

Genome-Wide Characterization of Tomato FAD Gene Family and Expression Analysis under Abiotic Stresses
 ,
,
Abstract
:1. Introduction
2. Result
2.1. Analysis of Physicochemical Properties of SlFAD Gene Family
2.2. Phylogenetic Analysis of the SlFAD Gene Family
2.3. SlFAD Gene Location on Chromosomes and Gene Duplication Analysis
2.4. Analysis of SlFAD Conserved Motifs and Gene Structure
2.5. Analysis of Collinearity Relationships of SlFAD Gene Family
2.6. Analysis of Cis-Acting Elements in SlFAD Promoters
2.7. Expression Analysis of SlFAD Genes in Different Tissues and Fruit Development Stages
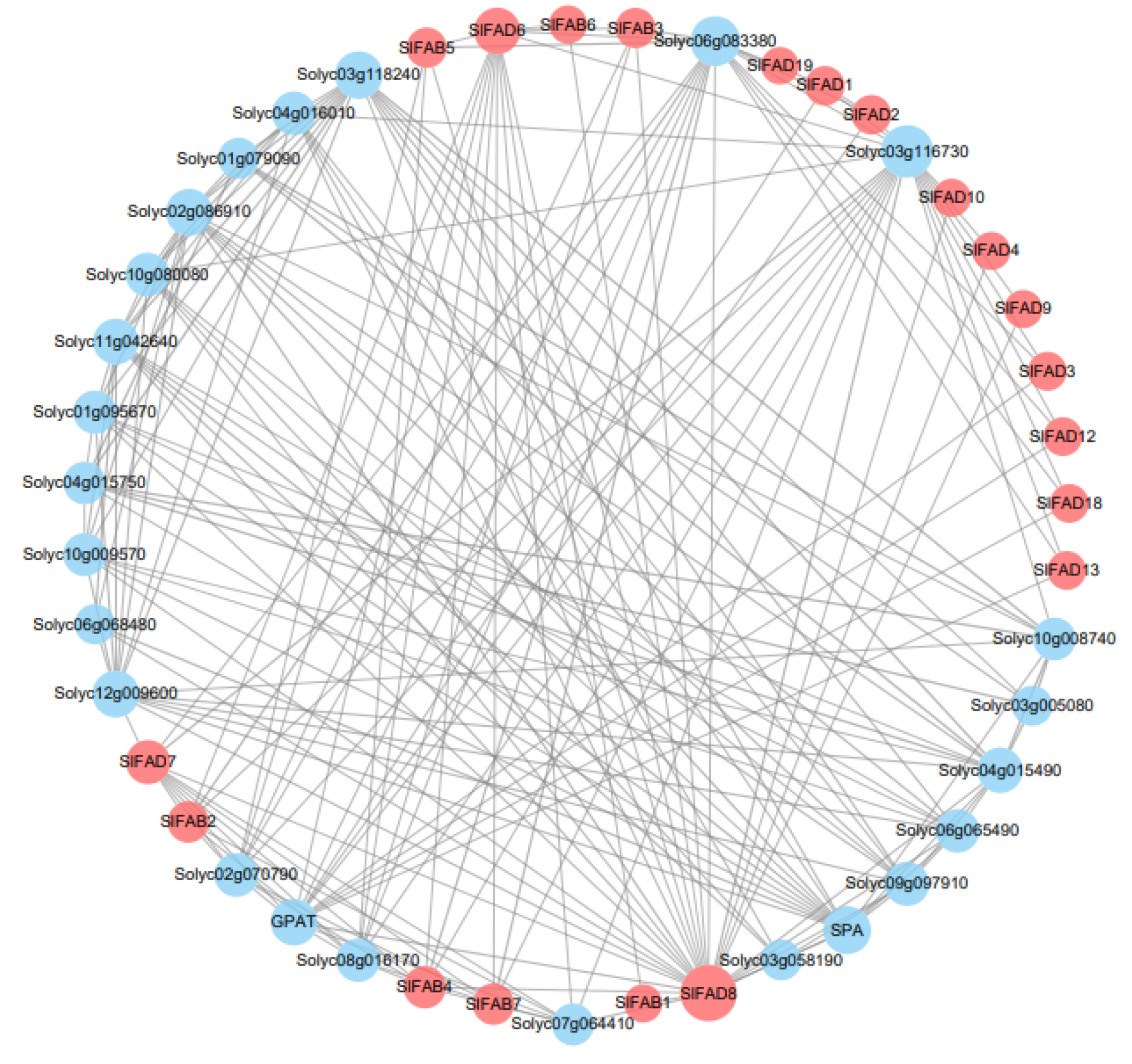
2.8. Protein–Protein Interaction Network Analysis of SlFAD Family Members
2.9. Real-Time Fluorescent Quantitative Analysis of SlFAD Genes
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Data Source
4.3. Identification and Physicochemical Property Analysis of SlFAD Gene Family
4.4. Construction of Phylogenetic Tree and Collinearity Analysis of SlFAD Family Genes
4.5. Analysis of Conserved Motifs, Gene Structure, and Chromosome Location of SlFAD
4.6. Analysis of Cis-Acting Elements, Interaction Networks, and Expression of SlFAD
4.7. Expression Analysis of Tomato FAD Family Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aejaz, A.D.; Abhikshit, R.C.; Pavan, K.K.; Pavan, K.K.; Neelakantan, A. The FAD2 gene in plants: Occurrence, regulation, and role. Front. Plant Sci. 2017, 8, 1789. [Google Scholar]
- Zhang, Z.-S.; Wei, X.-Y.; Liu, W.-X.; Min, X.-Y.; Jin, X.-Y.; Boniface, N.; Wang, Y.-R. Genome-wide identification and expression analysis of the fatty acid desaturase genes in Medicago truncatula. Biochem. Biophys. Res. Commun. 2018, 499, 361–367. [Google Scholar] [CrossRef]
- Kim, H.; Carlos, R.-N.; Rahul, K.K.; Payal, K.; Ivan, P.; James, B.; Ralf, K.; Ye, J. Unsaturated fatty acids stimulate tumor growth through stabilization of β-catenin. Cell Rep. 2015, 13, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.-S.; Zhu, Z.-Y.; Wang, H.-R.; Wang, L.; Cheng, L.-Z.; Yuan, Y.-J.; Zheng, Y.-S.; Li, D.-D. Characterization and functional analysis of a plastidial FAD6 gene and its promoter in the mesocarp of oil palm (Elaeis guineensis). Sci. Hortic. 2019, 239, 163–170. [Google Scholar] [CrossRef]
- Fatemeh, D.N.; Kazem, Y. Bioinformatics study of delta-12 fatty acid desaturase 2 (FAD2) gene in oilseeds. Mol. Biol. Rep. 2014, 41, 5077–5087. [Google Scholar]
- Shanklin, J.; Cahoon, E.B. Desaturation and related modifications of fatty acids 1. Annu. Rev. Plant Biol. 1998, 49, 611–641. [Google Scholar] [CrossRef]
- Dong, C.-J.; Cao, N.; Zhang, Z.-G.; Shang, Q.-M. Characterization of the fatty acid desaturase genes in cucumber: Structure, phylogeny, and expression patterns. PLoS ONE 2016, 11, e149917. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.-D.; Xu, W.-Z.; Shu, Q.-Y.; Li, S.-S.; Wu, Q.; Feng, C.-Y.; Gu, Z.-Y.; Wang, L.-S. Fatty acid desaturase 3 (PsFAD3) from Paeonia suffruticosa reveals high α-linolenic acid accumulation. Plant Sci. 2018, 274, 212–222. [Google Scholar] [CrossRef]
- He, M.; Qin, C.-X.; Wang, X.; Ding, N.-Z. Plant unsaturated fatty acids: Biosynthesis and regulation. Front. Plant Sci. 2020, 11, 390. [Google Scholar] [CrossRef]
- Xue, Y.-F.; Chai, C.-Y.; Chen, B.-J.; Shi, X.-F.; Wang, B.-T.; Mei, F.-R.; Jiang, M.-L.; Liao, X.-L.; Yang, X.; Yuan, C.-L.; et al. Whole-genome mining and in silico analysis of FAD gene family in Brassica juncea. J. Plant Biochem. Biotechnol. 2020, 29, 149–154. [Google Scholar] [CrossRef]
- Xue, Y.-F.; Chen, B.-J.; Wang, R.; Aung, N.W.; Li, J.-N.; Chai, Y.-R. Genome-wide survey and characterization of fatty acid desaturase gene family in Brassica napus and its parental species. Appl. Biochem. Biotech. 2018, 184, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Rashmi, S.; Sanjeev, K. Genome-wide identification, characterization and in-silico profiling of genes encoding FAD (fatty acid desaturase) proteins in chickpea (Cicer arietinum L.). Plant Gene 2019, 18, 100180. [Google Scholar]
- Cheng, C.-Z.; Liu, F.; Sun, X.-L.; Wang, B.; Liu, J.-P.; Ni, X.-T.; Hu, C.-H.; Deng, G.-M.; Tong, Z.; Zhang, Y.-Y.; et al. Genome-wide identification of FAD gene family and their contributions to the temperature stresses and mutualistic and parasitic fungi colonization responses in banana. Int. J. Biol. Macromol. 2022, 204, 661–676. [Google Scholar] [CrossRef]
- Wang, X.; Yu, C.; Liu, Y.; Yang, L.; Li, Y.; Yao, W.; Cai, Y.-C.; Yan, X.; Li, S.-B.; Cai, Y.-H.; et al. GmFAD3A, a ω-3 fatty acid desaturase gene, enhances cold tolerance and seed germination rate under low temperature in rice. Int. J. Mol. Sci. 2019, 20, 3796. [Google Scholar] [CrossRef] [PubMed]
- Zahra, H.; Amin, A.; Wei, H.; Sun, W.-B.; Ruan, H.-H.; Qiang, Z.-G.; Ali, M. Identification, evolution, expression, and docking studies of fatty acid desaturase genes in wheat (Triticum aestivum L.). BMC Genom. 2020, 10, 21. [Google Scholar]
- Ángela, R.; Andreu, V.; Hernández, M.L.; Lagunas, B.; Picorel, R.; Martínez, J.M.; Alfonso, M. Contribution of the different omega-3 fatty acid desaturase genes to the cold response in soybean. Exp. Bot. 2012, 63, 4973–4982. [Google Scholar]
- Peng, Z.-Y.; Ruan, J.; Tian, H.-Y.; Shan, L.; Meng, J.; Guo, F.; Zhang, Z.-M.; Ding, H.; Wan, S.; Li, X.-G. The family of peanut fatty acid desaturase genes and a functional analysis of four ω-3 AhFad3 members. Plant Mol. Biol. Rep. 2020, 38, 209–221. [Google Scholar] [CrossRef]
- Cheng, C.-Z.; Li, D.; Qi, Q.; Sun, X.-L.; Mensah, R.A.; Bodjrenou, M.D.; Zhang, Y.-Y.; Hao, X.-Y.; Zhang, Z.-H.; Lai, Z.-X. The root endophytic fungus serendipita indica improves resistance of banana to Fusarium oxysporum f. sp. cubense tropical race 4. Eur. J. Plant Pathol. 2020, 156, 87–100. [Google Scholar] [CrossRef]
- Jiao, S.-Q.; Zhou, J.-M.; Shang, Y.-Q.; Wang, J.-X.; Zhang, A.-J.; He, H.-B.; Zhao, Q.-Z.; Li, Y.; Yao, D. Cloning and genetic transformation of soybean fatty acid dehydrogenase GmFAD3C-1 gene. Chin. J. Oil Crop Sci. 2022, 44, 1006–1017. [Google Scholar]
- Zhang, M.; Barg, R.; Yin, M.-A.; Yardena, G.-D.; Alicia, L.-F.; Yehiam, S.; Sara, S.; Gozal, B.-H. Modulated fatty acid desaturation via overexpression of two distinct ω-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef]
- Chi, X.-Y.; Yang, Q.-L.; Lu, Y.-D.; Wang, J.-Y.; Zhang, Q.-F.; Pan, L.-J.; Chen, M.-N.; He, Y.-N.; Yu, S.-L. Genome-wide analysis of fatty acid desaturases in soybean (Glycine max). Plant Mol. Biol. Rep. 2011, 29, 769–783. [Google Scholar] [CrossRef]
- Wei, H.; Ali, M.; Xu, S.-Z.; Zhang, Y.-Y.; Liu, G.-Y.; Soheila, A.-D.; Mostafa, G.Z.; Zhu, S.; Yu, C.-M.; Chen, Y.-H.; et al. Genome-wide characterization and expression analysis of fatty acid desaturase gene family in poplar. Int. J. Mol. Sci. 2022, 23, 11109. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.-W.; Zhang, A.-D.; Wu, X.-X.; Zha, D.-S. Identification and bioinformatics analysis of fatty acid desaturase (FAD) gene family in eggplant (Solanum melongena L.). Mol. Plant Breed. 2023, 21, 2453–2463. [Google Scholar]
- Liu, W.; Li, W.; He, Q.-L.; Muhammad, K.D.; Chen, J.-H.; Zhu, S.-J. Characterization of 19 genes encoding membrane-bound fatty acid desaturases and their expression profiles in Gossypium raimondii under low temperature. PLoS ONE 2015, 10, e0123281. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-L.; Qi, C.-Y.; Ma, Y.-H.; Xiong, H.-Y.; Duan, R.-J. Identification and expression analysis of GPAT gene family in hulless barley. Plant Physiol. J. 2022, 58, 2006–2016. [Google Scholar]
- Sascha, L.; Virginie, M.; José, G.; Stephan, W.; Jessika, A.; Seonghoe, J.; Carmen, K.; Helen, B.; George, C.; Ute, H. Arabidopsis SPA proteins regulate photoperiodic flowering and interact with the floral inducer constans to regulate its stability. Development 2006, 133, 3213–3222. [Google Scholar]
- Zhang, Z.-S.; Jin, X.-Y.; Liu, Z.-P.; Zhang, J.-Y.; Liu, W.-X. Genome-wide identification of FAD gene family and functional analysis of MsFAD3.1 involved in the accumulation of-linolenic acid in alfalfa. Crop Sci. 2020, 61, 566–579. [Google Scholar] [CrossRef]
- Deborah, A.J.; Michael, A.T. The monosaccharide transporter gene family in Arabidopsis and rice: A history of duplications, adaptive evolution, and functional divergence. Mol. Biol. Evol. 2012, 24, 2412–2423. [Google Scholar]
- Steven, B.C.; Arvind, M.; Andrew, B.; Nevin, D.Y.; Georgiana, M. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar]
- Cao, J.; Shi, F. Evolution of the RALF gene family in plants: Gene duplication and selection patterns. Evol. Bioinform. 2012, 8, 271–292. [Google Scholar] [CrossRef]
- Xu, L.; Zeng, W.-J.; Li, J.-J.; Liu, H.; Yan, G.-J.; Si, P.; Yang, C.; Shi, Y.; He, Q.-L.; Zhou, W.-J. Characteristics of membrane-bound fatty acid desaturase (FAD) genes in Brassica napus L. and their expressions under different cadmium and salinity stresses. Environ. Exp. Bot. 2019, 162, 144–156. [Google Scholar] [CrossRef]
- Li, N.; He, Q.; Wang, J.; Wang, B.-K.; Zhao, J.-T.; Huang, S.-Y.; Yang, T.; Tang, Y.-P.; Yang, S.-B.; Aisimutuola, P.; et al. Super-pangenome analyses highlight genomic diversity and structural variation across wild and cultivated tomato species. Nat. Genet. 2023, 55, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, J.-Y.; Tong, H.-H.; Li, T.-T.; Wang, L.; Chen, H.-Q. Genome-wide analysis of fatty acid desaturase genes in rice (Oryza sativa L.). Sci. Rep. 2019, 9, 19445. [Google Scholar]
- Huang, W.; Xian, Z.-Q.; Kang, X.; Tang, N.; Li, Z.-G. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Huang, J.-J.; Xie, X.-L.; Luo, Z.-Y. Structure and function of the plant PDR-type ABC transporter protein. Chem. Life 2011, 31, 112–119. [Google Scholar]
- Huang, J.-X.; Xue, C.-W.; Wang, H.; Wang, L.-S.; Schmidt, W.; Shen, R.-F.; Lan, P. Genes of acyl carrier protein family show different expression profiles and overexpression of acyl carrier protein 5 modulates fatty acid composition and enhances salt stress tolerance in Arabidopsis. Front. Plant Sci. 2017, 8, 987. [Google Scholar] [CrossRef]
- Yang, J.I.; Oksoo, H.; Gap, C.C.; Baik, H.C. Antisense expression of an Arabidopsis omega-3 fatty acid desaturase gene reduces salt/drought tolerance in transgenic tobacco plants. Mol. Cells 2002, 13, 264–271. [Google Scholar]
- Jin, X.-L.; An, J.-B.; Qi, D.-S.; Qiao, F.; Jiang, D.; Du, S.-B.; Ji, S.; Xie, H.-C. Identification and expression pattern analysis of FAD gene family in Populus tomentosa. Mol. Plant Breed. 2023, 1–23. [Google Scholar] [CrossRef]
- Kim, H.; Go, Y.S.; Kim, A.Y.; Lee, S.; Kim, K.-N.; Lee, G.-J.; Kim, G.-J.; Suh, M.C. Isolation and functional analysis of three microsomal delta-12 fatty acid desaturase genes from Camelina sativa (L.). Plant Biochem. 2014, 41, 146–158. [Google Scholar]
- Ashwini, V.R.; Narendra, Y.K.; Sanjay, P.B.; Abhay, M.H.; Prakash, B.G.; Vidya, S.G. Differential transcriptional activity of SAD, FAD2 and FAD3 desaturase genes in developing seeds of linseed contributes to varietal variation in α-linolenic acid content. Phytochemistry 2014, 98, 41–53. [Google Scholar]
- Nishiuchi, T.; Hamada, T.; Kodama, H.; Iba, K. Wounding changes the spatial expression pattern of the arabidopsis plastid omega-3 fatty acid desaturase gene (FAD7) through different signal transduction pathways. Plant Cell 1997, 9, 1701–1712. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.-W.; Tian, Y.; Wei, L.-H. Research progress of relationship between Staphylococcus aureus surface protein and the biofilm. China Mod. Med. 2022, 29, 29–32. [Google Scholar]
- Vinh, N.P.; Inyup, P.; Ute, H.; Enamul, H. Genomic evidence reveals SPA-regulated developmental and metabolic pathways in dark-grown Arabidopsis seedlings. Physiol. Plant. 2020, 169, 380–396. [Google Scholar]
- Chen, W.-L.; Zhang, Q.-Q.; Tang, S.-H.; Gong, W.; Hong, Y.-Y. Glycerol-3-phosphate acyltransferase in lipid metabolism, growth and response to stresses in plants. Plant Physiol. J. 2018, 54, 725–735. [Google Scholar]
- Yang, C.-L.; Duan, R.-J.; Wu, X.-X.; Qi, C.-Y.; Ma, Y.-H.; Xiong, H.-Y. Genome-wide identification, sequence variation, and expression of the GPAT gene family in Medicago truncatula. Pratacultural Sci. 2021, 38, 1966–1974. [Google Scholar]
- Wang, B.-K.; Wang, J.; Yang, T.; Wang, J.-X.; Dai, Q.; Zhang, F.-L.; Xi, R.; Yu, Q.-H.; Li, N. The transcriptional regulatory network of hormones and genes under salt stress in tomato plants (Solanum lycopersicum L.). Front. Plant Sci. 2023, 14, 1115593. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Chen, H.; Zhang, Y.; Thomas, H.-R.; Frank, M.-H.; He, Y.-H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Name | Amino Acid Number | Molecular Weight | Isoelectric Point | Fat Index | Hydrophilic/Hydrophobic Proteins | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| Solyc01T000115.1 | SlFAD1 | 383 | 43,801.58 | 8.70 | 90.05 | Hydrophilic | cyto |
| Solyc03T001114.1 | SlFAD2 | 378 | 43,710.22 | 8.04 | 87.59 | Hydrophilic | plas |
| Solyc04T000841.1 | SlFAD3 | 771 | 89,844.76 | 8.56 | 92.14 | Hydrophilic | plas |
| Solyc05T002114.1 | SlFAD4 | 439 | 50,630.55 | 8.62 | 88.18 | Hydrophobic | plas |
| Solyc06T000108.1 | SlFAD5 | 377 | 43,959.96 | 8.90 | 90.69 | Hydrophilic | plas |
| Solyc06T000109.1 | SlFAD6 | 377 | 44,085.12 | 8.45 | 90.69 | Hydrophilic | E.R. |
| Solyc06T000998.1 | SlFAD7 | 435 | 49,659.73 | 7.78 | 83.59 | Hydrophilic | chlo |
| Solyc07T000048.1 | SlFAD8 | 441 | 50,592.87 | 8.84 | 89.37 | Hydrophilic | E.R. |
| Solyc08T001312.1 | SlFAD9 | 447 | 51,626.60 | 8.64 | 84.79 | Hydrophobic | plas |
| Solyc10T000506.1 | SlFAD10 | 439 | 50,655.63 | 8.70 | 89.70 | Hydrophobic | plas |
| Solyc10T000508.1 | SlFAD11 | 439 | 50,577.54 | 8.68 | 89.93 | Hydrophobic | plas |
| Solyc12T002062.1 | SlFAD12 | 736 | 86,023.56 | 8.69 | 88.22 | Hydrophilic | plas |
| Solyc12T002837.1 | SlFAD13 | 379 | 43,983.43 | 8.09 | 95.67 | Hydrophilic | chlo |
| Solyc12T002836.1 | SlFAD14 | 379 | 43,855.47 | 8.79 | 89.26 | Hydrophilic | E.R. |
| Solyc12T002838.1 | SlFAD15 | 406 | 47,182.43 | 8.90 | 90.52 | Hydrophilic | plas |
| Solyc12T002060.1 | SlFAD16 | 399 | 46,814.10 | 8.67 | 94.31 | Hydrophilic | plas |
| Solyc12T002061.1 | SlFAD17 | 399 | 46,814.10 | 8.67 | 94.31 | Hydrophilic | plas |
| Solyc12T002056.1 | SlFAD18 | 268 | 31,033.89 | 9.02 | 96.04 | Hydrophilic | cyto |
| Solyc12T002835.1 | SlFAD19 | 627 | 70,440.21 | 8.99 | 96.73 | Hydrophobic | cyto |
| Solyc01T000424.1 | SlFAB1 | 330 | 37,729.03 | 5.72 | 69.15 | Hydrophilic | chlo |
| Solyc03T001283.1 | SlFAB2 | 393 | 44,830.14 | 6.14 | 77.43 | Hydrophilic | chlo |
| Solyc06T001235.1 | SlFAB3 | 760 | 87,195.34 | 6.13 | 85.33 | Hydrophilic | chlo |
| Solyc06T001110.1 | SlFAB4 | 397 | 45,237.55 | 6.04 | 79.65 | Hydrophilic | chlo |
| Solyc06T001237.1 | SlFAB5 | 387 | 44,425.98 | 7.03 | 85.17 | Hydrophilic | chlo |
| Solyc06T000610.1 | SlFAB6 | 166 | 18,924.74 | 8.31 | 74.04 | Hydrophilic | cyto |
| Solyc11T000349.1 | SlFAB7 | 393 | 44,534.04 | 6.24 | 83.38 | Hydrophilic | chlo |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, R.; Liu, H.; Chen, Y.; Zhuang, H.; Han, H.; Wang, H.; Wang, Q.; Li, N. Genome-Wide Characterization of Tomato FAD Gene Family and Expression Analysis under Abiotic Stresses. Plants 2023, 12, 3818. https://doi.org/10.3390/plants12223818
Xi R, Liu H, Chen Y, Zhuang H, Han H, Wang H, Wang Q, Li N. Genome-Wide Characterization of Tomato FAD Gene Family and Expression Analysis under Abiotic Stresses. Plants. 2023; 12(22):3818. https://doi.org/10.3390/plants12223818
Chicago/Turabian StyleXi, Rui, Huifang Liu, Yijia Chen, Hongmei Zhuang, Hongwei Han, Hao Wang, Qiang Wang, and Ning Li. 2023. "Genome-Wide Characterization of Tomato FAD Gene Family and Expression Analysis under Abiotic Stresses" Plants 12, no. 22: 3818. https://doi.org/10.3390/plants12223818
APA StyleXi, R., Liu, H., Chen, Y., Zhuang, H., Han, H., Wang, H., Wang, Q., & Li, N. (2023). Genome-Wide Characterization of Tomato FAD Gene Family and Expression Analysis under Abiotic Stresses. Plants, 12(22), 3818. https://doi.org/10.3390/plants12223818