1. Introduction
Chemical pesticides are among the most applied agrochemicals for pest control. However, their repeated application has resulted in an increased risk of emergence of pest resistance and soil degradation, making the overuse of these synthetic pesticides a new problem for global environment preservation [
1,
2]. The potential hazardous effect on the environment and on human and animal health associated with the abuse of chemicals has encouraged scientists to search for eco-friendly alternatives for pest control [
3,
4]. The search for alternatives to these chemical pesticides with natural-based products and biological molecules would provide a sustainable solution to protect both crops and the environment [
5]. In this context, allelochemicals are molecules synthesized by plants and known to have several biological activities. These molecules are particularly involved in the defense systems of plants against pests and are also synthesized as part of the allelopathy process [
6,
7,
8]. It has been reported that several allelochemical molecules have different biological activities. They can act as phytotoxic molecules and thus inhibit the germination and growth of several plant species [
8,
9]. They can exert different types of effects against insects, both attractive and repellent, and even insecticidal and larvicidal effects [
3]. They are also known to inhibit the growth of various microorganisms, such as bacteria, fungi and yeasts [
10,
11,
12].
In this way, allelochemicals could be of great interest for the development of new eco-friendly bio-pesticides [
11,
13]. In fact, these molecules have a short half-life and therefore problems regarding the persistence of residues in the environment can be solved and they can be considered safe and eco-friendly for the environment [
13].
EOs plays a crucial role in the allelopathic interactions between plants and species of their biotopes, such as weeds, fungi, bacteria and insects [
3,
5,
10]. Terpenes are known as potent allelochemicals with herbicidal and antifungal activity [
6]. These molecules can be a good source for the discovery of natural fungicides and herbicides and can limit the harmful environmental effects caused by chemical pesticides [
11]. According to the literature, the EOs of various plants have been reported to possess a significant phytotoxic effect against weeds [
5,
8,
11]. Hence, exploiting the allelopathic potential of molecules with herbicidal properties could be a promising approach for weed control [
4,
7,
11,
14].
The
Eucalyptus genus is part of the
Myrtaceae family, comprising forest trees native to Australia; the
Eucalyptus genus includes 13 subgenera and more than 700 species [
15]. More than 100 species of
Eucalyptus have been introduced in Tunisia since the 1950s. These species have been used for reforestation programs. They are mainly exploited for wood production, erosion control and the production of EOs.
Eucalyptus species EOs are a mixture of volatile terpenes, such as 1,8-cineole, globulol, citronellol, spathunelol, limonene and pinenes [
5,
8,
9].
Various molecules extracted from
Eucalyptus species have been reported to have toxic effects against weeds, insects and microorganisms [
5,
8,
9,
15]. Plants’ secondary metabolites, known to possess several activities, and their applications in the field make them good alternatives for chemical pesticides without side effects—in particular, the persistence linked to the non-biodegradability of pesticides and resulting in toxicity to the environment, cultivated plants, wild animals, beneficial microorganisms and several insects [
13,
16].
The essential oils of
E. falcata,
E. sideroxylon and
E. citriodora, which grow in Tunisia, have been the subject of recent studies [
15,
17,
18,
19]; however, studies of their herbicidal potential have not been performed. Likewise, the majority of studies on their antifungal potential are focused on clinical fungi, and few are studies of phyto-pathogenic fungi [
17]. The herbicidal activity of
Eucalyptus citriodora has been reported in India [
20], but their mode of action was not described. On the other hand, knowing that chemical ecology is a very important strategy, it is widely exploited for the discovery of metabolites because plants are able to synthesize various molecules with the aim of adaptation and acclimatization to environmental pressure. The synthesis of these molecules continues to evolve according to the pedoclimatic conditions of their biotopes in order to act in a physiological or ecological way on the maintenance of the plant in its ecological environment [
13]. This may explain the variability in the production of essential oils depending on the origin of the plant and the usefulness of testing plant material from different origins.
In this sense, the present work aims to determine the chemical compositions of the EOs of E. falcata, E. sideroxylon and E. citriodora growing in Tunisia, to evaluate their antioxidant potential and to test their antifungal effects against eight strains of phyto-pathogenic fungi, as well as to study their herbicidal activity against the germination and seedling growth of weeds and cultivated crops at the germination stage. Moreover, at the post-emergence stage, our objective is to test the effect of EOs applied by spraying on the physiological effects of plants: the relative water content, the relative leakage of electrolytes, the synthesis of chlorophyll, the rate of malondialdehyde resulting from lipid peroxidation and also the proline content, always referring to the activity of a reference synthetic herbicide: glyphosate.
4. Discussion
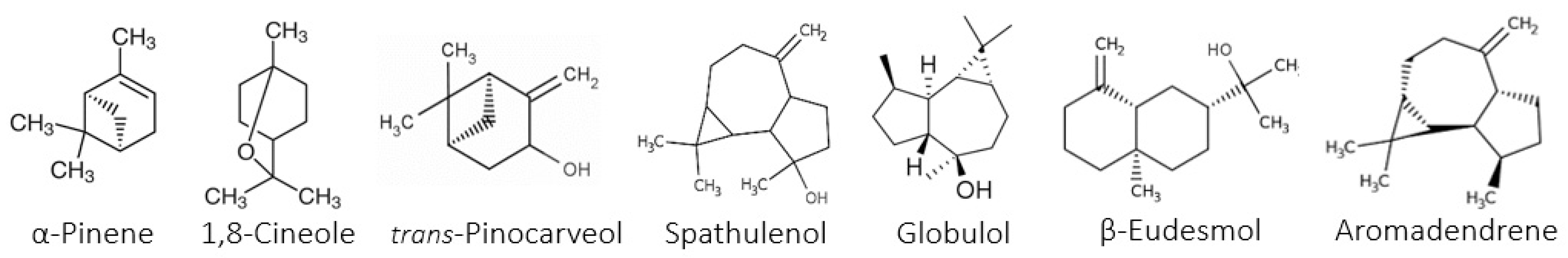
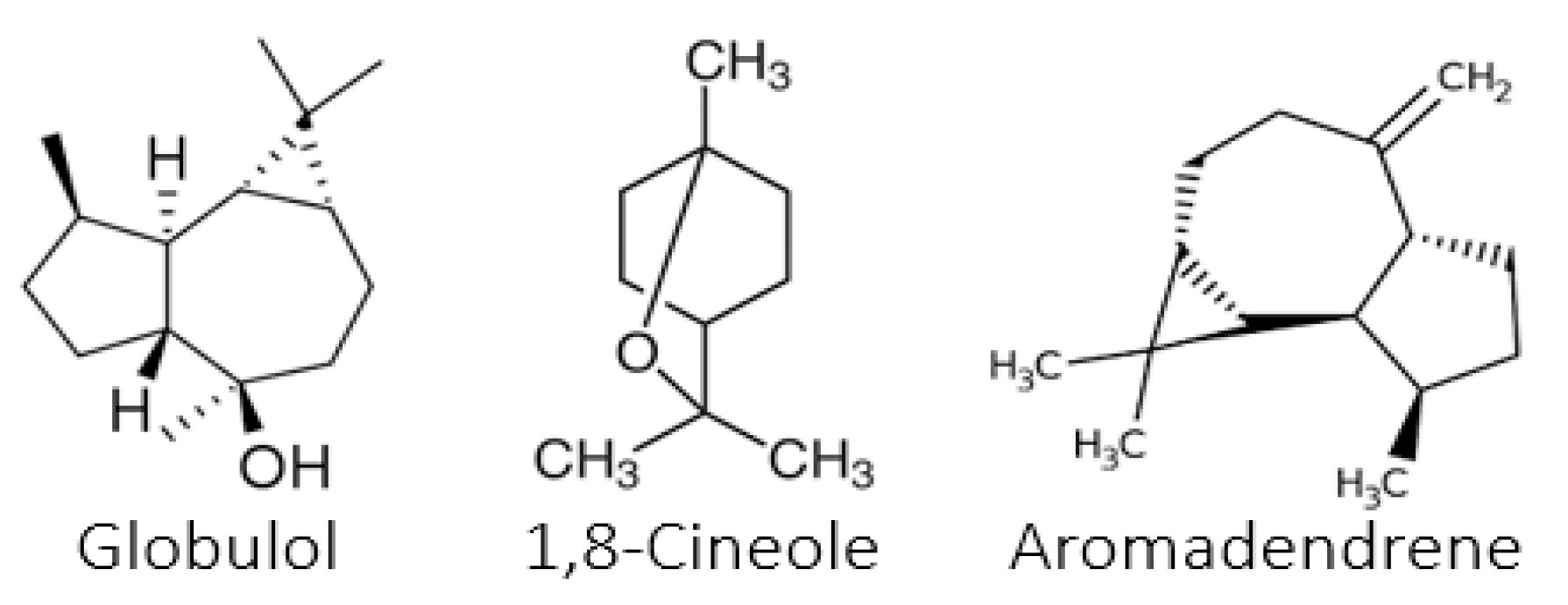
The study of the chemical composition of the essential oils of the three species of
Eucalyptus revealed the great importance and richness of various compounds. According to the literature, among the few studies on
E. falcata EOs, three studies were performed on plant material from Tunisia [
17,
18,
19]. The chemical composition of the EOs of
E. falcata is in agreement with that reported in the present study. In fact, the authors stated that the major components were 1,8-cineole (30.7%), trans-pinocarveol (26%), globulol (7%), and α-pinene (6%) [
17]. However, the main differences with the previous studies consist in the absence of spathunelol, the presence of globulol, and the high levels of trans-pinocarveol [
17,
18].
Similar results were obtained in previous reports for
E. sideroxylon [
15,
19]. According to these studies, the EOs of
E. sideroxylon were characterized by their high content of 1,8-cineole (69–81%), which is in agreement with the present study.
Several investigations on
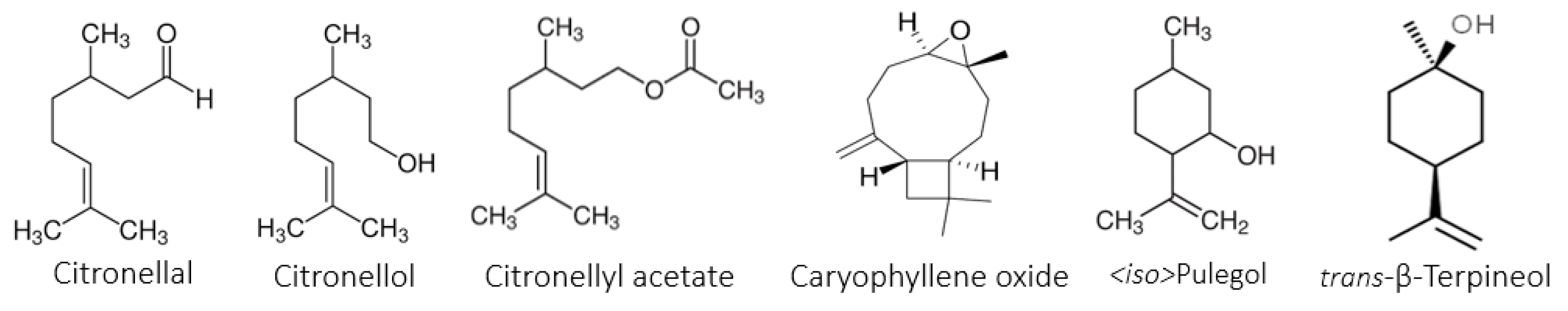
E. citriodora EOs have been reported worldwide. In fact, in Colombian
E. citriodora oils [
30,
31], in India [
20], in Taiwan [
32], and in the Congo [
33], the major components were citronellal (40–72%), citronellol (6–22%), and iso-pulegol (3–13%), which is in agreement with our obtained data. However, citronellyl acetate was detected for the first time in this study. On the other hand, our study is in contrast with the results of a Tunisian report [
2,
34] wherein the authors evidenced a different chemotype characterized by a specific richness in 1,8-cineole (54%) and α-pinene (23%) and the total absence of citronellal, citronellol, and iso-pulegone. Apart from this, the data of the current study are in good agreement with the literature. The small differences in the percentages may be due to several factors, such as the genetic background, biotic and abiotic factors, season of collection, and extraction methods [
35].
The evaluation of the antioxidant potential of the three oils showed interesting activities that can contribute to the valorization of essential oils of forest species, particularly of the Eucalyptus genus. These results are in agreement with the literature.
Indeed, several studies have investigated the antioxidant properties of different
Eucalyptus species, such as
E. oleosa,
E. grandis ×
E. urophylla,
E. gracilis,
E. citriodora,
E. salubris, and
E. salmonophloia [
36,
37,
38,
39].
Furthermore, the EOs of
E. citriodora exhibited higher antioxidant potential than those of
E. falcata and
E. sideroxylon. This appears to be related to the high level of oxygenated terpenes in
E. citriodora oil [
36,
37].
Similarly, the EOs of
E. citriodora, collected in India, contain oxygenated monoterpenes such as citronellal (60.66%), exhibiting strong antioxidant activity, in agreement with the current study [
38]. In the same report, the authors evaluated and showed the good antioxidant activity of α-citronellal, β-citronellol, and isopulegol, the major compounds of the oil. Furthermore, the synergism between oil components could be related to the remarkable antioxidant capacity of the tested oils [
40].
The study of the allelopathic properties of the EOs obtained from the three species of Eucalyptus revealed remarkable herbicidal potential, which even exceeds the activity of the synthetic herbicide glyphosate.
The germination- and growth-inhibiting effects of the
Eucalyptus oils tested in the present study appear to be linked to the presence of several chemical components, particularly monoterpenes and sesquiterpenes, which are the main constituents and are known for their herbicidal potential [
41]. Indeed, EOs of species belonging to the
Myrtaceae family are known for their richness in allelochemicals [
5,
8,
9,
16,
42]. In particular, in previous studies, we showed the phytotoxic potential of volatile oils and crude extracts of
Eucalyptus erythrocorys against weeds [
16,
42].
Recently, the herbicidal potential of 22
Eucalyptus species growing in Tunisia has been reported. These studies reveal enormous allelopathic potential linked to the EOs of
Eucalyptus species; this could be in agreement with the present study [
5,
8,
9,
16].
Observing the chemical composition of the
Eucalyptus oils tested in the present work (
Table 2), we can assume that the activity is due to different components, such as 1,8-cineole (0.3–65.4% in
Eucalyptus oils), citronellal (48.7% in
E. citriodora oil), citronellol (20.2% in
E. citriodora oil), and α-pinene (ranging between 0 and 6% in
Eucalyptus oils), already known for their significant phytotoxic effects [
20,
43,
44].
Several studies report that terpenes, both hydrocarbons and oxygenated, are phytotoxic molecules. Additionally, oxygenated monoterpenes (51.3, 75.5, and 90%, respectively, in
E. falcata,
E. sideroxylon, and
E. citriodora) have been reported to have stronger herbicidal properties than the corresponding hydrocarbon derivatives [
20,
43,
44]; this can explain the high herbicidal activity of
E. citriodora compared to
E. falcata and
E. sideroxylon.
To explain the mode of action of terpenes on the inhibition of germination, various physiological and biochemical mechanisms have been reported.
In fact, such monoterpenes can affect physiological functions, such as cell division, viability, and growth inhibition by the alteration of membrane integrity [
45,
46].
Other studies have reported that EOs reduce germination and growth by inhibiting the incorporation of nitrogen into amino acid synthesis, leading to the accumulation of ammonia, and by impairing photosynthesis and photorespiration [
47].
In the same way, Abrahim et al., 2003 demonstrated that the monoterpene hydrocarbon α-pinene reduces the growth of maize by inhibiting electron transfer and uncoupling oxidative phosphorylation, resulting in the alteration of energy metabolism and the blocking of ATP synthesis in the mitochondria [
48]. Similarly, some sesquiterpenes have been reported to have phytotoxic effects by causing oxidative stress, the inhibition of photosynthesis, and the induction of microtubular alterations [
47]. Previous reports have shown that EOs induce oxidative stress to weeds and increase peroxidase and superoxide dismutase activities [
49,
50,
51]. Other studies have shown that EOs and their constituents have remarkable herbicidal potential, the effects of which result in anatomical and physiological effects, such as the formation of lipid globules in the cytoplasm, inhibition of mitochondrial development, and alteration of the integrity of the membrane of the nucleus and mitochondria [
52,
53].
For post-emergence trials and the application of EOs by spraying, few studies have been undertaken and the exact modes of action still remain poorly known. On the other hand, data obtained in this study are in agreement with those reported in the literature. Several studies have reported that various constituents of EOs, when applied at the post-emergence stage, are able to inhibit the growth of weeds [
54], hypothesizing oxidative stress and alteration of the water content [
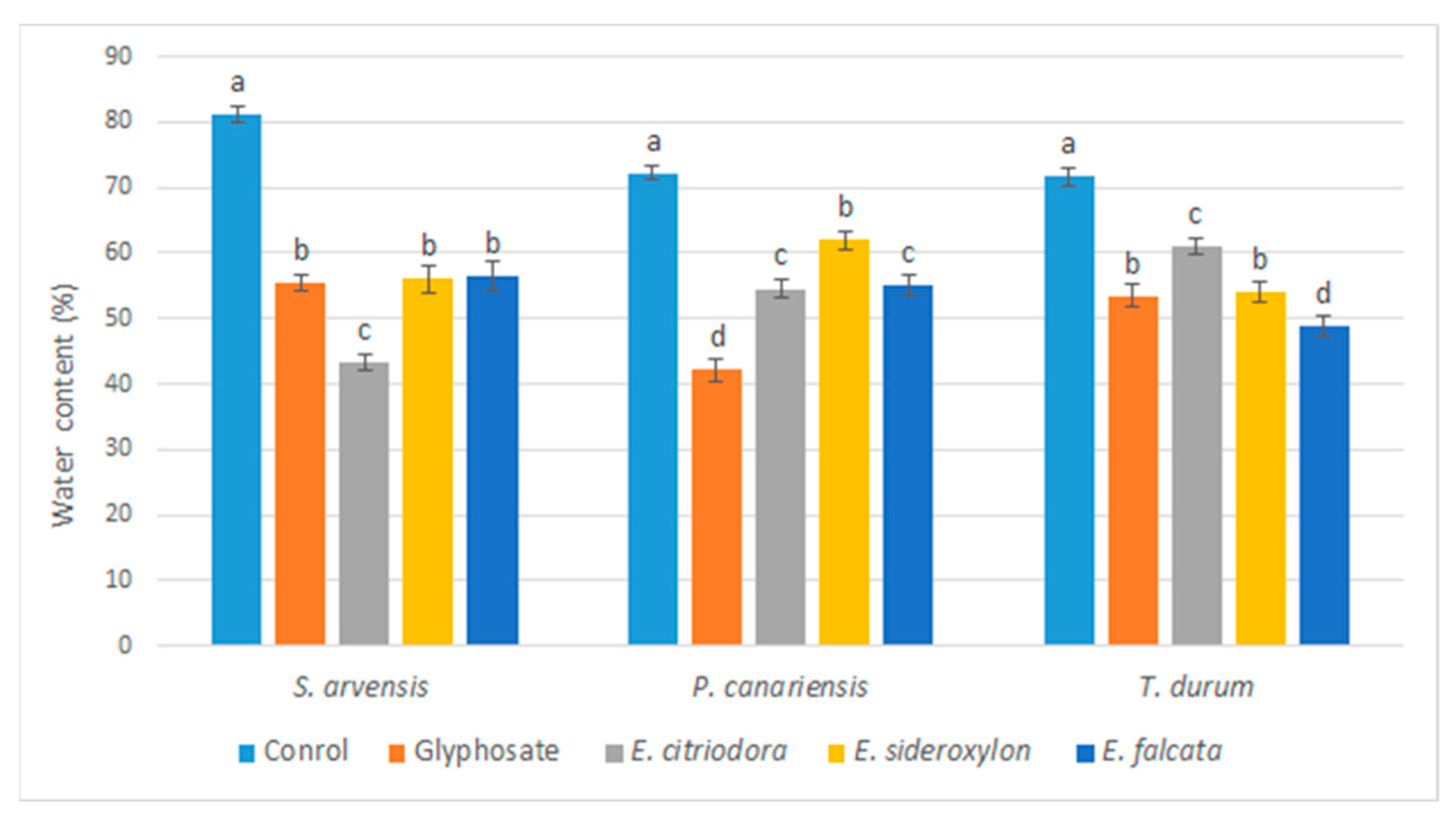
47]. The evidence that emerged in the present study showed a correlation between the decrease in the water content and the arrest of the growth of the plants treated with
Eucalyptus EOs.
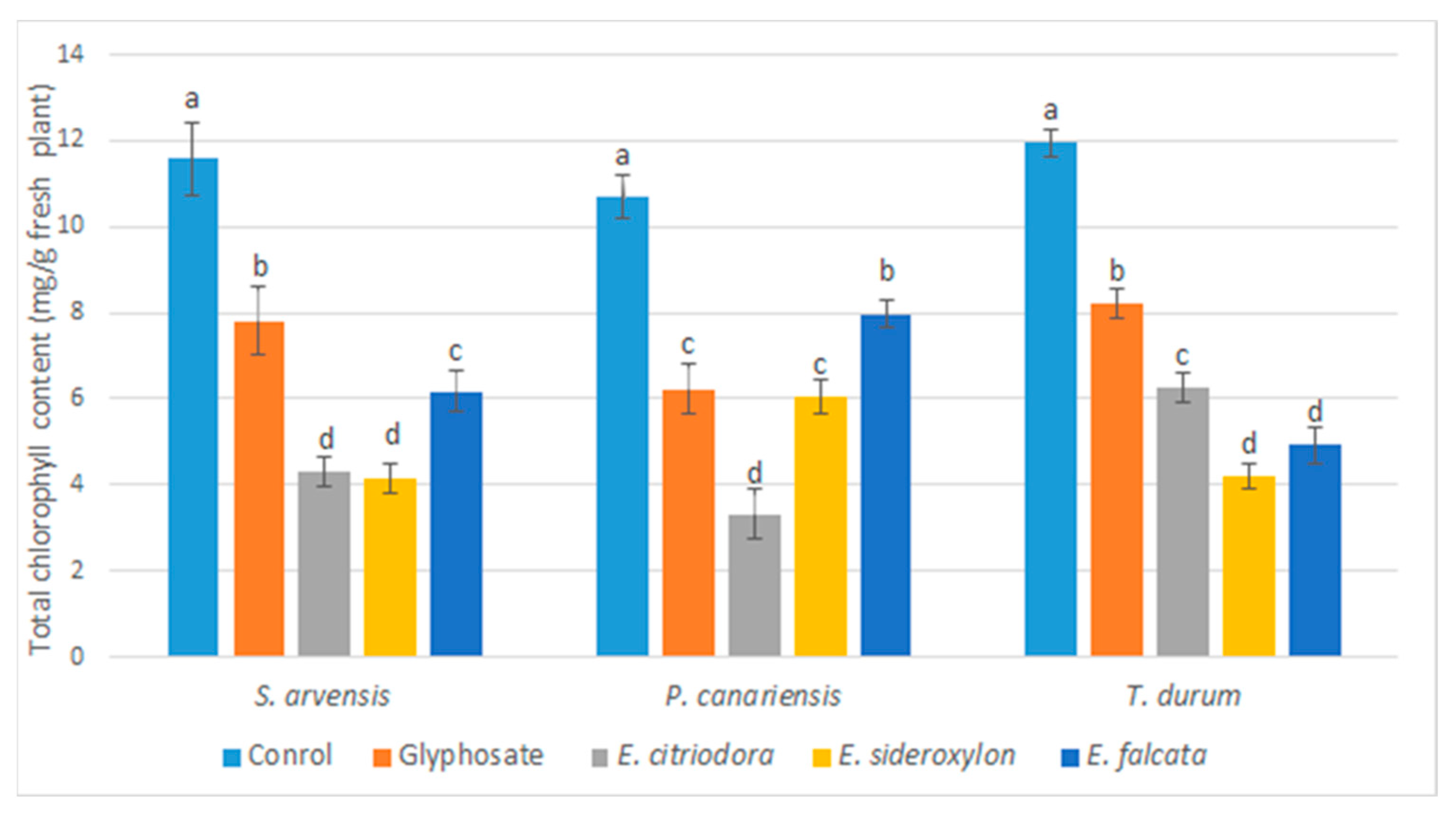
Following the application of
Eucalyptus EOs, a marked decrease in chlorophyll synthesis was noted, suggesting a correlation between the visible effects on weeds (i.e., chlorosis, necrosis, and photosynthetic functions) and their growth. According to previous studies, several EOs and their pure components have been described to inhibit photosynthesis and chlorophyll synthesis [
55]. Indeed, some oxygenated monoterpenes have been reported to induce a photosynthetic decrease by reducing the chlorophyll content in various weeds [
56].
In addition, citronellol and 1,8-cineole, major compounds of the
Eucalyptus species in the current study, reduced the chlorophyll content of
Ageratum conyzoides by 60 and 66%, respectively [
57,
58]. Earlier studies concluded that the decrease in chlorophyll content appeared to be related to the inhibition of chlorophyll synthesis and degradation of photosynthetic pigments [
59,
60].
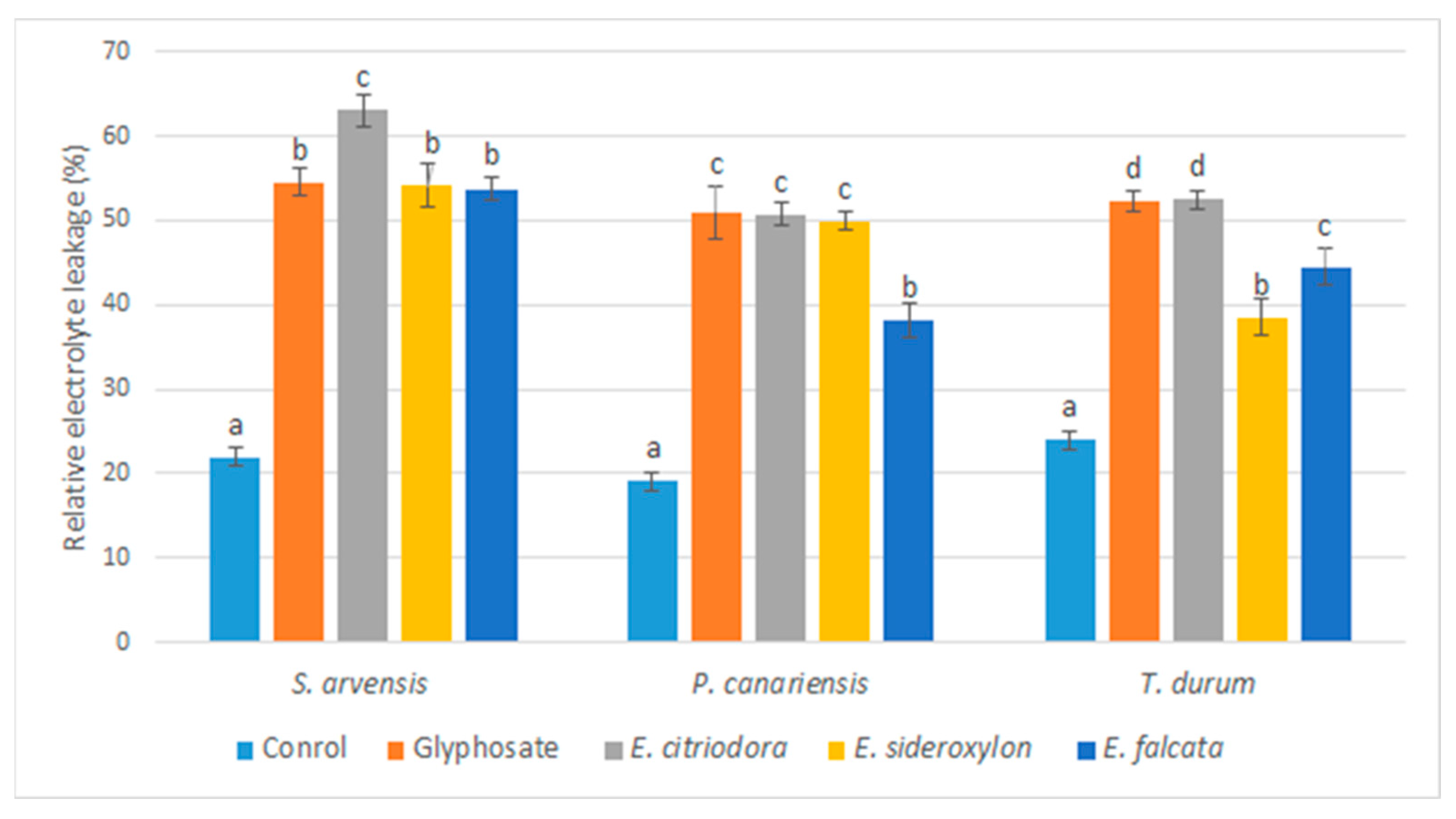
Among the physiological effects of
Eucalyptus EOs on membrane integrity, the excessive leakage of electrolytes was noted, which can be explained by the loss of integrity of membrane cells. On the other hand, membrane integrity is essential for vital functions and plays several roles, including that of a barrier, selective permeability to nutrients, and, indeed, control of the electrolyte balance [
61]. Various secondary metabolites have been reported to alter the integrity of the cell membrane in plants, particularly EOs and their pure constituents. A previous study has shown that linalool caused increased membrane permeability, further demonstrating that EOs can cause important damage to membrane permeability [
62]. According to the data obtained, the application of
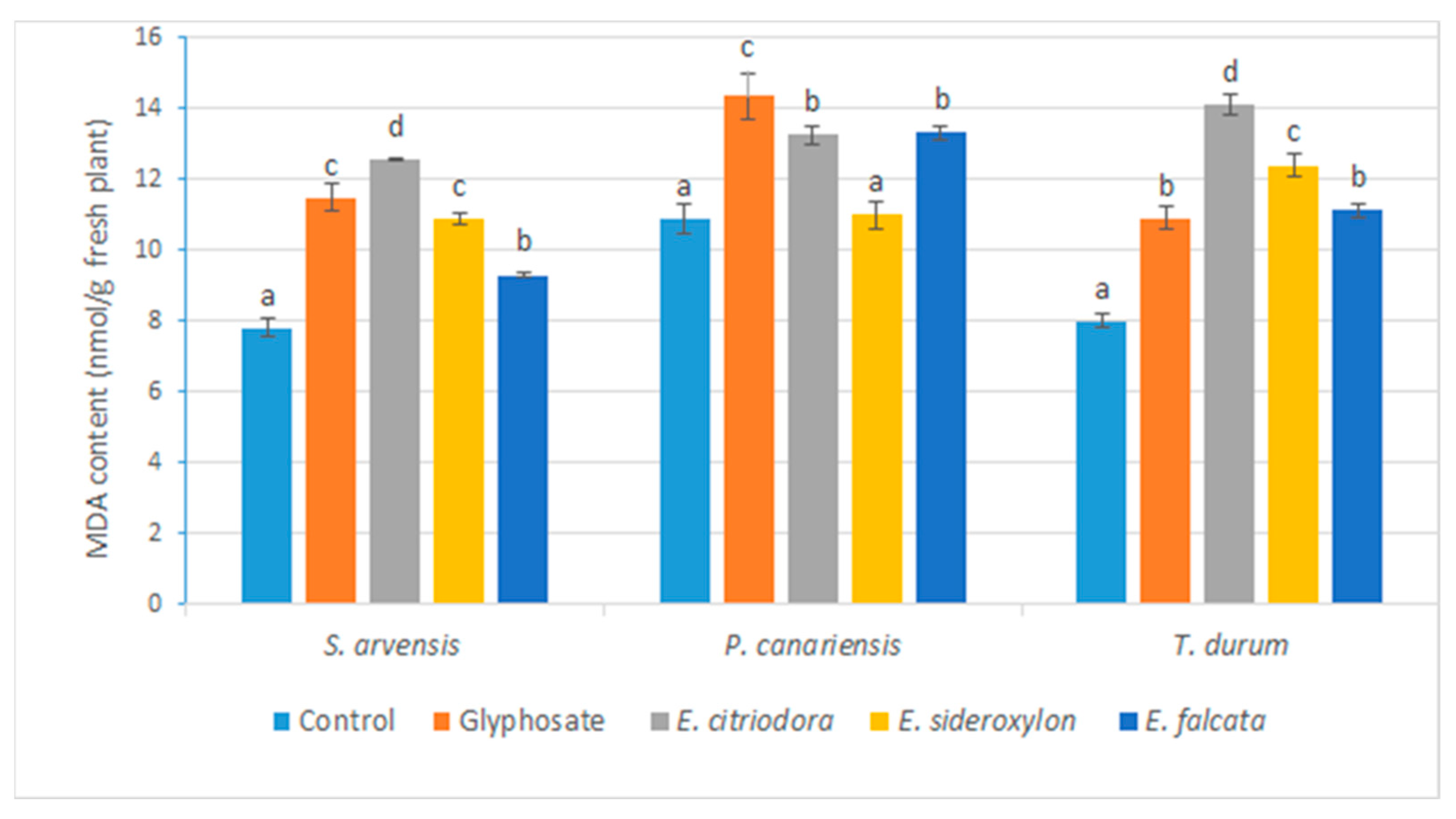
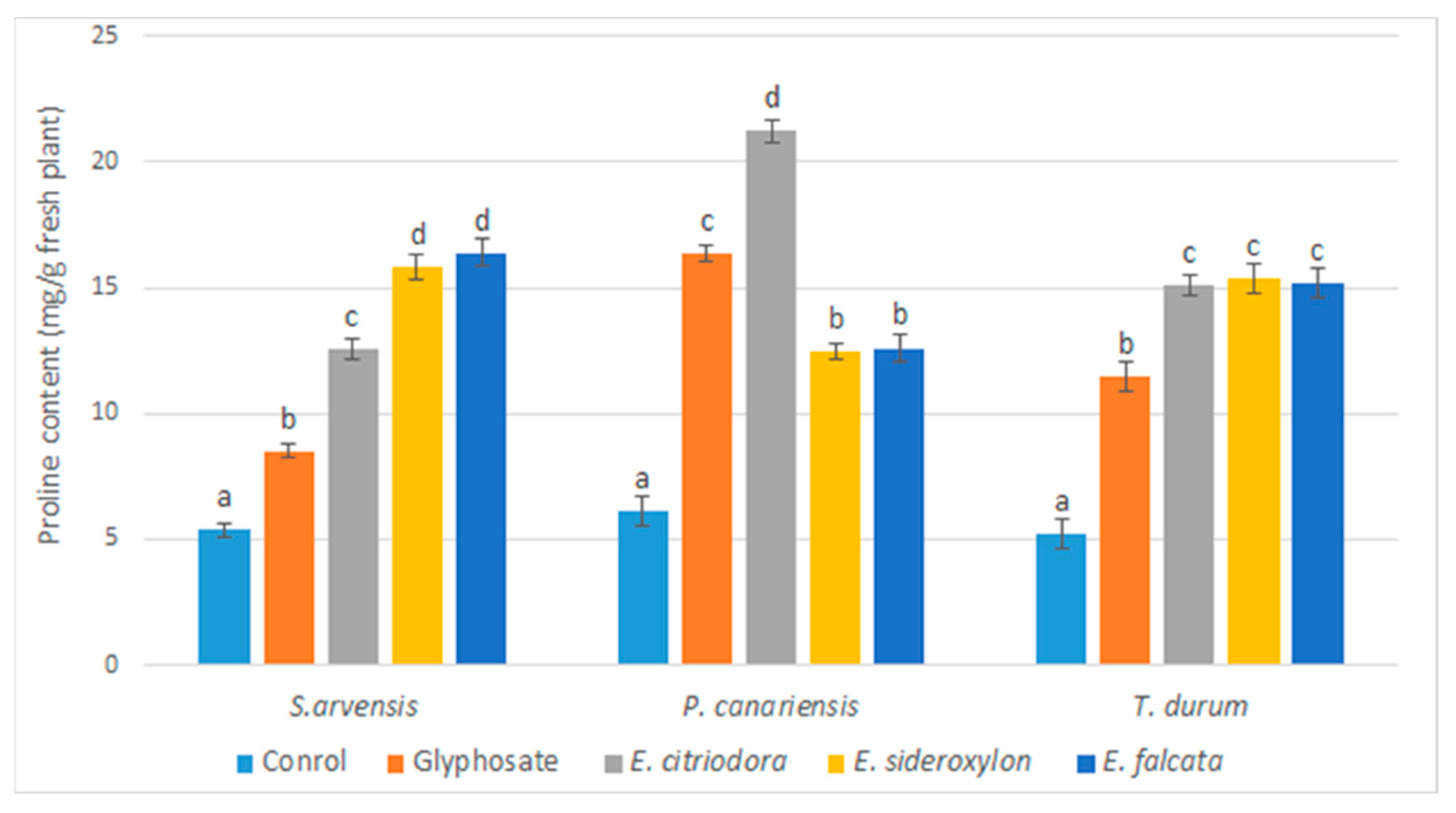
Eucalyptus EOs induced, in the tested herbs, an increase in the rate of MDA and proline compared to the control, in agreement with the previous observations. Indeed, it has been reported that the application of EOs during the post-emergence stage increases the permeability of the membrane by disrupting its integrity because of lipid peroxidation [
47,
63,
64,
65]. Additionally, some EOs and monoterpenes have been reported to induce oxidative stress. α-pinene caused lipid peroxidation when applied to
Cassia occidentalis, resulting in increased solute leakage [
25]. Furthermore, the increase in MDA levels was related to the lipid peroxidation of the membrane and alteration of its permeability.
Finally, the current study has shown that Eucalyptus oils have significant in vitro antifungal potential against the growth of phytopathogenic fungi.
The differences in the resistance of different fungal strains to
Eucalyptus volatile oil seem to be related to the different capacities of the oil components to disrupt the cell membrane [
66].
Eucalyptus oils are known to have remarkable antimicrobial activity that is related to their richness in oxygenated compounds [
67].
Indeed, we have previously reported the antifungal potential of
Eucalyptus erhytrocorys EOs and its crude extracts against phytopathogenic fungi [
16,
42], which is in agreement with the current study. Similarly, the inhibitory activity of the volatile oil obtained from
Eucalyptus camaldulensis against the growth of fungi such as
Thanatephorus cucumeris,
Aspergillus niger,
F. oxysporum, and
Rhizopus oryzae is also known [
67].
In agreement with the current study, Ivanov et al. (2021) demonstrated the fungicidal activity of 1,8-cineole and camphor (major compounds in
Eucalyptus volatile oils) against the growth of different
Candida species [
68]. In another research work, it was reported that terpinen-4-ol and α-terpineol, terpinolene, and 1,8-cineole, constituents of
Eucalyptus oils, are fungitoxic against
Botrytis cinerea [
69]. However, with regard to the antimicrobial mechanism, it has been reported that the apolar nature of terpenes may favor their penetration into the lipid bilayer of the fungal membrane, inducing its disruption. Many terpenes have been reported to exert their antifungal activity by increasing the permeability of fungal cells and membrane fluidity [
70].



 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}