
Untargeted Metabolomics for Integrative Taxonomy: Metabolomics, DNA Marker-Based Sequencing, and Phenotype Bioimaging

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
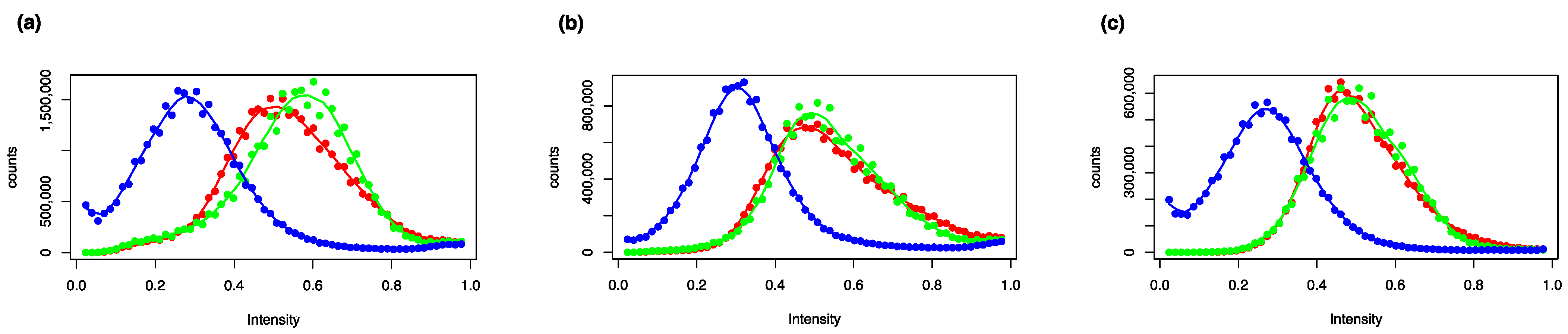
2.1. Phenotypic Analysis (Bioimaging)
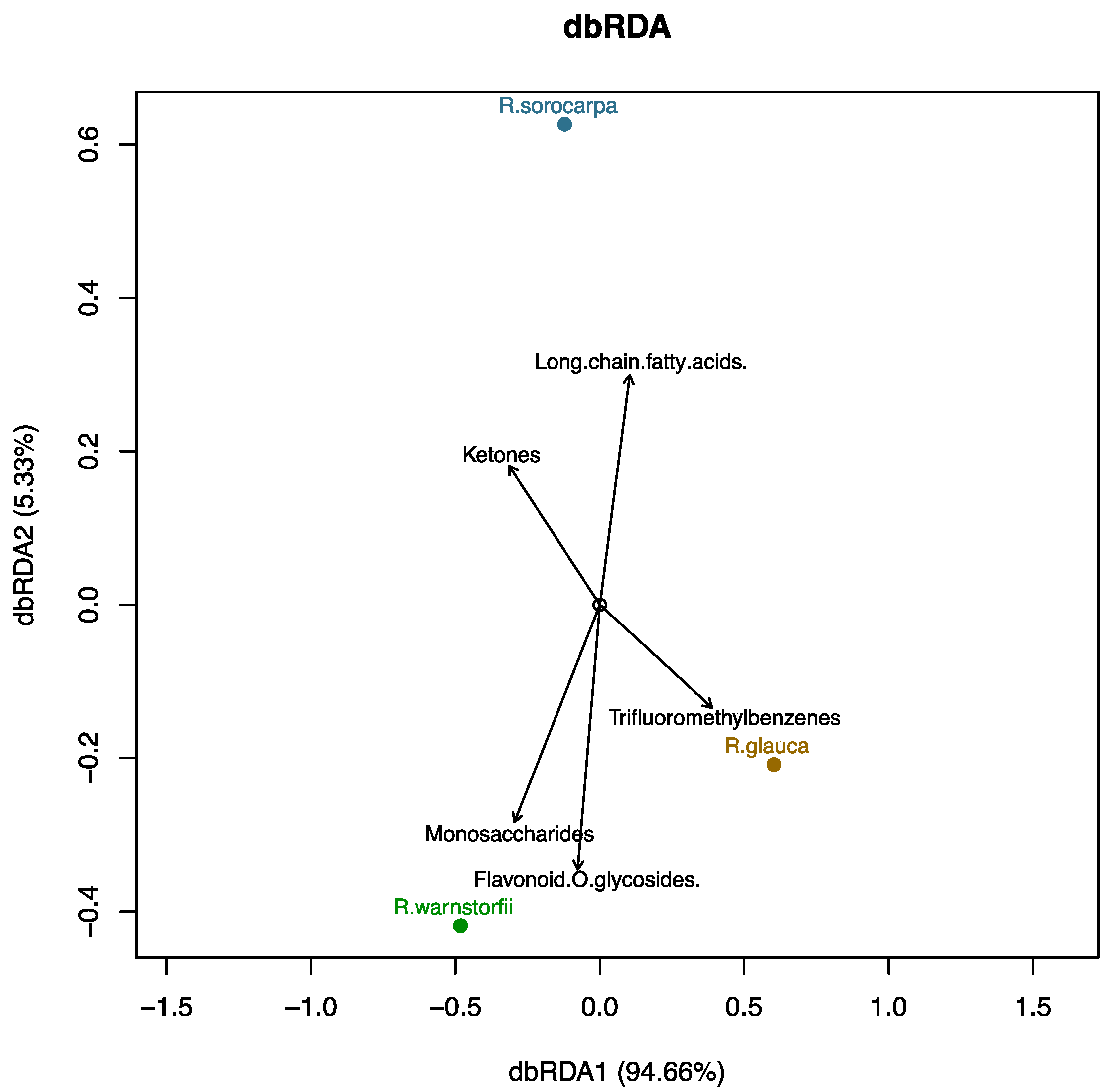
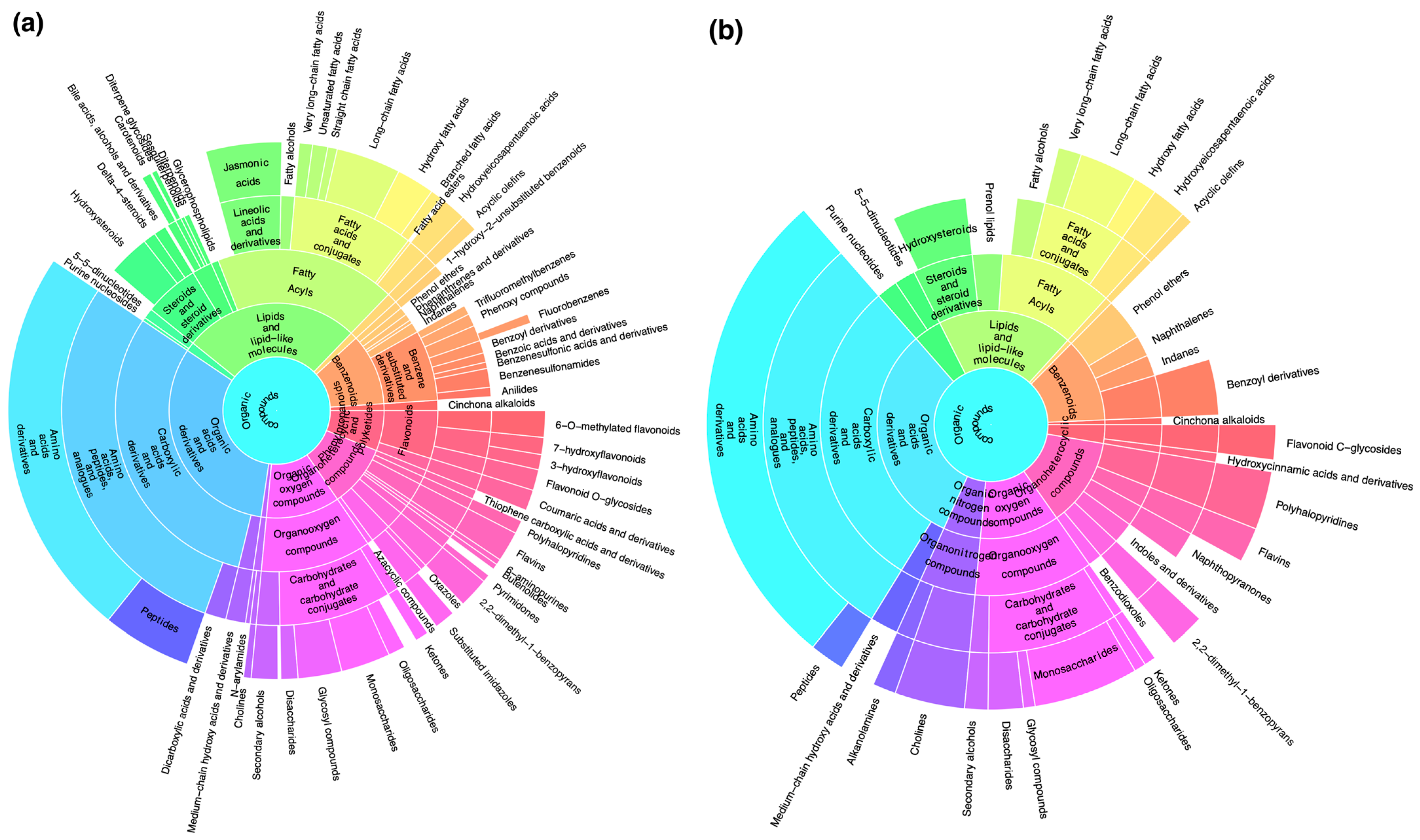
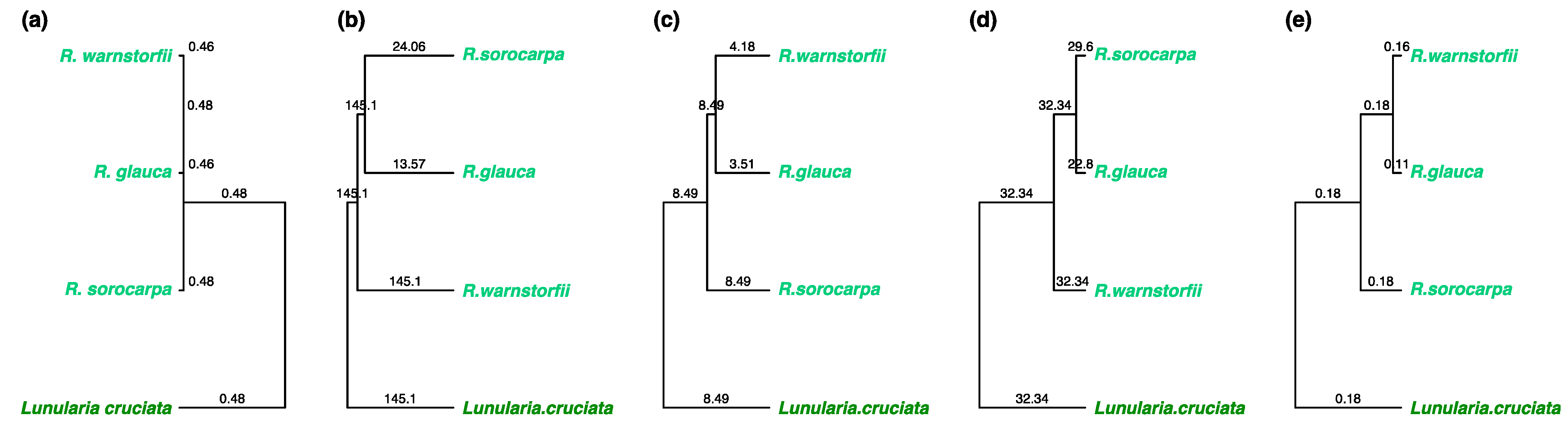
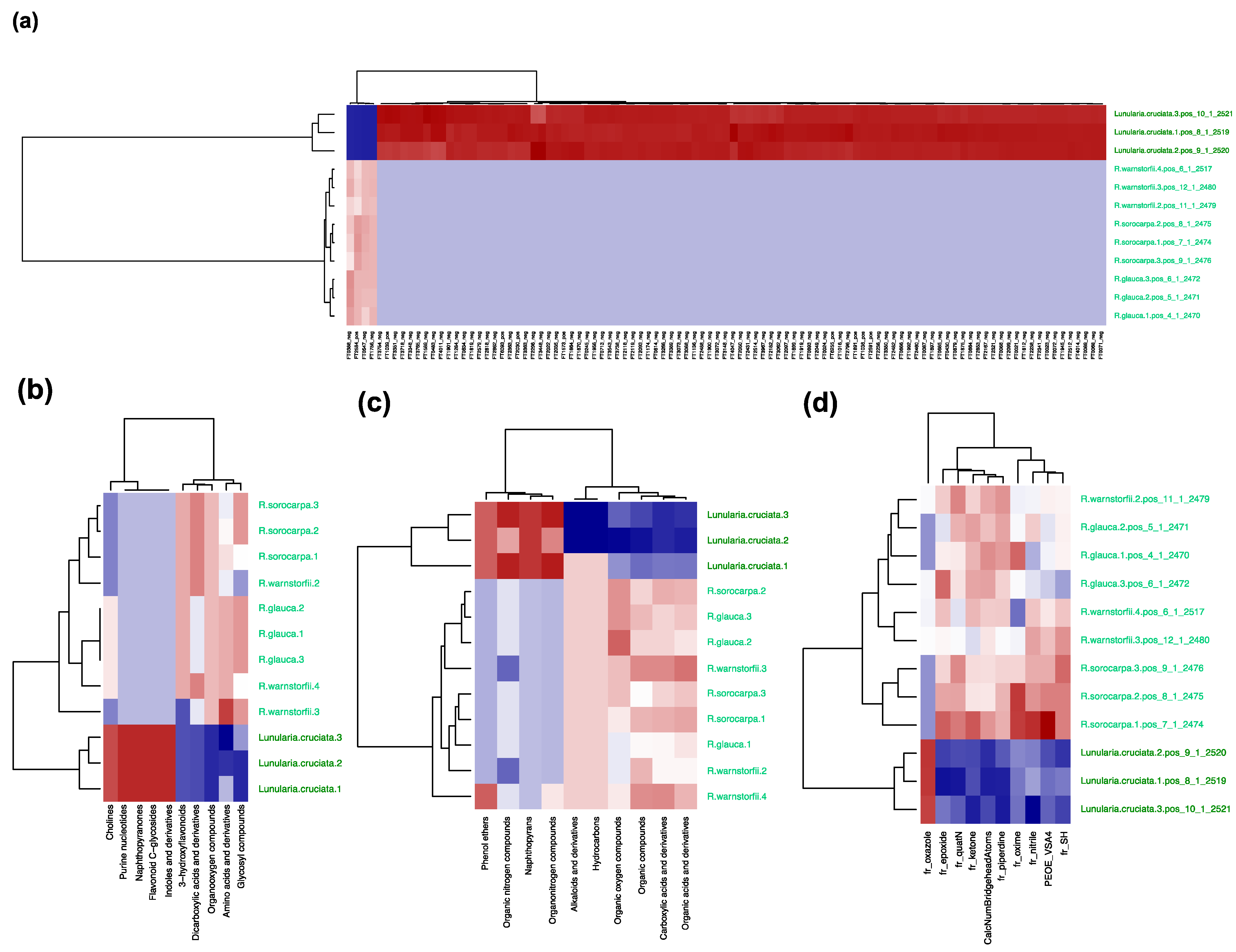
2.2. Chemotaxonomic Analysis Characterizing the Riccia Species Infragenerically
2.3. Chemotaxonomic Analysis Characterizing the Riccia Species at the Genus Level from the Outgroup
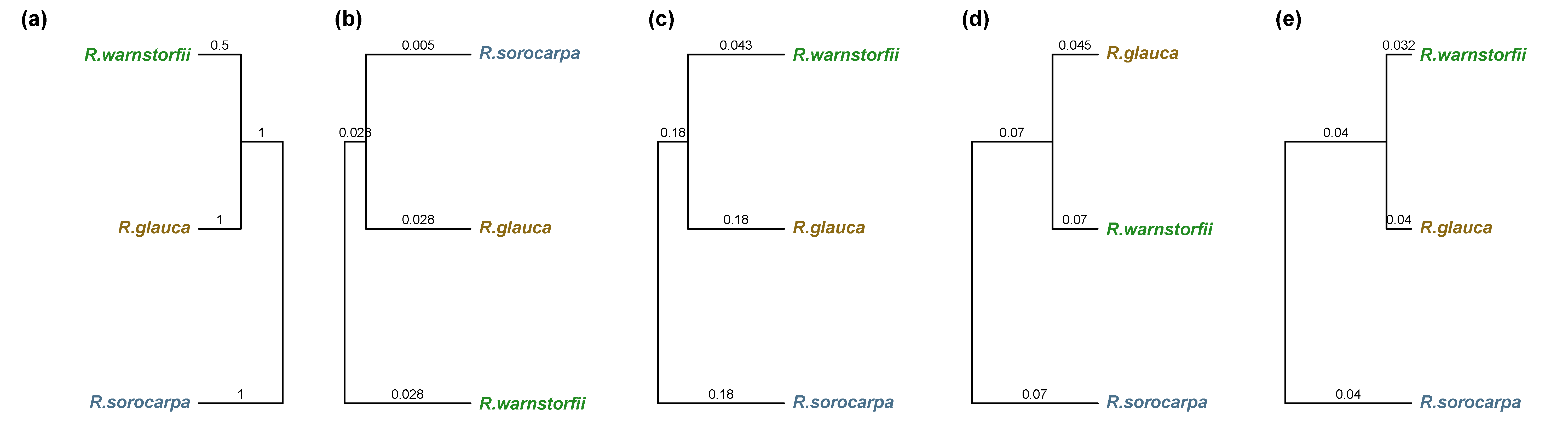
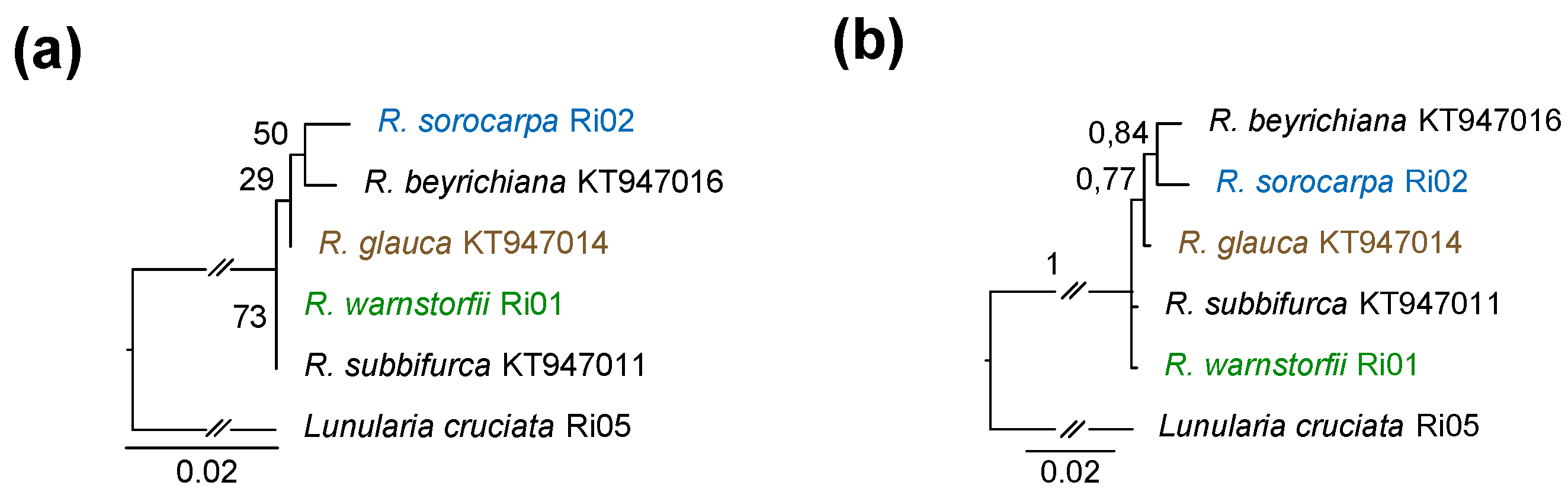
2.4. DNA Sequence Analysis
3. Discussion
3.1. DNA Sequence Data
3.2. Bioimaging Data
3.3. Chemotaxonomic Data
3.4. Novel Insights from Untargeted Chemotaxonomy
3.5. Applicability of Untargeted Chemotaxonomy
3.6. Integration of Untargeted Metabolomics into Integrative Taxonomy
4. Materials and Methods
4.1. Sample Collection and Processing
4.2. DNA Sequence Analysis
4.3. Phenotypic Analysis (Bioimaging)
4.4. Untargeted Metabolomics
4.4.1. Metabolite Extraction and Untargeted Mass-Spectrometry
4.4.2. Raw Data and MS1 Data Processing
4.4.3. Processing of MS/MS Data
4.4.4. Chemodiversity Analyses
4.4.5. Explorative and Unsupervised Multivariate Analyses
4.4.6. Selection of Chemophenetic Molecular Features
4.4.7. Construction of Taxonomic Trees
4.4.8. Deposition of Metabolomics Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative Taxonomy: A Multisource Approach to Exploring Biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef] [PubMed]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Jordon-Thaden, I.E.; Chanderbali, A.S.; Gitzendanner, M.A.; Soltis, D.E. Modified CTAB and TRIzol protocols improve RNA extraction from chemically complex Embryophyta. Appl. Plant Sci. 2015, 3, 1400105. [Google Scholar] [CrossRef] [PubMed]
- Križman, M.; Jakše, J.; Baričevič, D.; Javornik, B.; Prošek, M. Robust CTAB-activated charcoal protocol for plant DNA extraction. Acta Agric. Slov. 2006, 87, 427–433. [Google Scholar]
- Cargill, D.C.; Beckmann, K.; Seppelt, R. Taxonomic revision of Riccia (Ricciaceae, Marchantiophyta) in the monsoon tropics of the Northern Territory, Australia. Aust. Syst. Bot. 2021, 34, 336–430. [Google Scholar] [CrossRef]
- Wheeler, J.A. Molecular Phylogenetic Reconstructions of the Marchantioid Liverwort Radiation. Bryologist 2000, 103, 314–333. [Google Scholar] [CrossRef]
- Cargill, D.C.; Neal, W.C.; Sharma, I.; Gueidan, C. A preliminary molecular phylogeny of the genus Riccia L. (Ricciaceae) in Australia. Aust. Syst. Bot. 2016, 29, 197. [Google Scholar] [CrossRef]
- Dirkse, G.M.; Losada-Lima, A.; Stech, M. Riccia boumanii Dirkse, Losada & M.Stech sp. nov. (Ricciaceae, Marchantiophyta) in the Canary Islands, the first species of Riccia subgenus Riccia section Pilifer Volk outside South Africa. J. Bryol. 2016, 38, 94–102. [Google Scholar] [CrossRef]
- Hinchliff, C.E.; Smith, S.A.; Allman, J.F.; Burleigh, J.G.; Chaudhary, R.; Coghill, L.M.; Crandall, K.A.; Deng, J.; Drew, B.T.; Gazis, R.; et al. Synthesis of phylogeny and taxonomy into a comprehensive tree of life. Proc. Natl. Acad. Sci. USA 2015, 112, 12764–12769. [Google Scholar] [CrossRef] [Green Version]
- Fox, H.M. Chemical Taxonomy. Nature 1946, 157, 511. [Google Scholar] [CrossRef]
- McClure, J.W.; Miller, H.A. Moss chemotaxonomy. A survey for flavonoids and their taxonomicimplications. Nova Hedwig. 1967, 14, 111–125. [Google Scholar]
- Singh, R. Chemotaxonomy: A Tool for Plant Classification. J. Med. Plants Stud. 2016, 4, 90–93. [Google Scholar]
- Zidorn, C. Plant chemophenetics—A new term for plant chemosystematics/plant chemotaxonomy in the macro-molecular era. Phytochemistry 2019, 163, 147–148. [Google Scholar] [CrossRef] [PubMed]
- Brodo, I.M. Interpreting Chemical Variation in Lichens for Systematic Purposes. Bryologist 1986, 89, 132. [Google Scholar] [CrossRef]
- Rogers, R.W. Chemical variation and the species concept in lichenized ascomycetes. Bot. J. Linn. Soc. 1989, 101, 229–239. [Google Scholar] [CrossRef]
- Willer, J.; Christensen, E.; Wahl, A.; Gemeinholzer, B.; Zidorn, C. Phylogeny and chemophenetics of the newly described Doronicum × longeflorens and related Doronicum taxa (Senecioneae, Asteraceae). Biochem. Syst. Ecol. 2022, 101, 104400. [Google Scholar] [CrossRef]
- Culberson, W.L. The use of chemistry in the systematics of the lichens. Taxon 1969, 18, 152–166. [Google Scholar] [CrossRef]
- Lumbsch, H.T. Analysis of Phenolic Products in Lichens for Identification and Taxonomy. In Protocols in Lichenology; Kranner, I.C., Beckett, R.P., Varma, A.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 281–295. ISBN 978-3-540-41139-0. [Google Scholar]
- Figueiredo, A.C.; Sim-Sim, M.; Barroso, J.G.; Pedro, L.G.; Esquível, M.G.; Fontinha, S.; Luís, L.; Martins, S.; Lobo, C.; Stech, M. Liverwort Radula species from Portugal: Chemotaxonomical evaluation of volatiles composition. Flavour Fragr. J. 2009, 24, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Hawrył, A.; Bogucka-Kocka, A.; Świeboda, R.; Hawrył, M.; Stebel, A.; Waksmundzka-Hajnos, M. Thin-layer chromatography fingerprint and chemometric analysis of selected Bryophyta species with their cytotoxic activity. JPC J. Planar Chromatogr. Mod. TLC 2018, 31, 28–35. [Google Scholar] [CrossRef]
- Hu, T.; Jin, W.-Y.; Cheng, C.-G. Classification of Five Kinds of Moss Plants with the Use of Fourier Transform Infrared Spectroscopy and Chemometrics. Spectroscopy 2011, 25, 271–285. [Google Scholar] [CrossRef]
- Lee, G.E.; Bechteler, J.; Pócs, T.; Schäfer-Verwimp, A.; Tang, H.Y.; Chia, P.W. Integrative Taxonomy Reveals a New Species of the Genus Lejeunea (Marchantiophya: Lejeuneaceae) from Peninsular Malaysia. Plants 2022, 12, 1642. [Google Scholar] [CrossRef]
- Ludwiczuk, A.; Raharivelomanana, P.; Pham, A.; Bianchini, J.-P.; Asakawa, Y. Chemical variability of the Tahitian Marchantia hexaptera Reich. Phytochem. Lett. 2014, 10, xcix-ciii. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A. Chemical Constituents of Bryophytes: Structures and Biological Activity. J. Nat. Prod. 2018, 81, 641–660. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.R.; Dorrestein, P.C.; Quinn, R.A. Illuminating the dark matter in metabolomics. Proc. Natl. Acad. Sci. USA 2015, 112, 12549–12550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S. Computational strategies for metabolite identification in metabolomics. Bioanalysis 2009, 1, 1579–1596. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Balcke, G.; Kleinenkuhnen, N.; Treutler, H.; Neumann, S. Untargeted In Silico Compound Classification—A Novel Metabolomics Method to Assess the Chemodiversity in Bryophytes. IJMS 2021, 22, 3251. [Google Scholar] [CrossRef]
- Ruttkies, C.; Schymanski, E.L.; Wolf, S.; Hollender, J.; Neumann, S. MetFrag relaunched: Incorporating strategies beyond in silico fragmentation. J. Cheminform. 2016, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.A.; Nothias, L.-F.; Ludwig, M.; Fleischauer, M.; Gentry, E.C.; Witting, M.; Dorrestein, P.C.; Dührkop, K.; Böcker, S. High-confidence structural annotation of metabolites absent from spectral libraries. Nat. Biotechnol. 2022, 40, 411–421. [Google Scholar] [CrossRef]
- Sedio, B.E. Recent breakthroughs in metabolomics promise to reveal the cryptic chemical traits that mediate plant community composition, character evolution and lineage diversification. New Phytol. 2017, 214, 952–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakawa, Y.; Ludwiczuk, A.; Nagashima, F.; Asakawa, Y.; Ludwiczuk, A.; Nagashima, F. Fortschritte der Chemie organischer Naturstoffe = Progress in the chemistry of organic natural products. In Chemical Constituents of Bryophytes: Bio- and Chemical Diversity, Biological Activity and Chemosystematics; Asakawa, Y., Ludwiczuk, A., Nagashima, F., Eds.; Springer: Wien, Austria; New York, NY, USA, 2013; ISBN 978-3-7091-1083-6. [Google Scholar]
- Kohn, G.; Vandekerkhove, O.; Hartmann, E.; Beutelmann, P. Acetylenic fatty acids in the Ricciaceae (Hepaticae). Phytochemistry 1988, 27, 1049–1051. [Google Scholar] [CrossRef]
- Markham, K.R.; J. Porter, L. Evidence of biosynthetic simplicity in the flavonoid chemistry of the Ricciaceae. Phytochemistry 1975, 14, 199–201. [Google Scholar] [CrossRef]
- Kunz, S.; Burkhardt, G.; Becker, H. Riccionidins a and b, anthocyanidins from the cell walls of the liverwort Ricciocarpos natans. Phytochemistry 1993, 35, 233–235. [Google Scholar] [CrossRef]
- Shaw, A.J.; Szovenyi, P.; Shaw, B. Bryophyte diversity and evolution: Windows into the early evolution of land plants. Am. J. Bot. 2011, 98, 352–369. [Google Scholar] [CrossRef]
- Peters, K.; Blatt-Janmaat, K.; Tkach, N.; Van Dam, N.M.; Neumann, S. Investigating untargeted metabolomics for its use in integrative taxonomy—Linking metabolomics, DNA marker-based se-quencing and bioimaging of phenotypes. Zenodo 2023. [Google Scholar] [CrossRef]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Rutz, A.; Sorokina, M.; Galgonek, J.; Willighagen, E.; Gaudry, A.; Graham, J.G.; Stephan, R.; Page, R.; Vondrášek, J.; Steinbeck, C.; et al. The LOTUS Initiative for Open Natural Products Research: Knowledge Management through Wikidata. BioRxiv 2021, 78. [Google Scholar]
- Nakamura, Y.; Mochamad Afendi, F.; Kawsar Parvin, A.; Ono, N.; Tanaka, K.; Hirai Morita, A.; Sato, T.; Sugiura, T.; Altaf-Ul-Amin, M.; Kanaya, S. KNApSAcK Metabolite Activity Database for Retrieving the Relationships Between Metabolites and Biological Activities. Plant Cell Physiol. 2014, 55, e7. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, J.; Swedlow, J.R.; Barlow, M.; Cook, C.E.; Sarkans, U.; Patwardhan, A.; Brazma, A.; Birney, E. A call for public archives for biological image data. Nat. Methods 2018, 15, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Löffler, F.; Wesp, V.; König-Ries, B.; Klan, F. Dataset search in biodiversity research: Do metadata in data repositories reflect scholarly information needs? PLoS ONE 2021, 16, e0246099. [Google Scholar] [CrossRef]
- Meijering, E.; Carpenter, A.E.; Peng, H.; Hamprecht, F.A.; Olivo-Marin, J.-C. Imagining the future of bioimage analysis. Nat. Biotechnol. 2016, 34, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Samuel, S.; Taubert, F.; Walther, D.; König-Ries, B.; Bücker, H.M. Towards Reproducibility of Microscopy Experiments. D-Lib Mag. 2017, 23, 245–253. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167. [Google Scholar] [CrossRef]
- Kommineni, V.K.; Tautenhahn, S.; Baddam, P.; Gaikwad, J.; Wieczorek, B.; Triki, A.; Kattge, J. Comprehensive leaf size traits dataset for seven plant species from digitised herbarium specimen images covering more than two centuries. BDJ 2021, 9, e69806. [Google Scholar] [CrossRef]
- Chen, D.; Shi, R.; Pape, J.-M.; Neumann, K.; Arend, D.; Graner, A.; Chen, M.; Klukas, C. Predicting plant biomass accumulation from image-derived parameters. GigaScience 2018, 7, giy001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, O.L.P.; Svenning, J.; Olsen, K.; Dupont, S.; Garner, B.H.; Iosifidis, A.; Price, B.W.; Høye, T.T. Species-level image classification with convolutional neural network enables insect identification from habitus images. Ecol. Evol. 2020, 10, 737–747. [Google Scholar] [CrossRef]
- Høye, T.T.; Ärje, J.; Bjerge, K.; Hansen, O.L.P.; Iosifidis, A.; Leese, F.; Mann, H.M.R.; Meissner, K.; Melvad, C.; Raitoharju, J. Deep learning and computer vision will transform entomology. Proc. Natl. Acad. Sci. USA 2021, 118, e2002545117. [Google Scholar] [CrossRef]
- Peters, K.; Gorzolka, K.; Bruelheide, H.; Neumann, S. Seasonal variation of secondary metabolites in nine different bryophytes. Ecol. Evol. 2018, 8, 9105–9117. [Google Scholar] [CrossRef] [PubMed]
- Jarmusch, S.A. Advancements in capturing and mining mass spectrometry data are transforming natural products research. Nat. Prod. Rep. 2021, 17, 2066–2082. [Google Scholar] [CrossRef]
- Peters, K.; Worrich, A.; Weinhold, A.; Alka, O.; Balcke, G.; Birkemeyer, C.; Bruelheide, H.; Calf, O.; Dietz, S.; Dührkop, K.; et al. Current Challenges in Plant Eco-Metabolomics. Int. J. Mol. Sci. 2018, 19, 1385. [Google Scholar] [CrossRef] [Green Version]
- Peters, K.; Gorzolka, K.; Bruelheide, H.; Neumann, S. Computational workflow to study the seasonal variation of secondary metabolites in nine different bryophytes. Sci. Data 2018, 5, 180179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haug, K.; Salek, R.M.; Conesa, P.; Hastings, J.; de Matos, P.; Rijnbeek, M.; Mahendraker, T.; Williams, M.; Neumann, S.; Rocca-Serra, P.; et al. MetaboLights—An open-access general-purpose repository for metabolomics studies and associated meta-data. Nucleic Acids Res. 2013, 41, D781–D786. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.W.N.; Alexander, J.M.; Allard, P.; Baines, O.; Baldy, V.; Bardgett, R.D.; Capdevila, P.; Coley, P.D.; David, B.; Defossez, E.; et al. Functional Traits 2.0: The power of the metabolome for ecology. J. Ecol. 2022, 110, 4–20. [Google Scholar] [CrossRef]
- Renner, M.A. Opportunities and challenges presented by cryptic bryophyte species. Telopea 2020, 23, 41–60. [Google Scholar] [CrossRef]
- Shaw, J. Biogeographic patterns and cryptic speciation in bryophytes: Cryptic speciation in bryophytes. J. Biogeogr. 2001, 28, 253–261. [Google Scholar] [CrossRef]
- Djoumbou Feunang, Y.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated chemical classification with a comprehensive, computable taxonomy. J. Cheminformatics 2016, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Soriano, G.; Del-Castillo-Alonso, M.-Á.; Monforte, L.; Tomás-Las-Heras, R.; Martínez-Abaigar, J.; Núñez-Olivera, E. Developmental Stage Determines the Accumulation Pattern of UV-Absorbing Compounds in the Model Liverwort Marchantia polymorpha subsp. ruderalis under Controlled Conditions. Plants 2021, 10, 473. [Google Scholar] [CrossRef]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef]
- Shahaf, N.; Rogachev, I.; Heinig, U.; Meir, S.; Malitsky, S.; Battat, M.; Wyner, H.; Zheng, S.; Wehrens, R.; Aharoni, A. The WEIZMASS spectral library for high-confidence metabolite identification. Nat. Commun. 2016, 7, 12423. [Google Scholar] [CrossRef] [Green Version]
- Stelmasiewicz, M.; Świątek, Ł.; Ludwiczuk, A. Phytochemical Profile and Anticancer Potential of Endophytic Microorganisms from Liverwort Species, Marchantia polymorpha L. Molecules 2021, 27, 153. [Google Scholar] [CrossRef]
- Wangikar, H.; Chavan, S.J.; Bankar, P.; Gavali, P.; Taware, T. Analysis and fungal Isolation of some mosses, Riccia discolor and Targionia hyophylla from Baramati, district-Pune, Maharashtra, India. Int. J. Bot. Stud. 7 2021, 6, 37–42. [Google Scholar]
- Wankhede Tb, W.T. Mycorrhization in bryophyte riccia discolor lehm. et. lindenb. IJRBAT 2017, 5, 120–127. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Bottcher, C.; Neumann, S. Highly sensitive feature detection for high resolution LC/MS. BMC Bioinform. 2008, 9, 504. [Google Scholar] [CrossRef] [Green Version]
- Klavina, L. A study on bryophyte chemical composition–search for new applications. Agron. Res. 2015, 13, 969–978. [Google Scholar]
- Uthe, H.; van Dam, N.M.; Hervé, M.R.; Sorokina, M.; Peters, K.; Weinhold, A. A practical guide to implementing metabolomics in plant ecology and biodiversity research. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2020; p. S0065229620300732. [Google Scholar]
- Sabovljević, M.S.; Sabovljević, A.D.; Ikram, N.K.K.; Peramuna, A.; Bae, H.; Simonsen, H.T. Bryophytes—An emerging source for herbal remedies and chemical production. Plant Genet. Resour. 2016, 14, 314–327. [Google Scholar] [CrossRef]
- Khalkar, K.M.; Kadam, V.B. Biochemical Evaluation of Some Liverworts Pigments and Phenolics. J. Drug Delivery Ther. 2021, 11, 78–80. [Google Scholar] [CrossRef]
- van Dam, N.M.; van der Meijden, E. A Role for Metabolomics in Plant Ecology. In Annual Plant Reviews Volume 43; Hall, R.D., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 87–107. ISBN 978-1-4443-3995-6. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification And Direct Sequencing Of Fungal Ribosomal RNA Genes For Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Vanderpoorten, A.; Quandt, D.; Goffinet, B. Utility of the Internal Transcribed Spacers of the 18S-5.8S-26S Nuclear Ribosomal DNA in Land Plant Systematics with Special Emphasis on Bryophytes. In Plant Genome: Biodiversity and Evolution—Volume 2, Part B; Science Publishers: New York, NY, USA, 2006; pp. 385–407. [Google Scholar]
- Forrest, L.L.; Crandall-Stotler, B.J. A phylogeny of the simple thalloid liverworts (Junger-manniopsida, subclass Metzgeriidae) as inferred from five chloroplast genes. Monogr. Syst. Bot. Mo. Bot. Gard. 2004, 98, 119–140. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In 2010 Gateway Computing Environments Workshop (GCE); IEEE: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Peters, K.; König-Ries, B. Reference bioimaging to assess the phenotypic trait diversity of bryophytes within the family Scapaniaceae. Sci. Data 2022, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Pau, G.; Fuchs, F.; Sklyar, O.; Boutros, M.; Huber, W. EBImage--an R package for image processing with applications to cellular phenotypes. Bioinformatics 2010, 26, 979–981. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.; Moore, J.; Li, S.W.; Rustici, G.; Tarkowska, A.; Chessel, A.; Leo, S.; Antal, B.; Ferguson, R.K.; Sarkans, U.; et al. Image Data Resource: A bioimage data integration and publication platform. Nat. Methods 2017, 14, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, K. Data Integration in Biodiversity—A Principal Investigation on Three Liverwort Species of Riccia Integrating Metabolomics, Sequencing and Phenotypic Data for Use in Integrative Taxonomy; University of Dundee: Dundee, UK, 2022. [Google Scholar]
- Böttcher, C.; Westphal, L.; Schmotz, C.; Prade, E.; Scheel, D.; Glawischnig, E. The Multifunctional Enzyme CYP71B15 (PHYTOALEXIN DEFICIENT3) Converts Cysteine-Indole-3-Acetonitrile to Camalexin in the Indole-3-Acetonitrile Metabolic Network of Arabidopsis thaliana. Plant Cell. Online 2009, 21, 1830–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Eiriksson, F.F.; Thorsteinsdóttir, M.; Simonsen, H.T. Effects of extraction parameters on lipid profiling of mosses using UPLC-ESI-QTOF-MS and multivariate data analysis. Metabolomics 2021, 17, 96. [Google Scholar] [CrossRef]
- Blatt-Janmaat, K.L.; Neumann, S.; Schmidt, F.; Ziegler, J.; Peters, K.; Qu, Y. Impact of in vitro hormone treatments on the bibenzyl production of Radula complanata. Botany 2022. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Spicer, R.A.; Salek, R.; Steinbeck, C. Compliance with minimum information guidelines in public metabolomics repositories. Sci. Data 2017, 4, 170137. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing Mass Spectrometry Data for Metabolite Profiling Using Nonlinear Peak Alignment, Matching, and Identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Dührkop, K.; Nothias, L.-F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C.; et al. Systematic classification of unknown metabolites using high-resolution fragmentation mass spectra. Nat. Biotechnol. 2020, 39, 462–471. [Google Scholar] [CrossRef]
- Bento, A.P.; Hersey, A.; Félix, E.; Landrum, G.; Gaulton, A.; Atkinson, F.; Bellis, L.J.; De Veij, M.; Leach, A.R. An open source chemical structure curation pipeline using RDKit. J. Cheminform 2020, 12, 51. [Google Scholar] [CrossRef]
- Peters, K.; Poeschl, Y.; Blatt-Janmaat, K.L.; Uthe, H. Ecometabolomics Studies of Bryophytes. In Bioactive Compounds in Bryophytes and Pteridophytes; Murthy, H.N., Ed.; Reference Series in Phytochemistry; Springer International Publishing: Cham, Germany, 2022; pp. 1–43. ISBN 978-3-030-97415-2. [Google Scholar]
- Peters, K. Chemical Diversity and Classification of Secondary Metabolites in Nine Bryophyte Species. Metabolites 2019, 9, 222. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Grau, J.; Grosse, I.; Keilwagen, J. PRROC: Computing and visualizing precision-recall and receiver operating characteristic curves in R. Bioinformatics 2015, 31, 2595–2597. [Google Scholar] [CrossRef] [PubMed]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Tharwat, A. Classification assessment methods. ACI 2020, 17, 68–192. [Google Scholar] [CrossRef]
- Peters, K. iESTIMATE Computational Analysis Framework for Eco-Metabolomics Data Version 0.4. Zenodo. 2023. Available online: https://zenodo.org/record/7615220#.Y-xNKHZByUk (accessed on 12 February 2023).
- Stanton, D.E.; Coe, K.K. 500 million years of charted territory: Functional ecological traits in bryophytes. BDE 2021, 43, 234–252. [Google Scholar] [CrossRef]
- Price, S.A.; Schmitz, L. A promising future for integrative biodiversity research: An increased role of scale-dependency and functional biology. Phil. Trans. R. Soc. B 2016, 371, 20150228. [Google Scholar] [CrossRef] [Green Version]
- Goble, C.; Cohen-Boulakia, S.; Soiland-Reyes, S.; Garijo, D.; Gil, Y.; Crusoe, M.R.; Peters, K.; Schober, D. FAIR Computational Workflows. Data Intell. 2020, 2, 108–121. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.-W.; da Silva Santos, L.B.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef] [Green Version]
- Heberling, J.M.; Miller, J.T.; Noesgaard, D.; Weingart, S.B.; Schigel, D. Data integration enables global biodiversity synthesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2018093118. [Google Scholar] [CrossRef] [PubMed]
- König, C.; Weigelt, P.; Schrader, J.; Taylor, A.; Kattge, J.; Kreft, H. Biodiversity data integration—The significance of data resolution and domain. PLoS Biol. 2019, 17, e3000183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arribas, P.; Andújar, C.; Bidartondo, M.I.; Bohmann, K.; Coissac, É.; Creer, S.; deWaard, J.R.; Elbrecht, V.; Ficetola, G.F.; Goberna, M.; et al. Connecting high-throughput biodiversity inventories: Opportunities for a site-based genomic framework for global integration and synthesis. Mol. Ecol. 2021, 30, 1120–1135. [Google Scholar] [CrossRef] [PubMed]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peters, K.; Blatt-Janmaat, K.L.; Tkach, N.; van Dam, N.M.; Neumann, S. Untargeted Metabolomics for Integrative Taxonomy: Metabolomics, DNA Marker-Based Sequencing, and Phenotype Bioimaging. Plants 2023, 12, 881. https://doi.org/10.3390/plants12040881
Peters K, Blatt-Janmaat KL, Tkach N, van Dam NM, Neumann S. Untargeted Metabolomics for Integrative Taxonomy: Metabolomics, DNA Marker-Based Sequencing, and Phenotype Bioimaging. Plants. 2023; 12(4):881. https://doi.org/10.3390/plants12040881
Chicago/Turabian StylePeters, Kristian, Kaitlyn L. Blatt-Janmaat, Natalia Tkach, Nicole M. van Dam, and Steffen Neumann. 2023. "Untargeted Metabolomics for Integrative Taxonomy: Metabolomics, DNA Marker-Based Sequencing, and Phenotype Bioimaging" Plants 12, no. 4: 881. https://doi.org/10.3390/plants12040881
APA StylePeters, K., Blatt-Janmaat, K. L., Tkach, N., van Dam, N. M., & Neumann, S. (2023). Untargeted Metabolomics for Integrative Taxonomy: Metabolomics, DNA Marker-Based Sequencing, and Phenotype Bioimaging. Plants, 12(4), 881. https://doi.org/10.3390/plants12040881