

Variation in Photosynthetic Efficiency under Fluctuating Light between Rose Cultivars and its Potential for Improving Dynamic Photosynthesis

Abstract
:1. Introduction
2. Results
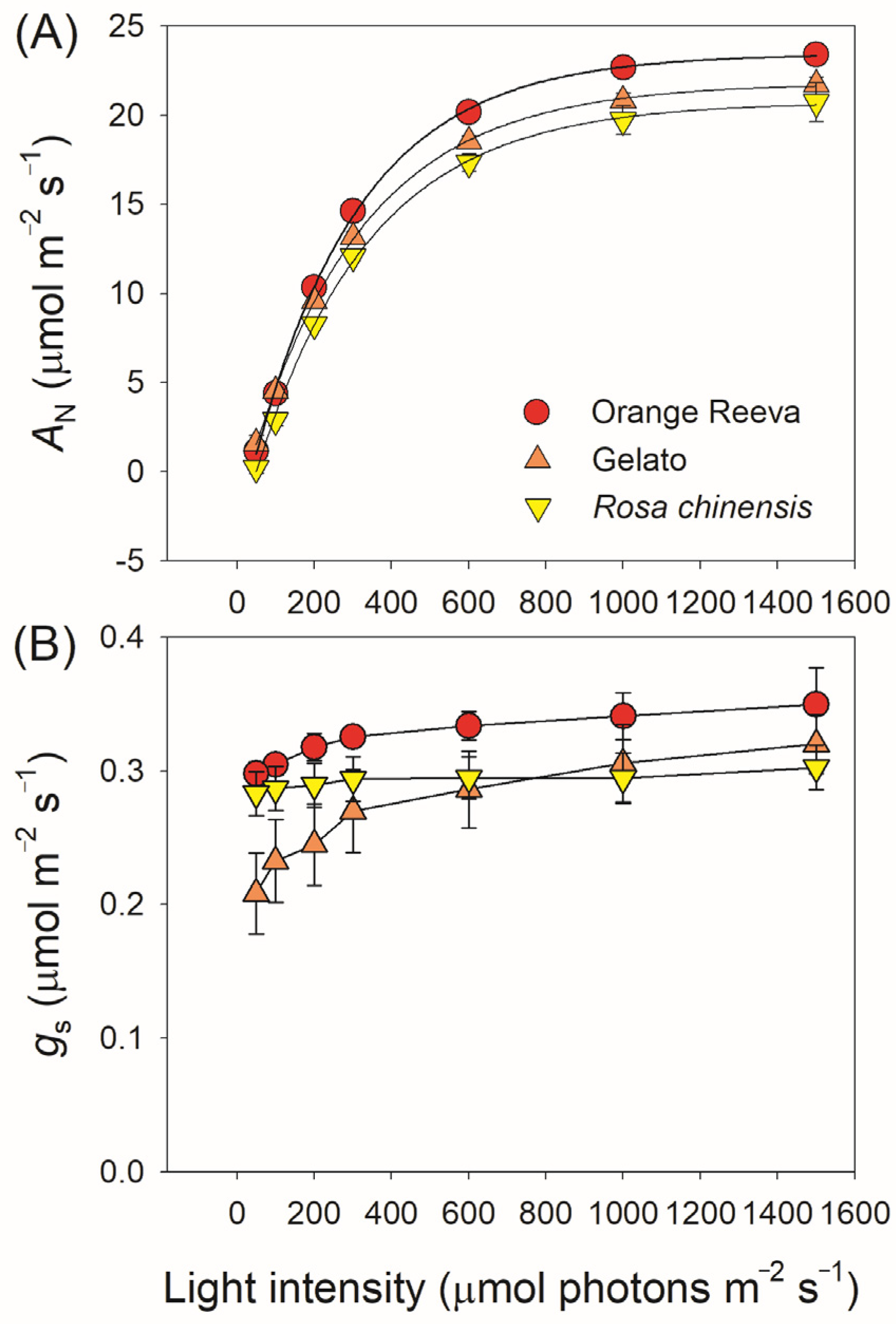
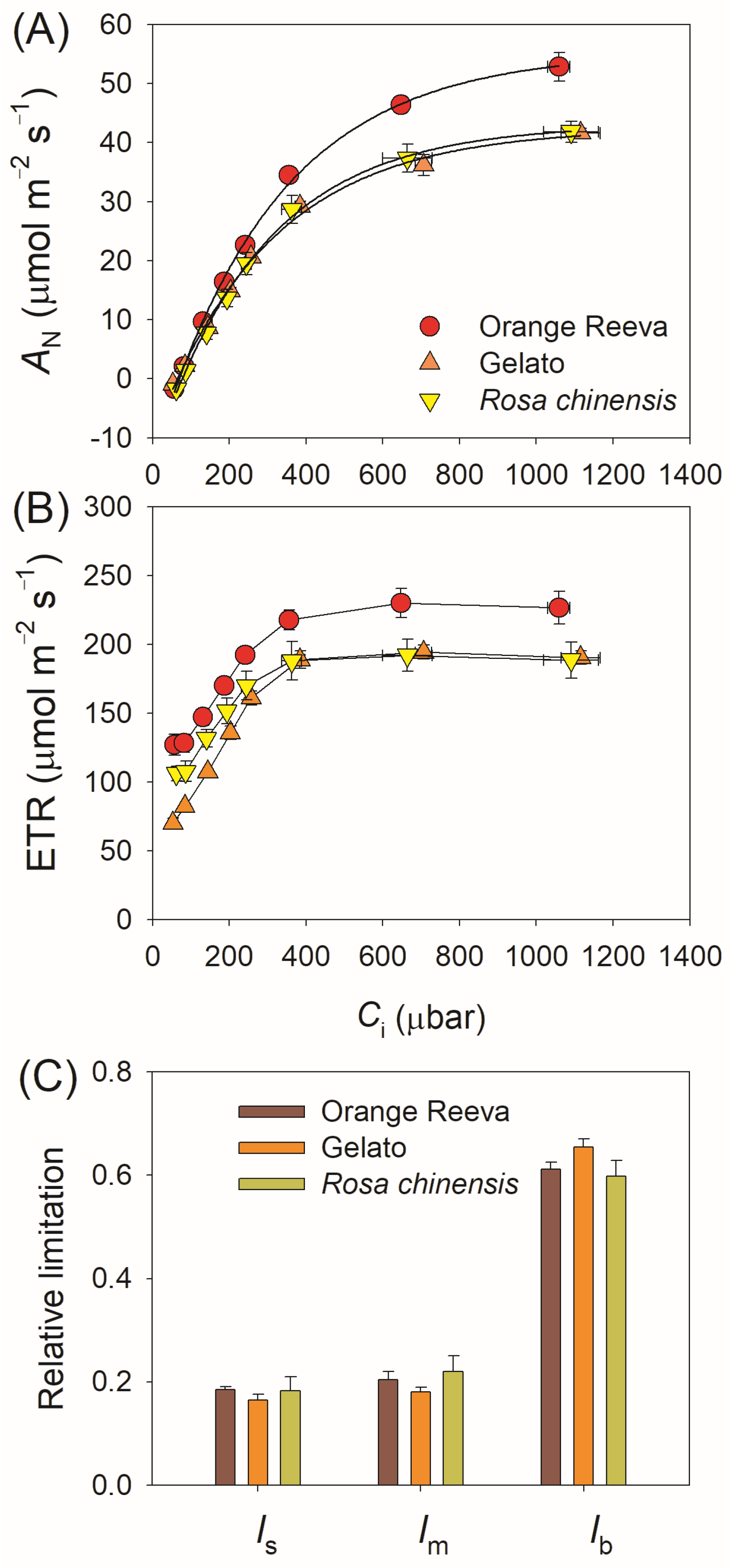
2.1. Photosynthetic Characteristics under Steady-State Differ Slightly between Rose Genotypes
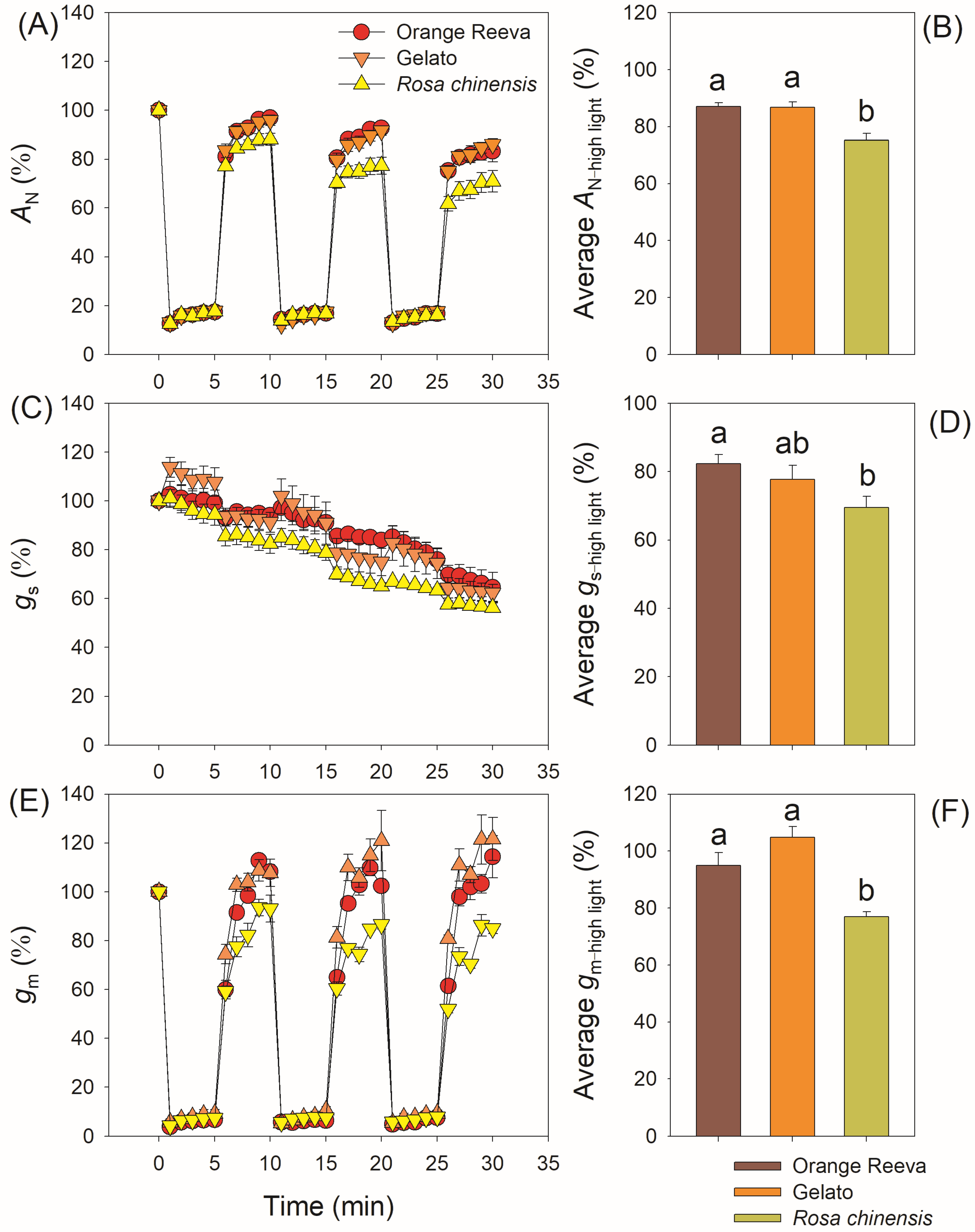
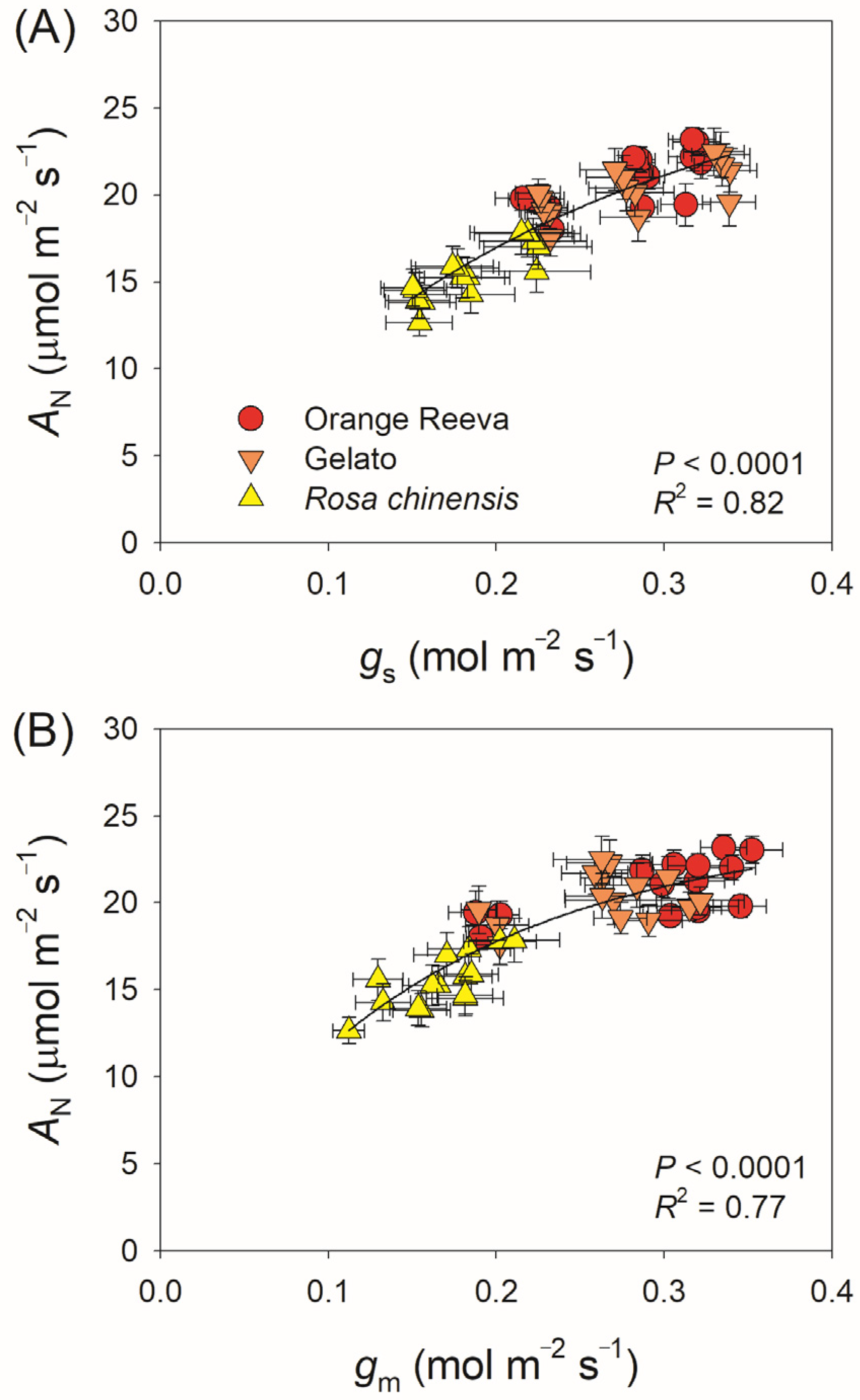
2.2. Modern Rose Cultivars Use Fluctuating Light More Efficiently Than the Old Rose Species
3. Discussion
3.1. Steady-State Photosynthesis across Rose Germplasm Is Mainly Limited by Biochemical Capacity
3.2. Modern Rose cultivars have Stronger Dynamic Photosynthetic Efficiency Than the Old Rose Rosa chinensis
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Gas Exchange and Chlorophyll Fluorescence Measurements
4.3. Calculations of gm, Cc and Vcmax
4.4. Quantitative Limitation Analysis of AN
4.5. SPAD Index and Leaf Nutrient Content Measurements
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Flexas, J.; Niinemets, Ü.; Gallé, A.; Barbour, M.M.; Centritto, M.; Diaz-Espejo, A.; Douthe, C.; Galmés, J.; Ribas-Carbo, M.; Rodriguez, P.L.; et al. Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef]
- Carriquí, M.; Cabrera, H.M.; Conesa, M.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Galmés, J.; Ribas-Carbo, M.; Tomás, M.; et al. Diffusional limitations explain the lower photosynthetic capacity of ferns as compared with angiosperms in a common garden study. Plant Cell Environ. 2015, 38, 448–460. [Google Scholar] [CrossRef]
- Yamori, W.; Nagai, T.; Makino, A. The rate-limiting step for CO2 assimilation at different temperatures is influenced by the leaf nitrogen content in several C3 crop species. Plant Cell Environ. 2011, 34, 764–777. [Google Scholar] [CrossRef]
- Flexas, J.; Díaz-Espejo, A.; Conesa, M.A.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Galmés, J.; Medrano, H.; Ribas-Carbo, M.; et al. Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant Cell Environ. 2016, 39, 965–982. [Google Scholar] [CrossRef]
- Campany, C.E.; Tjoelker, M.G.; von Caemmerer, S.; Duursma, R.A. Coupled response of stomatal and mesophyll conductance to light enhances photosynthesis of shade leaves under sunflecks. Plant Cell Environ. 2016, 39, 2762–2773. [Google Scholar] [CrossRef]
- Xiong, D.; Douthe, C.; Flexas, J. Differential coordination of stomatal conductance, mesophyll conductance, and leaf hydraulic conductance in response to changing light across species. Plant. Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Sisó, S.; Flexas, J.; Galmés, J.; García-Nogales, A.; Niinemets, Ü.; Sancho-Knapik, D.; Saz, M.Á.; Gil-Pelegrín, E. Cell-level anatomical characteristics explain high mesophyll conductance and photosynthetic capacity in sclerophyllous Mediterranean oaks. New Phytol. 2017, 214, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yang, Y.-J.; Wang, J.-H.; Hu, H. Photorespiration is the major alternative electron sink under high light in alpine evergreen sclerophyllous Rhododendron species. Plant Sci. 2019, 289, 110275. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-H.; Huang, W.; Yang, Q.-Y.; Chang, W.; Zhang, S.-B. Anatomical and diffusional determinants inside leaves explain the difference in photosynthetic capacity between Cypripedium and Paphiopedilum, Orchidaceae. Photosynth. Res. 2018, 136, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Pearcy, R.W. Sunflecks and photosynthesis in plant canopies. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 421–453. [Google Scholar] [CrossRef]
- Slattery, R.A.; Walker, B.J.; Weber, A.P.M.; Ort, D.R. The impacts of fluctuating light on crop performance. Plant Physiol. 2018, 176, 990–1003. [Google Scholar] [CrossRef] [Green Version]
- Adachi, S.; Tanaka, Y.; Miyagi, A.; Kashima, M.; Tezuka, A.; Toya, Y.; Kobayashi, S.; Ohkubo, S.; Shimizu, H.; Kawai-Yamada, M.; et al. High-yielding rice Takanari has superior photosynthetic response to a commercial rice Koshihikari under fluctuating light. J. Exp. Bot. 2019, 70, 5287–5297. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Adachi, S.; Yamori, W. Natural genetic variation of the photosynthetic induction response to fluctuating light environment. Curr. Opin. Plant Biol. 2019, 49, 52–59. [Google Scholar] [CrossRef]
- Acevedo-Siaca, L.G.; Coe, R.; Wang, Y.; Kromdijk, J.; Quick, W.P.; Long, S.P. Variation in photosynthetic induction between rice accessions and its potential for improving productivity. New Phytol. 2020, 227, 1097–1108. [Google Scholar] [CrossRef] [Green Version]
- De Souza, A.P.; Wang, Y.; Orr, D.J.; Carmo-Silva, E.; Long, S.P. Photosynthesis across African cassava germplasm is limited by Rubisco and mesophyll conductance at steady state, but by stomatal conductance in fluctuating light. New Phytol. 2020, 225, 2498–2512. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Hashimoto-Sugimoto, M.; Iba, K.; Terashima, I.; Yamori, W. Improved stomatal opening enhances photosynthetic rate and biomass production in fluctuating light. J. Exp. Bot. 2020, 71, 2339–2350. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, J.; Estavillo, G.M.; Luo, T.; Hu, L. Leaf N content regulates the speed of photosynthetic induction under fluctuating light among canola genotypes (Brassica napus L.). Physiol. Plant. 2021, 172, 1844–1852. [Google Scholar] [CrossRef]
- Salter, W.T.; Merchant, A.M.; Richards, R.A.; Trethowan, R.; Buckley, T.N. Rate of photosynthetic induction in fluctuating light varies widely among genotypes of wheat. J. Exp. Bot. 2019, 70, 2787–2796. [Google Scholar] [CrossRef]
- Soleh, M.A.; Tanaka, Y.; Nomoto, Y.; Iwahashi, Y.; Nakashima, K.; Fukuda, Y.; Long, S.P.; Shiraiwa, T. Factors underlying genotypic differences in the induction of photosynthesis in soybean [Glycine max (L.) Merr.]. Plant. Cell Environ. 2016, 39, 685–693. [Google Scholar] [CrossRef]
- Li, Y.-T.; Luo, J.; Liu, P.; Zhang, Z.-S. C4 species utilize fluctuating light less efficiently than C3 species. Plant Physiol. 2021, 187, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Kusumi, K.; Iba, K.; Terashima, I. Increased stomatal conductance induces rapid changes to photosynthetic rate in response to naturally fluctuating light conditions in rice. Plant. Cell Environ. 2020, 43, 1230–1240. [Google Scholar] [CrossRef]
- Sakoda, K.; Yamori, W.; Groszmann, M.; Evans, J.R. Stomatal, mesophyll conductance, and biochemical limitations to photosynthesis during induction Research Article. Plant Physiol. 2021, 185, 146–160. [Google Scholar] [CrossRef]
- Liu, T.; Barbour, M.M.; Yu, D.; Rao, S.; Song, X. Mesophyll conductance exerts a significant limitation on photosynthesis during light induction. New Phytol. 2022, 233, 360–372. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Y.-Q.; Zhang, S.; Huang, W. Photosynthetic Induction Under Fluctuating Light Is Affected by Leaf Nitrogen Content in Tomato. Front. Plant Sci. 2022, 13, 835571. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- South, P.F.; Cavanagh, A.P.; Liu, H.W.; Ort, D.R. Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field. Science 2019, 363, eaat9077. [Google Scholar] [CrossRef] [Green Version]
- Simkin, A.J.; López-Calcagno, P.E.; Raines, C.A. Feeding the world: Improving photosynthetic efficiency for sustainable crop production. J. Exp. Bot. 2019, 70, 1119–1140. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.G.; Ort, D.R.; Whitmarsh, J.; Long, S.P. The slow reversibility of photosystem II thermal energy dissipation on transfer from high to low light may cause large losses in carbon gain by crop canopies: A theoretical analysis. J. Exp. Bot. 2004, 55, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.H.; Long, S.P. Slow induction of photosynthesis on shade to sun transitions in wheat may cost at least 21% of productivity. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160543. [Google Scholar] [CrossRef] [Green Version]
- Papanatsiou, M.; Petersen, J.; Henderson, L.; Wang, Y.; Christie, J.M.; Blatt, M.R. Optogenetic manipulation of stomatal kinetics improves carbon assimilation, water use, and growth. Science 2019, 363, 1456–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masumoto, C.; Ishii, T.; Hatanaka, T.; Uchida, N. Mechanism of High Photosynthetic Capacity in BC 2 F 4 LinesDerived from a Cross between Oryza sativa and Wild Relatives O. rufipogon. Plant Prod. Sci. 2005, 8, 539–545. [Google Scholar] [CrossRef]
- Driever, S.M.; Lawson, T.; Andralojc, P.J.; Raines, C.A.; Parry, M.A.J. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes. J. Exp. Bot. 2014, 65, 4959–4973. [Google Scholar] [CrossRef] [Green Version]
- Parry, M.A.J.; Madgwick, P.J.; Carvalho, J.F.C.; Andralojc, P.J. Paper Presented at International Workshop on Increasing Wheat Yield Potential, Cimmyt, Obregon, Mexico, 20–24 March 2006 Prospects for increasing photosynthesis by overcoming the limitations of Rubisco. J. Agric. Sci. 2007, 145, 31–43. [Google Scholar] [CrossRef]
- Whitney, S.M.; Houtz, R.L.; Alonso, H. Advancing Our Understanding and Capacity to Engineer Nature’s CO2-Sequestering Enzyme, Rubisco. Plant Physiol. 2011, 155, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Carmo-Silva, E.; Scales, J.C.; Madgwick, P.J.; Parry, M.A.J. Optimizing Rubisco and its regulation for greater resource use efficiency. Plant. Cell Environ. 2015, 38, 1817–1832. [Google Scholar] [CrossRef]
- Ohkubo, S.; Tanaka, Y.; Yamori, W.; Adachi, S. Rice Cultivar Takanari Has Higher Photosynthetic Performance Under Fluctuating Light Than Koshihikari, Especially Under Limited Nitrogen Supply and Elevated CO2. Front. Plant Sci. 2020, 11, 1308. [Google Scholar] [CrossRef]
- Matthews, J.S.; Vialet-Chabrand, S.; Lawson, T. Acclimation to Fluctuating Light Impacts the Rapidity of Response and Diurnal Rhythm of Stomatal Conductance. Plant Physiol. 2018, 176, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Loriaux, S.D.; Avenson, T.J.; Welles, J.M.; Mcdermitt, D.K.; Eckles, R.D.; Riensche, B.; Genty, B. Closing in on maximum yield of chlorophyll fluorescence using a single multiphase flash of sub-saturating intensity. Plant Cell Environ. 2013, 36, 1755–1770. [Google Scholar] [CrossRef] [PubMed]
- Harbinson, J.; Genty, B.; Baker, N.R. Relationship between the Quantum Efficiencies of Photosystems I and II in Pea Leaves. Plant Physiol. 1989, 90, 1029–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krall, J.P.; Edwards, G.E. Relationship between photosystem II activity and CO2 fixation in leaves. Physiol. Plant. 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Yin, X.; Struik, P.C.; Romero, P.; Harbinson, J.; Evers, J.B.; Van Der Putten, P.E.L.; Vos, J. Using combined measurements of gas exchange and chlorophyll fluorescence to estimate parameters of a biochemical C photosynthesis model: A critical appraisal and a new integrated approach applied to leaves in a wheat (Triticum aestivum) canopy. Plant. Cell Environ. 2009, 32, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.R.; Dreyer, E. Temperature response of photosynthesis and internal conductance to CO2: Results from two independent approaches. J. Exp. Bot. 2006, 57, 3057–3067. [Google Scholar] [CrossRef] [Green Version]
- Eyland, D.; van Wesemael, J.; Lawson, T.; Carpentier, S. The impact of slow stomatal kinetics on photosynthesis and water use efficiency under fluctuating light. Plant Physiol. 2021, 186, 998–1012. [Google Scholar] [CrossRef]
- Hermida-Carrera, C.; Kapralov, M.V.; Galmés, J. Rubisco Catalytic Properties and Temperature Response in Crops. Plant Physiol. 2016, 171, 2549–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Orange Reeva | Gelato | Rosa chinensis |
|---|---|---|---|
| SPAD | 51.4 ± 0.48 a | 53.1 ± 0.81 a | 46.9 ± 0.39 b |
| LMA (g m−2) | 58.04 ± 2.3 a | 48.0 ± 0.83 b | 53.02 ± 0.93 c |
| Leaf N content (mg/g) | 43.2 ± 0.72 a | 40.5 ± 1.4 a | 26.4 ± 1.7 c |
| Leaf K content (mg/g) | 22.2 ± 0.7 a | 22.3 ± 0.92 a | 12.4 ± 0.30 b |
| Leaf P content (mg/g) | 5.76 ± 0.06 a | 5.08 ± 0.04 b | 3.35 ± 0.18 c |
| AN (μmol m−2 s−1) | 23.9 ± 0.4 a | 21.7 ± 0.4 b | 20.7 ± 1.1 b |
| gs (mol m−2 s−1) | 0.35 ± 0.03 a | 0.32 ± 0.02 a | 0.30 ± 0.02 a |
| gm (mol m−2 s−1) | 0.31 ± 0.03 a | 0.27 ± 0.03 a | 0.19 ± 0.02 b |
| Rd (μmol m−2 s−1) | 1.18 ± 0.04 a | 1.00 ± 0.04 a | 1.22 ± 0.06 a |
| Vcmax (μmol m−2 s−1) | 123 ± 5.6a | 97.5 ± 2.7b | 98.6 ± 4.4b |
| Jmax (μmol m−2 s−1) | 130 ± 8.0a | 99.8 ± 4.7b | 101 ± 4.5b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-Q.; Zeng, Z.-L.; Shi, Z.-M.; Wang, J.-H.; Huang, W. Variation in Photosynthetic Efficiency under Fluctuating Light between Rose Cultivars and its Potential for Improving Dynamic Photosynthesis. Plants 2023, 12, 1186. https://doi.org/10.3390/plants12051186
Wang X-Q, Zeng Z-L, Shi Z-M, Wang J-H, Huang W. Variation in Photosynthetic Efficiency under Fluctuating Light between Rose Cultivars and its Potential for Improving Dynamic Photosynthesis. Plants. 2023; 12(5):1186. https://doi.org/10.3390/plants12051186
Chicago/Turabian StyleWang, Xiao-Qian, Zhi-Lan Zeng, Zi-Ming Shi, Ji-Hua Wang, and Wei Huang. 2023. "Variation in Photosynthetic Efficiency under Fluctuating Light between Rose Cultivars and its Potential for Improving Dynamic Photosynthesis" Plants 12, no. 5: 1186. https://doi.org/10.3390/plants12051186
APA StyleWang, X. -Q., Zeng, Z. -L., Shi, Z. -M., Wang, J. -H., & Huang, W. (2023). Variation in Photosynthetic Efficiency under Fluctuating Light between Rose Cultivars and its Potential for Improving Dynamic Photosynthesis. Plants, 12(5), 1186. https://doi.org/10.3390/plants12051186