSchmidt, Thomas J. 2023. "Arnica montana L.: Doesn’t Origin Matter?" Plants 12, no. 20: 3532. https://doi.org/10.3390/plants12203532
APA StyleSchmidt, T. J. (2023). Arnica montana L.: Doesn’t Origin Matter? Plants, 12(20), 3532. https://doi.org/10.3390/plants12203532