
The Evolution of Agrarian Landscapes in the Tropical Andes

 , ,
, ,
Abstract
:1. Introduction
1.1. An Historical Overview of Early Settlement and Cultivation
1.2. Human Responses to Holocene Climate Change and the Temporal Scale of Study
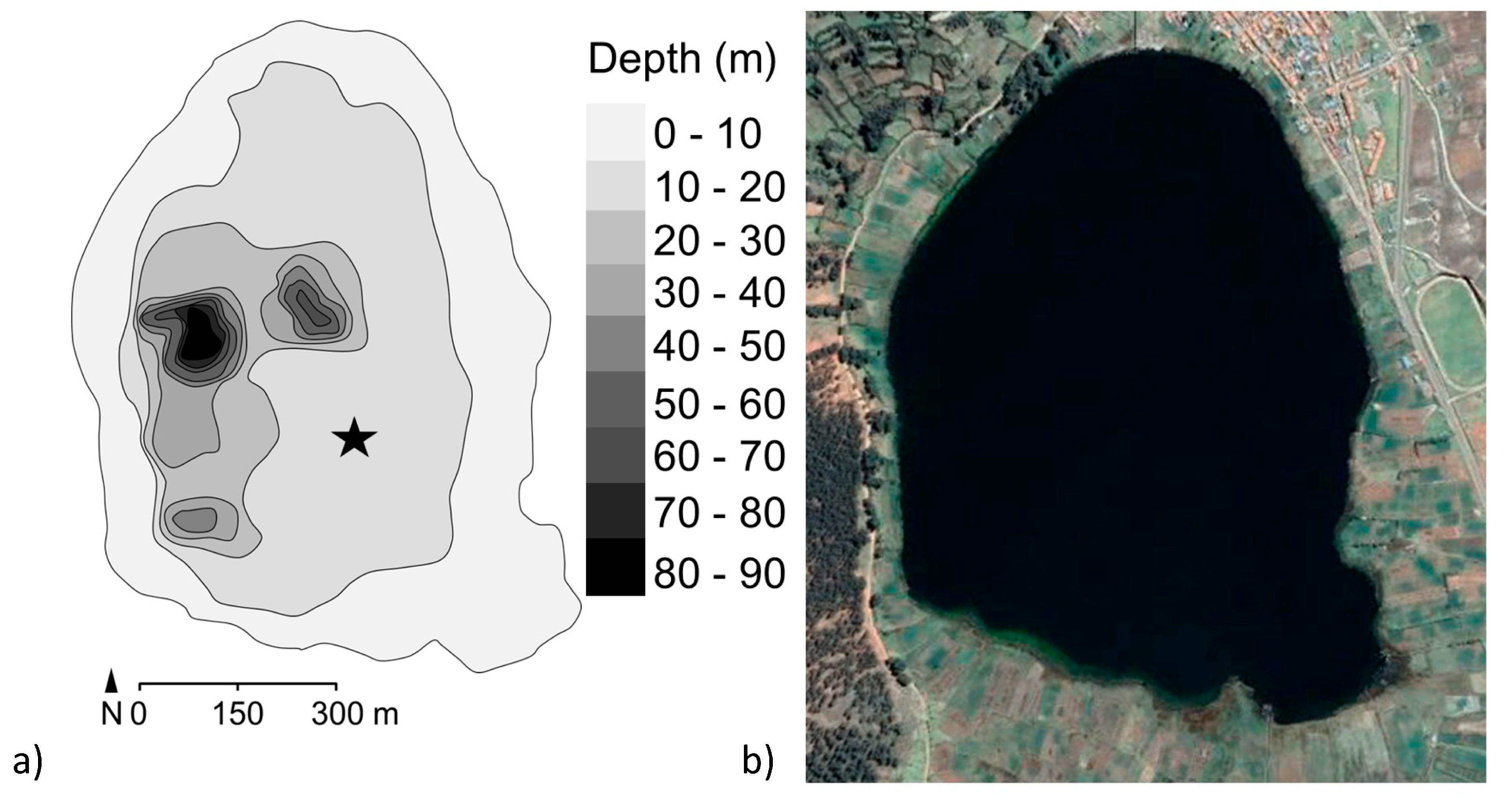
2. Site Description, Materials and Methods
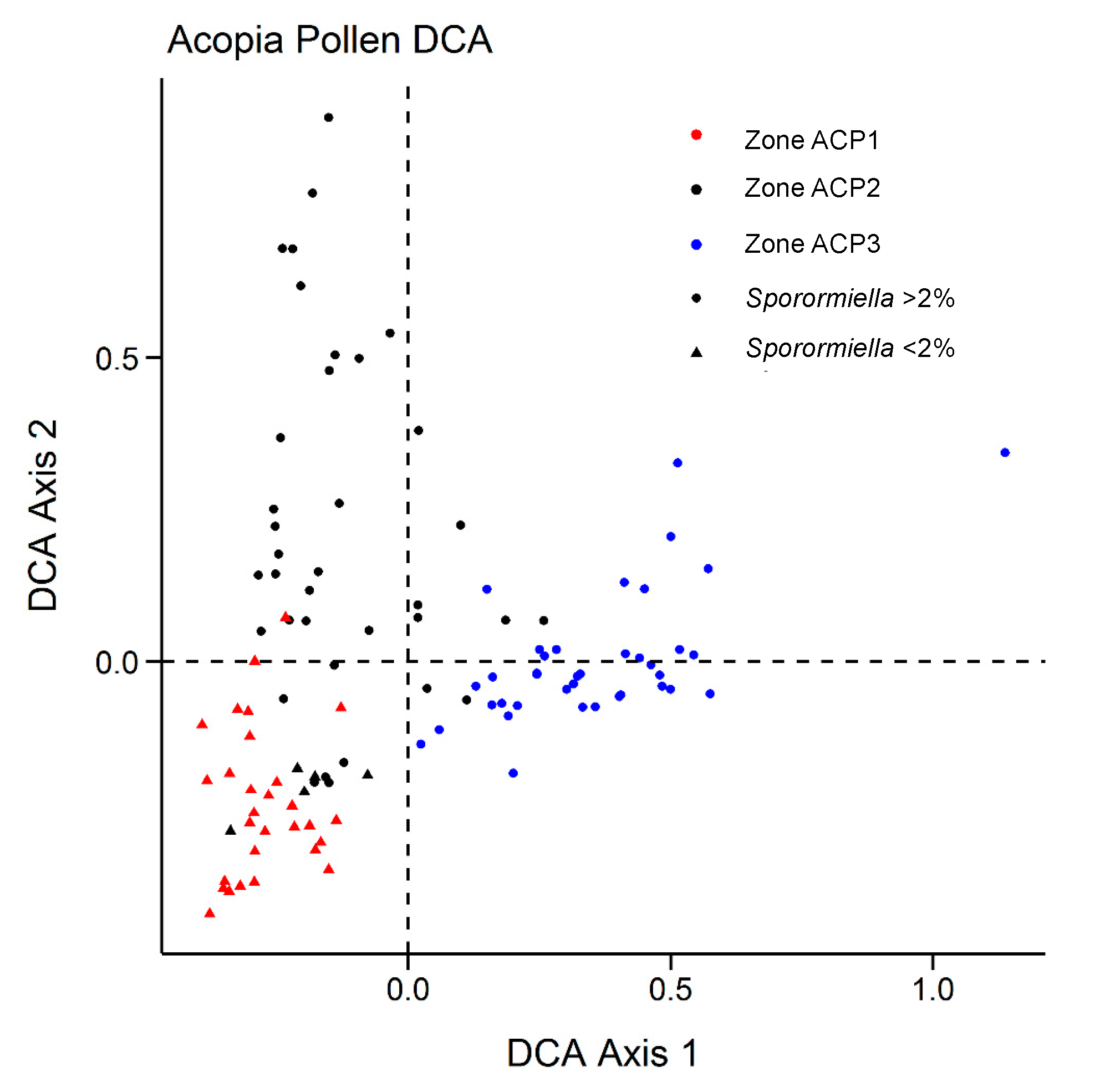
3. Results
- ACP–1 (768–543 cm; c. 7100–4450 cal BP)
- ACP–2 (542–139 cm; c. 4450–2090 cal BP)
- ACP–3 (138–0 cm; c. 2090–0 cal BP)
4. Discussion
4.1. How Did Climate Change Influence Land Use around Lake Acopia?
4.2. Can We Detect Adaptations to Environmental Change in the Way the Land Was Used?
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Echeverry, J.; Leiton, M. State of the landscape and dynamics of loss and fragmentation of forest critically endangered in the tropical Andes hotspot: Implications for conservation planning. J. Landsc. Ecol. 2021, 14, 73–91. [Google Scholar] [CrossRef]
- Fernández, J.; Markgraf, V.; Panarello, H.O.; Albero, M.; Angiolini, F.E.; Valencio, S.; Arriaga, M. Late Pleistocene/Early Holocene environments and climates, fauna, and human occupation in the Argentine Altiplano. Geoarchaeology 1991, 6, 251–272. [Google Scholar] [CrossRef]
- Brunschön, C.; Behling, H. Reconstruction and visualization of upper forest line and vegetation changes in the Andean depression region of southeastern Ecuador since the last glacial maximum—A multi-site synthesis. Rev. Palaeobot. Palynol. 2010, 163, 139–152. [Google Scholar] [CrossRef]
- Raczka, M.F.; Mosblech, N.A.; Giosan, L.; Folcik, A.M.; Valencia, B.G.; Kingston, M.; Baskin, S.; Bush, M.B. A human role in Andean megafaunal extinction? Quat. Sci. Rev. 2019, 205, 154–165. [Google Scholar] [CrossRef]
- Rozas-Davila, A.; Rodbell, D.; Bush, M.B. Pleistocene megafaunal extinction in the grasslands of Junin-Peru. Quat. Res. 2023, 50, 755–766. [Google Scholar] [CrossRef]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Alta Mira Press: Lanham, MD, USA, 2006; p. 304. [Google Scholar]
- Piperno, D.R. The Origins of Plant Cultivation and Domestication in the Neotropics. In Foraging Theory and the Transition to Agriculture; Kennett, D., Winterhalder, B., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 137–166. [Google Scholar]
- McMichael, C.; Witteveen, N.; Scholz, S.; Zwier, M.; Prins, M.; Lougheed, B.; Mothes, P.; Gosling, W. 30,000 years of landscape and vegetation dynamics in a mid-elevation Andean valley. Quat. Sci. Rev. 2021, 258, 106866. [Google Scholar] [CrossRef]
- Borrero, L.A. The human colonization of the high Andes and Southern South America during the cold pulses of the late Pleistocene. In Hunter-Gatherer Behavior; Routledge: Abingdon, UK, 2016; pp. 57–78. [Google Scholar]
- Le Neün, M.; Dufour, E.; Zazzo, A.; Tombret, O.; Thil, F.; Wheeler, J.C.; Cucchi, T.; Goepfert, N. Holocene occupation of the Andean highlands: A new radiocarbon chronology for the Telarmachay rockshelter (Central Andes, Peru). Quat. Sci. Rev. 2023, 312, 108146. [Google Scholar] [CrossRef]
- Lynch, T.F. Quaternary climate, environment, and the human occupation of the south-central Andes. Geoarchaeology 1990, 5, 199–228. [Google Scholar] [CrossRef]
- Bush, M.B.; Rozas-Davila, A.; Raczka, M.; Nascimento, M.; Valencia, B.; Sales, R.K.; McMichael, C.N.H.; Gosling, W.D. A paleoecological perspective on the transformation of the tropical Andes by early human activity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2022, 377, 20200497. [Google Scholar] [CrossRef]
- Schiferl, J.; Kingston, M.; Åkesson, C.; Valencia, B.; Rozas-Davila, A.; McGee, D.; Woods, A.; Chen, C.; Hatfield, R.; Rodbell, D. A neotropical perspective on the uniqueness of the Holocene among interglacials. Nat. Commun. 2023, 14, 7404. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento, F.O. Breaking mountain paradigms: Ecological effects on human impacts in managed tropandean landscapes. AMBIO J. Hum. Environ. 2000, 29, 423–431. [Google Scholar] [CrossRef]
- Erickson, C.L. The Lake Titicaca basin: A precolumbian built landscape. In Imperfect Balance; Lentz, D.E., Ed.; Columbia University Press: New York, NY, USA, 2000; pp. 311–356. [Google Scholar]
- Rademaker, K.; Hodgins, G.; Moore, K.; Zarrillo, S.; Miller, C.; Bromley, G.R.; Leach, P.; Reid, D.A.; Álvarez, W.Y.; Sandweiss, D.H. Paleoindian settlement of the high-altitude Peruvian Andes. Science 2014, 346, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Garvey, R.; Poe, K.; Pratt, L. Spatial and temporal trends in Peru’s radiocarbon record of middle Holocene foragers. Quat. Int. 2023. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Goodbred, S.; Pino, M.; Sánchez, V.F.V.; Tham, T.R.; Adovasio, J.; Collins, M.B.; Netherly, P.J.; Hastorf, C.A.; Chiou, K.L.; et al. Simple technologies and diverse food strategies of the Late Pleistocene and Early Holocene at Huaca Prieta, Coastal Peru. Sci. Adv. 2017, 3, e1602778. [Google Scholar] [CrossRef]
- Piperno, D.R. The origins of plant cultivation and domestication in the New World tropics: Patterns, process, and new developments. Curr. Anthropol. 2011, 52, S453–S470. [Google Scholar] [CrossRef]
- Pearsall, D.M. Plant Domestication and the Shift to Agriculture in the Andes. In The Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 105–120. [Google Scholar]
- Yacobaccio, H.D. The domestication of South American camelids: A review. Anim. Front. 2021, 11, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kuznar, L.A. Mutualism between Chenopodium, Herd Animals, and Herders in the South Central Andes. Mt. Res. Dev. 1993, 13, 257–265. [Google Scholar] [CrossRef]
- Kuentz, A.; Ledru, M.-P.; Thouret, J.-C. Environmental changes in the highlands of the western Andean Cordillera, southern Peru, during the Holocene. Holocene 2012, 22, 1215–1226. [Google Scholar] [CrossRef]
- Baker, P.A.; Fritz, S.C. Nature and causes of Quaternary climate variation of tropical South America. Quat. Sci. Rev. 2015, 124, 31–47. [Google Scholar] [CrossRef]
- Riris, P.; Arroyo-Kalin, M. Widespread population decline in South America correlates with mid-Holocene climate change. Sci. Rep. 2019, 9, 6850. [Google Scholar] [CrossRef] [PubMed]
- Vining, B.R.; Steinman, B.A.; Abbott, M.B.; Woods, A. Paleoclimatic and archaeological evidence from Lake Suches for highland Andean refugia during the arid middle-Holocene. Holocene 2019, 29, 328–344. [Google Scholar] [CrossRef]
- Knapp, G. Andean Ecology: Adaptive Dynamics in Ecuador; Routledge: Abingdon, UK, 2019. [Google Scholar]
- Brooks, S.O. Prehistoric Agricultural Terraces in the Rio Japo Basin, Colca Valley, Peru. Ph.D. Dissertation, The University of Wisconsin-Madison, Madison, WI, USA, 1998; p. 1020. [Google Scholar]
- Denevan, W.M. Cultivated Landscapes of Native Amazonia and the Andes; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Erickson, C.L. Neo-environmental determinism and agrarian ‘collapse’ in Andean prehistory. Antiquity 1999, 73, 634. [Google Scholar] [CrossRef]
- Binford, M.W.; Kolata, A.L.; Brenner, M.; Janusek, J.; Seddon, M.T.; Abbott, M.B.; Curtis, J.H. Climate variation and the rise and fall of an Andean civilization. Quat. Res. 1997, 47, 235–248. [Google Scholar] [CrossRef]
- Sandweiss, D.H.; Solís, R.S.; Moseley, M.E.; Keefer, D.K.; Ortloff, C.R. Environmental change and economic development in coastal Peru between 5800 and 3600 years ago. Proc. Natl. Acad. Sci. USA 2009, 106, 1359–1363. [Google Scholar] [CrossRef] [PubMed]
- Meyfroidt, P.; Chowdhury, R.R.; de Bremond, A.; Ellis, E.C.; Erb, K.-H.; Filatova, T.; Garrett, R.; Grove, J.M.; Heinimann, A.; Kuemmerle, T. Middle-range theories of land system change. Glob. Environ. Chang. 2018, 53, 52–67. [Google Scholar] [CrossRef]
- Contreras, D.A. Correlation is not enough: Building better arguments in the archaeology of human-environment interactions. In The Archaeology of Human-Environment Interactions; Contreras, D.A., Ed.; Routledge: Abingdon, UK, 2016; pp. 17–36. [Google Scholar]
- Bruno, M.C.; Capriles, J.M.; Hastorf, C.A.; Fritz, S.C.; Weide, D.M.; Domic, A.I.; Baker, P.A. The rise and fall of Wiñaymarka: Rethinking cultural and environmental interactions in the Southern Basin of Lake Titicaca. Hum. Ecol. 2021, 49, 131–145. [Google Scholar] [CrossRef]
- Luzar, J. The political ecology of a “forest transition”: Eucalyptus forestry in the Southern Peruvian Andes. Ethnobot. Res. Appl. 2007, 5, 85–93. [Google Scholar] [CrossRef]
- Colinvaux, P.; de Oliveira, P.E.; Moreno, P.J.E. Amazon Pollen Manual and Atlas; Harwood Academic Publishers: Amsterdam, The Netherlands, 1999; p. 332. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 5 May 2023).
- Reimer, P.J.; Austin, W.E.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.; Reimer, R. CALIB 8.2 [WWW program]. 2021. Available online: http://calib.org (accessed on 8 August 2023).
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; Wiley: Chichester, UK, 1989. [Google Scholar]
- Markgraf, V.; D’Antoni, H.L. Pollen Flora of Argentina; The University of Arizona Press: Tucson, AZ, USA, 1978; p. 208. [Google Scholar]
- Hooghiemstra, H. Vegetational and Climatic History of the High Plain of Bogota, Colombia. In Dissertaciones Botanicae 79; J. Cramer: Vaduz, Liechtenstein, 1984; p. 368. [Google Scholar]
- Bush, M.B.; Weng, C. Introducing a new (freeware) tool for palynology. J. Biogeogr. 2007, 34, 377–380. [Google Scholar] [CrossRef]
- Whitney, B.S.; Rushton, E.A.; Carson, J.F.; Iriarte, J.; Mayle, F.E. An improved methodology for the recovery of Zea mays and other large crop pollen, with implications for environmental archaeology in the Neotropics. Holocene 2012, 22, 1087–1096. [Google Scholar] [CrossRef]
- Grimm, E.C. CONISS: A fortran 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Rasband, W.S.; Rasband, W.S. ImageJ, U. S. National Institutes of Health, Bethesda, MD, USA. 1997–2016. Available online: https://imagej.nih.gov/ij/ (accessed on 5 May 2023).
- Richter, T.O.; Van der Gaast, S.; Koster, B.; Vaars, A.; Gieles, R.; de Stigter, H.C.; De Haas, H.; van Weering, T.C. The Avaatech XRF Core Scanner: Technical description and applications to NE Atlantic sediments. Geol. Soc. Lond. Spec. Publ. 2006, 267, 39–50. [Google Scholar] [CrossRef]
- Killick, R.; Eckley, I. changepoint: An R package for changepoint analysis. J. Stat. Softw. 2014, 58, 1–19. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J.A. rbacon: Age-Depth Modelling Using Bayesian Statistics, R Package Version 2.4 (2018); Queen’s University Belfast: Belfast, UK, 2018. [Google Scholar]
- Paduano, G.M.; Bush, M.B.; Baker, P.A.; Fritz, S.C.; Seltzer, G.O. A vegetation and fire history of Lake Titicaca since the Last Glacial Maximum. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 194, 259–279. [Google Scholar] [CrossRef]
- Valencia, B.G.; Urrego, D.H.; Silman, M.R.; Bush, M.B. From ice age to modern: A record of landscape change in an Andean cloud forest. J. Biogeogr. 2010, 37, 1637–1647. [Google Scholar] [CrossRef]
- Bird, B.W.; Abbott, M.B.; Rodbell, D.T.; Vuille, M. Holocene tropical South American hydroclimate revealed from a decadally resolved lake sediment δ18O record. Earth Planet. Sci. Lett. 2011, 310, 192–202. [Google Scholar] [CrossRef]
- Kanner, L.C.; Burns, S.J.; Cheng, H.; Edwards, R.L.; Vuille, M. High-resolution variability of the South American summer monsoon over the last seven millennia: Insights from a speleothem record from the central Peruvian Andes. Quat. Sci. Rev. 2013, 75, 1–10. [Google Scholar] [CrossRef]
- Haberzettl, T.; Corbella, H.; Fey, M.; Janssen, S.; Lücke, A.; Mayr, C.; Ohlendorf, C.; Schäbitz, F.; Schleser, G.H.; Wille, M. Lateglacial and Holocene wet—Dry cycles in southern Patagonia: Chronology, sedimentology and geochemistry of a lacustrine record from Laguna Potrok Aike, Argentina. Holocene 2007, 17, 297–310. [Google Scholar] [CrossRef]
- White, S. Grass páramo as hunter-gatherer landscape. Holocene 2013, 23, 898–915. [Google Scholar] [CrossRef]
- Perry, L.; Sandweiss, D.H.; Piperno, D.R.; Rademaker, K.; Malpass, M.A.; Umire, A.N.; de la Vera, P. Early maize agriculture and interzonal interaction in southern Peru. Nature 2006, 440, 76. [Google Scholar] [CrossRef] [PubMed]
- Washburn, E.; Nesbitt, J.; Burger, R.; Tomasto-Cagigao, E.; Oelze, V.M.; Fehren-Schmitz, L. Maize and dietary change in early Peruvian civilization: Isotopic evidence from the Late Preceramic Period/Initial Period site of La Galgada, Peru. J. Archaeol. Sci. Rep. 2020, 31, 102309. [Google Scholar] [CrossRef]
- Coetzee, J.A. Pollen analytical studies in east and southern Africa. Palaeoecol. Afr. 1967, 3, 1–146. [Google Scholar]
- Chepstow-Lusty, A.J.; Bennett, K.D.; Switsur, V.R.; Kendall, A. 4000 years of human impact and vegetation change in the central Peruvian Andes—With events parallelling the Maya record? Antiquity 1996, 70, 824–833. [Google Scholar] [CrossRef]
- Mosblech, N.A.S.; Chepstow-Lusty, A.; Valencia, B.G.; Bush, M.B. Anthropogenic control of late-Holocene landscapes in the Cuzco region, Peru. Holocene 2012, 22, 1361–1372. [Google Scholar] [CrossRef]
- Bush, M.B. Neotropical plant reproductive strategies and fossil pollen representation. Am. Nat. 1995, 145, 594–609. [Google Scholar] [CrossRef]
- Schittek, K.; Forbriger, M.; Mächtle, B.; Schäbitz, F.; Wennrich, V.; Reindel, M.; Eitel, B. Holocene environmental changes in the highlands of the southern Peruvian Andes (14 S) and their impact on pre-Columbian cultures. Clim. Past 2015, 11, 27–44. [Google Scholar] [CrossRef]
- Paduano, G. Vegetation and fire history of two tropical Andean lakes, Titicaca (Peru/Bolivia), and Caserochocha (Peru) with special emphasis on the Younger Dryas chronozone. In Department of Biological Sciences; Florida Institute of Technology: Melbourne, FL, USA, 2001. [Google Scholar]
- Villota, A.; Leon-Yanez, S.; Behling, H. Vegetation and environmental dynamics in the Páramo of Jimbura region in the southeastern Ecuadorian Andes during the late Quaternary. J. S. Am. Earth Sci. 2012, 40, 85–93. [Google Scholar] [CrossRef]
- Williams, J.J.; Gosling, W.D.; Coe, A.L.; Brooks, S.J.; Gulliver, P. Four thousand years of environmental change and human activity in the Cochabamba Basin, Bolivia. Quat. Res. 2011, 76, 58–68. [Google Scholar] [CrossRef]
- Bush, M.B.; Hansen, B.C.S.; Rodbell, D.; Seltzer, G.O.; Young, K.R.; Leo, B.; Silman, M.R.; Abbott, M.B.; Gosling, W.D. A 17,000 year history of Andean climatic and vegetation change from Laguna de Chochos, Peru. J. Quat. Sci. 2005, 20, 703–714. [Google Scholar] [CrossRef]
- Davis, O.K.; Shafer, D.S. Sporormiella fungal spores, a palynological means of detecting herbivore density. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 237, 40–50. [Google Scholar] [CrossRef]
- Gill, J.L. Bison Grazing Intensity Predicts the Abundance of the Dung Fungus Proxy Sporormiella at Konza Prairie: Implications for the Holocene Paleoecology of the Great Plains; Ecological Society of America: Portland, OR, USA, 2012. [Google Scholar]
- Ochoa, C. Maca (Lepidium meyenii Walp.; Brassicaceae): A nutritious root crop of the central Andes. Econ. Bot. 2001, 55, 344. [Google Scholar] [CrossRef]
- Hermann, M.; Bernet, T. The Transition of Maca from Neglect to Market Prominence; Agricultural Biodiversity and Livelihoods, Discussion Papers 1; Bioversity International: Rome, Italy, 2009. [Google Scholar]
- Pearsall, D.M. Adaptation of prehistoric hunter-gatherers to the high Andes: The changing role of plant resources. In Foraging and Farming; Routledge: Abingdon, UK, 2014; pp. 318–332. [Google Scholar]
- Bush, M.B.; Correa-Metrio, A.; McMichael, C.H.; Sully, S.; Shadik, C.R.; Valencia, B.G.; Guilderson, T.; Steinitz-Kannan, M.; Overpeck, J.T. A 6900-year history of landscape modification by humans in lowland Amazonia. Quat. Sci. Rev. 2016, 141, 52–64. [Google Scholar] [CrossRef]
- Rowe, H.D.; Guilderson, T.P.; Dunbar, R.B.; Southon, J.R.; Seltzer, G.O.; Mucciarone, D.A.; Fritz, S.C.; Baker, P.A. Late Quaternary lake-level changes constrained by radiocarbon and stable isotope studies on sediment cores from Lake Titicaca, South America. Glob. Planet. Chang. 2003, 38, 273–290. [Google Scholar] [CrossRef]
- Edwards, K.J.; Whittington, G. Lake sediments, erosion and landscape change during the Holocene in Britain and Ireland. Catena 2001, 42, 143–173. [Google Scholar] [CrossRef]
- Kiage, L.M.; Liu, K.-b. Late Quaternary paleoenvironmental changes in East Africa: A review of multiproxy evidence from palynology, lake sediments, and associated records. Prog. Phys. Geogr. 2006, 30, 633–658. [Google Scholar] [CrossRef]
- Bush, M.B.; Listopad, M.C.S.; Silman, M.R. A regional study of Holocene climate change and human occupation in Peruvian Amazonia. J. Biogeogr. 2007, 34, 1342–1356. [Google Scholar] [CrossRef]
- Moody, J.A.; Martin, D.A.; Cannon, S.H. Post-wildfire erosion response in two geologic terrains in the western USA. Geomorphology 2008, 95, 103–118. [Google Scholar] [CrossRef]
- Dorren, L.; Rey, F. A review of the effect of terracing on erosion. In Briefing Papers of the 2nd SCAPE Workshop; Boix-Fayons, C., Imeson, A., Eds.; 2004; pp. 97–108. Available online: https://www.ecorisq.org/docs/Dorren_Rey.pdf (accessed on 18 March 2024).
- Molina, A.; Govers, G.; Vanacker, V.; Poesen, J.; Zeelmaekers, E.; Cisneros, F. Runoff generation in a degraded Andean ecosystem: Interaction of vegetation cover and land use. Catena 2007, 71, 357–370. [Google Scholar] [CrossRef]
- Xu, H.; Liu, B.; Wu, F. Spatial and temporal variations of Rb/Sr ratios of the bulk surface sediments in Lake Qinghai. Geochem. Trans. 2010, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Cao, J.; Wu, J.; Wang, S. A Rb/Sr record of catchment weathering response to Holocene climate change in Inner Mongolia. Earth Surf. Process. Landf. J. Br. Geomorphol. Res. Group 2006, 31, 285–291. [Google Scholar] [CrossRef]
- Isbell, W.H. Wari and Tiwanaku: International Identities in the Central Andean Middle Horizon. In The Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 731–759. [Google Scholar]
- Williams, P.R. Agricultural innovation, intensification, and sociopolitical development: The case of highland irrigation agriculture on the Pacific Andean watersheds. In Agricultural Strategies; Marcus, J., Stanish, C., Eds.; Cotsen Institute of Archaeology, Univ of California: Los Angeles, CA, USA, 2006; pp. 309–333. [Google Scholar]
- Covey, A.R. The Inca Empire. In Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 809–830. [Google Scholar]
- Hemming, J. The Conquest of the Incas; Harcourt: New York, NY, USA, 1970. [Google Scholar]
- Cook, N.D. Born to Die: Disease and New World Conquest, 1492–1650; Cambridge University Press: Cambridge, UK, 1998; Volume 1. [Google Scholar]
- Langlie, B.S. Building ecological resistance: Late intermediate period farming in the south-central highland Andes (CE 1100–1450). J. Anthropol. Archaeol. 2018, 52, 167–179. [Google Scholar] [CrossRef]
- Kemp, R.; Branch, N.; Silva, B.; Meddens, F.; Williams, A.; Kendall, A.; Vivanco, C. Pedosedimentary, cultural and environmental significance of paleosols within pre-hispanic agricultural terraces in the southern Peruvian Andes. Quat. Int. 2006, 158, 13–22. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab I | Depth (cm) | 14C (yr BP) ± 1 σ | cal BP 2σ Range | Median Age Cal BP | 13C | ||||
|---|---|---|---|---|---|---|---|---|---|
| OS-135406 * | 7 | 5070 | ± | 40 | 5660 | – | 5897 | 5817 | −27.51 |
| OS-125490 * | 71 | 4500 | ± | 20 | 5159 | – | 5287 | 5167 | −25.5 |
| DAMS-052062 * | 79 | 3814 | ± | 34 | 4089 | – | 4396 | 4203 | |
| OS-126479 * | 81 | 2610 | ± | 20 | 2698 | – | 2757 | 2749 | −24.35 |
| OS-135405 * | 120 | 3140 | ± | 25 | 3222 | – | 3383 | 3368 | −27.23 |
| OS-125489 | 156 | 2480 | ± | 20 | 2633 | – | 2701 | 2589 | −25.4 |
| OS-125503 | 365.5 | 3070 | ± | 20 | 3283 | – | 3345 | 3291 | −25.84 |
| OS-125504 | 578 | 4180 | ± | 20 | 4570 | – | 4823 | 4728 | −25.91 |
| OS-125505 | 698 | 5610 | ± | 25 | 6298 | – | 6407 | 6372 | −26.69 |
| Max % of Brassicaceae% | Peak Time Cal BP | Max % Solanaceae% | Peak Time Cal BP | Site Elevation (masl) | Source | |
|---|---|---|---|---|---|---|
| Cerro Llamoca | 11 | 600 | 2 | 400 | 4450 | [65] |
| Caserococha | 3 | 3000 | 3 | 2000 | 3900 | [66] |
| Titicaca | 3 | 4000 | 0.6 | 4720 | 3810 | [53] |
| Acopia | 11.3 | 424 | 3.5 | 4280 | 3750 | this ms |
| Huaypo | 3.7 | 340 | 0.4 | 3500 | [63] | |
| Natosa Peat bog | 5 | 4000 | <2 | 3482 | [67] | |
| Challacaba | 7 | 300 | <2 | 3400 | [68] | |
| Marcacocha | 30 | −20 | <2 | 3350 | [62] | |
| Chochos | 1 | 1.9 | 3285 | [69] | ||
| Pacucha | 6 | 270 | 1.5 | 3100 | [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shadik, C.R.; Bush, M.B.; Valencia, B.G.; Rozas-Davila, A.; Plekhov, D.; Breininger, R.D.; Davin, C.; Benko, L.; Peterson, L.C.; VanValkenburgh, P. The Evolution of Agrarian Landscapes in the Tropical Andes. Plants 2024, 13, 1019. https://doi.org/10.3390/plants13071019
Shadik CR, Bush MB, Valencia BG, Rozas-Davila A, Plekhov D, Breininger RD, Davin C, Benko L, Peterson LC, VanValkenburgh P. The Evolution of Agrarian Landscapes in the Tropical Andes. Plants. 2024; 13(7):1019. https://doi.org/10.3390/plants13071019
Chicago/Turabian StyleShadik, Courtney R., Mark B. Bush, Bryan G. Valencia, Angela Rozas-Davila, Daniel Plekhov, Robert D. Breininger, Claire Davin, Lindsay Benko, Larry C. Peterson, and Parker VanValkenburgh. 2024. "The Evolution of Agrarian Landscapes in the Tropical Andes" Plants 13, no. 7: 1019. https://doi.org/10.3390/plants13071019
APA StyleShadik, C. R., Bush, M. B., Valencia, B. G., Rozas-Davila, A., Plekhov, D., Breininger, R. D., Davin, C., Benko, L., Peterson, L. C., & VanValkenburgh, P. (2024). The Evolution of Agrarian Landscapes in the Tropical Andes. Plants, 13(7), 1019. https://doi.org/10.3390/plants13071019