Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean


 ,
, 


Abstract
:1. Introduction
2. Results
2.1. Biostimulant Application Positively Influenced Seed Morphological Parameters and Germination Percentage under Controlled Conditions
2.2. Biostimulant Application Modulated the Expression of 879 Genes under Controlled Conditions
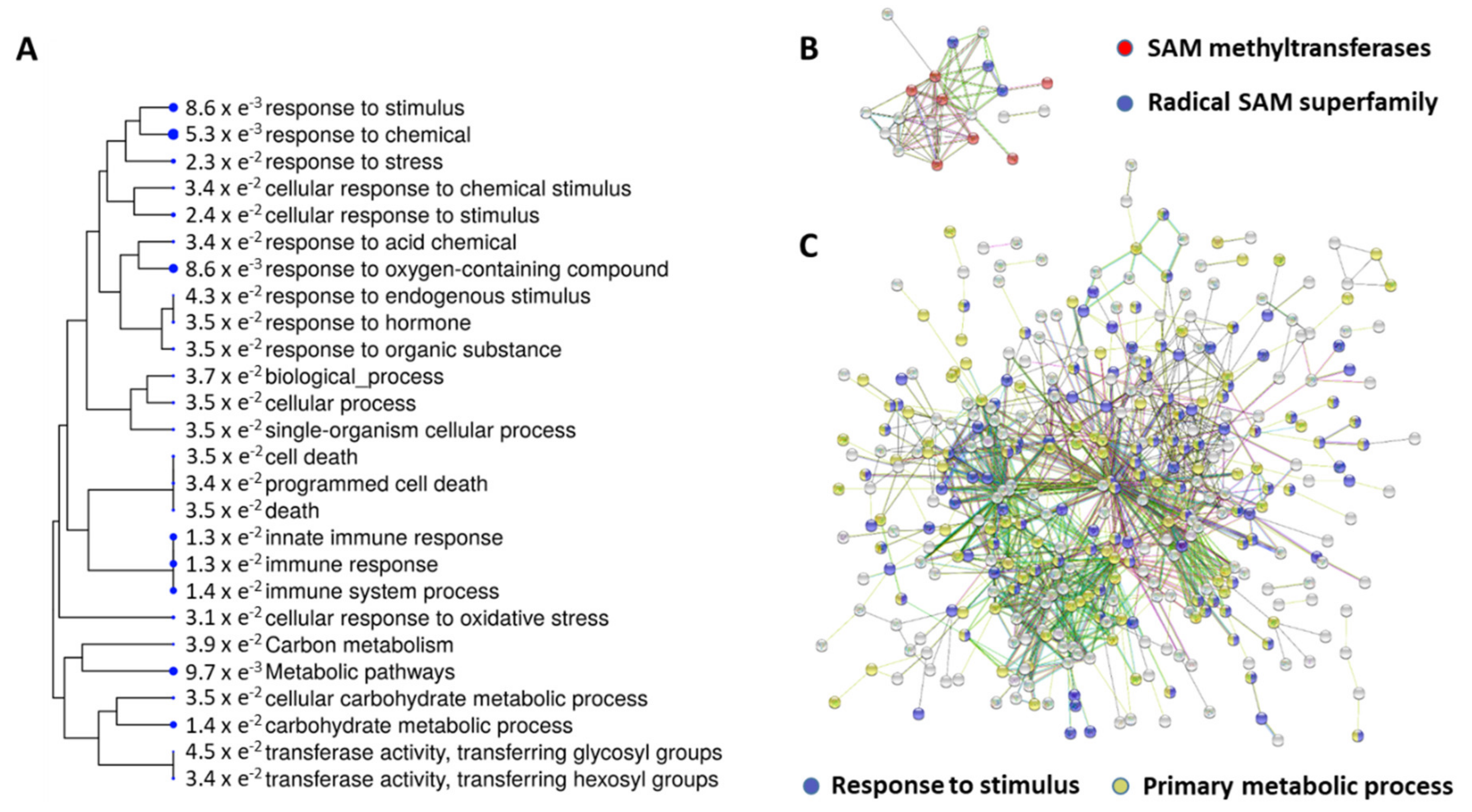
2.3. String Suite Analysis Revealed Two Functional Interaction Networks
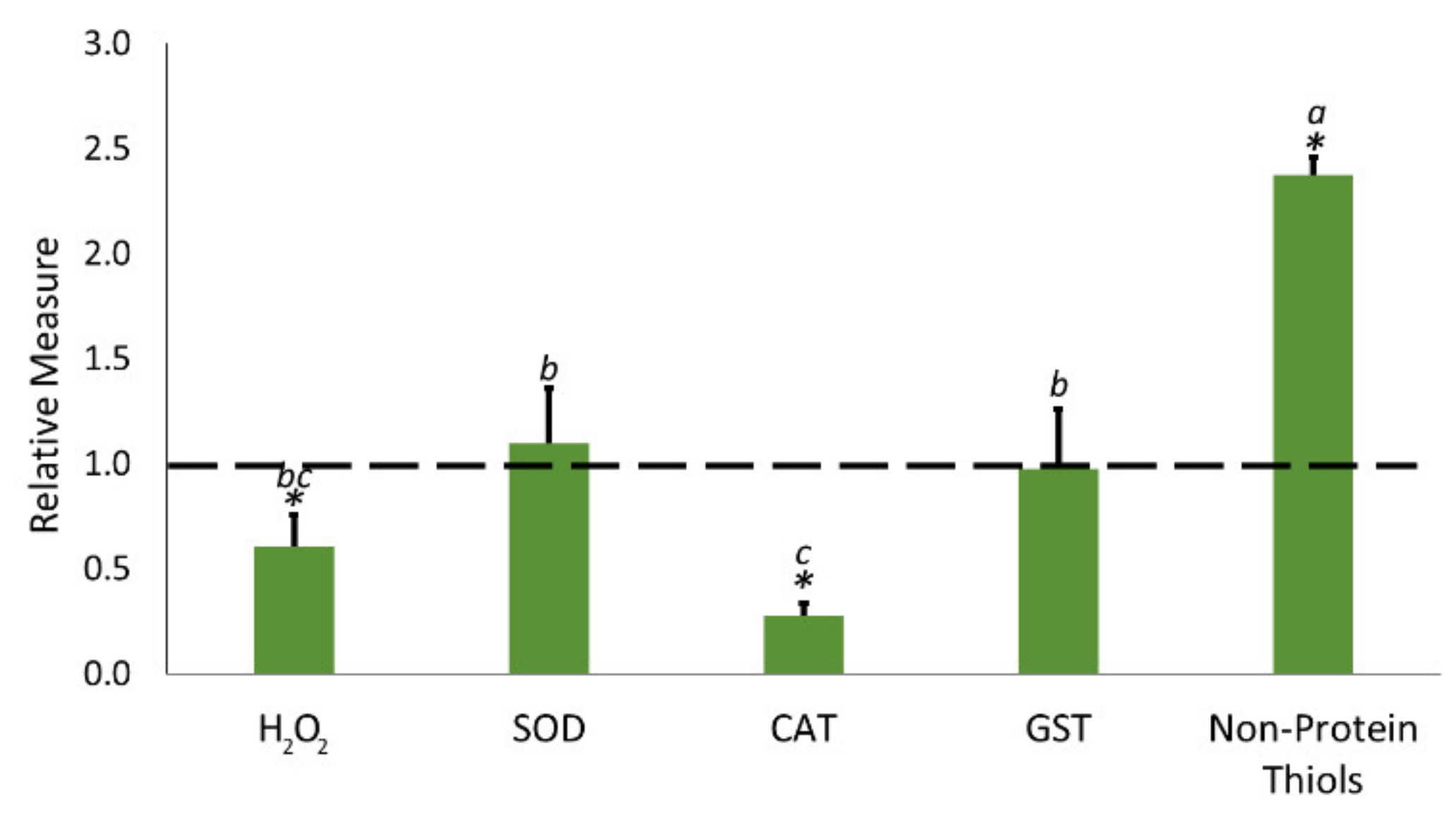
2.4. Biostimulant Application Reduced Oxidative Menaces
3. Discussion
3.1. The Biostimulant Promoted Soybean Germination and Growth after Heat Stress Exposure
3.2. Rna-Seq Analysis Displayed That Heat Stress Mitigation after Biostimulant-Treatment May Be Linked to Different Molecular Pathways
3.3. Biostimulant Treatment Mitigated the Accumulation of Reactive Oxygen Species (Ros) via a Non-Enzymatic Way
3.4. Soybean Seed Primary Metabolism and Hormone Signaling Are Affected by the Biostimulant Treatment
4. Materials and Methods
4.1. Plant Material and Biostimulant
4.2. Experiments in Controlled Conditions
4.3. Evaluation of Biometric Parameters under Controlled Conditions
4.4. Rna Isolation and Rna-Seq Analysis of Seeds Incubated under Controlled Conditions
4.4.1. Total Rna Isolation
4.4.2. RNA-Seq Analysis
4.5. Evaluation of Antioxidant Enzyme Activity of Seeds Incubated under Controlled Conditions
4.5.1. Superoxide Dismutase Activity (Sod, EC 1.15.1.1).
4.5.2. Catalase Activity (CAT, EC 1.11.1.6).
4.5.3. Glutathione-S-transferases (GST, EC 2.5.1.18)
4.6. Evaluation of Non-Protein Thiol Content of Seeds Incubated under Controlled Conditions
4.7. Evaluation of Hydrogen Peroxide Levels of Seeds Incubated under Controlled Conditions
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pagano, M.C.; Miransari, M. Production Worldwide; Elsevier Inc.: Amsterdam, The Netherlands, 2016; ISBN 9780128015360. [Google Scholar]
- Wingeyer, A.B.; Amado, T.J.C.; Pérez-Bidegain, M.; Studdert, G.A.; Perdomo Varela, C.H.; Garcia, F.O.; Karlen, D.L. Soil quality impacts of current South American agricultural practices. Sustainability 2015, 7, 2213–2242. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Chebrolu, K.K.; Fritschi, F.B.; Ye, S.; Krishnan, H.B.; Smith, J.R.; Gillman, J.D. Impact of heat stress during seed development on soybean seed metabolome. Metabolomics 2016, 12, 1–14. [Google Scholar] [CrossRef]
- Teixeira, E.I.; Fischer, G.; Van Velthuizen, H.; Walter, C.; Ewert, F. Global hot-spots of heat stress on agricultural crops due to climate change. Agric. For. Meteorol. 2013, 170, 206–215. [Google Scholar] [CrossRef]
- Probert, R.J. The role of temperature in the regulation of seed dormancy and germination. Seeds Ecol. Regen. Plant Communities 2000, 2, 261–292. [Google Scholar]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Pre-Sowing Seed Treatment-A Shotgun Approach to Improve Germination, Plant Growth, and Crop Yield Under Saline and Non-Saline Conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar]
- Campobenedetto, C.; Grange, E.; Mannino, G.; Van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A Biostimulant Seed Treatment Improved Heat Stress Tolerance During Cucumber Seed Germination by Acting on the Antioxidant System and Glyoxylate Cycle. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Ziosi, V.; Zandoli, R.; Vitali, F.; Di Nardo, A. Folicist®, a biostimulant based on acetyl-thioproline, folic acid and plant extracts, improves seed germination and radicle extension. Acta Hortic. 2012, 1009, 79–82. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Masondo, N.A.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Influence of biostimulants-seed-priming on Ceratotheca triloba germination and seedling growth under low temperatures, low osmotic potential and salinity stress. Ecotoxicol. Environ. Saf. 2018, 147, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Kavipriya, R.; Dhanalakshmi, P.K.; Jayashree, S.; Thangaraju, N. Seaweed extract as a biostimulant for legume crop, green gram. J. Ecobiotechnol. 2011, 3, 16–19. [Google Scholar]
- Qiu, Y.; Amirkhani, M.; Mayton, H.; Chen, Z.; Taylor, A.G. Biostimulant seed coating treatments to improve cover crop germination and seedling growth. Agronomy 2020, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Mannino, G.; Nerva, L.; Gritli, T.; Novero, M.; Fiorilli, V.; Bacem, M.; Bertea, C.M.; Lumini, E.; Chitarra, W.; Balestrini, R. Effects of Different Microbial Inocula on Tomato Tolerance to Water Deficit. Agronomy 2020, 10, 170. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; Fernandes, Â.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Barreira, J.C.M.; Barros, L.; Ferreira, I.C.F.R. Biostimulants application alleviates water stress effects on yield and chemical composition of greenhouse green bean (Phaseolus vulgaris L.). Agronomy 2020, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 871. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Lucini, L.; Miras-Moreno, B.; Colla, G.; Bonini, P.; Cardarelli, M. Metabolomic Responses of Maize Shoots and Roots Elicited by Combinatorial Seed Treatments With Microbial and Non-microbial Biostimulants. Front. Microbiol. 2020, 11, 664. [Google Scholar] [CrossRef]
- De Pádua, G.P.; França-Neto, J.D.B.; De Carvalho, M.L.M.; Krzyzanowski, F.C.; Guimarães, R.M. Incidence of green soybean seeds as a function of environmental stresses during seed maturation. Rev. Bras. Sementes 2009, 31, 150–159. [Google Scholar] [CrossRef]
- Hulme, M.; Sheard, N. Climate Change Scenarios for Brazil; Climate Research Unit: Norwich, UK, 1999. [Google Scholar]
- Ferreira, D.B.; Rao, V.B. Recent climate variability and its impacts on soybean yields in Southern Brazil. Theor. Appl. Climatol. 2011, 105, 83–97. [Google Scholar] [CrossRef]
- Struck, A.W.; Thompson, M.L.; Wong, L.S.; Micklefield, J. S-Adenosyl-Methionine-Dependent Methyltransferases: Highly Versatile Enzymes in Biocatalysis, Biosynthesis and Other Biotechnological Applications. ChemBioChem 2012, 13, 2642–2655. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Frey, P.A. S-adenosylmethionine as an oxidant: The radical SAM superfamily. Trends Biochem. Sci. 2007, 32, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Wang, Y.; Li, Q.; Wang, D.; Xing, C.; Zhang, L.; Xu, T.; Fang, F.; Wang, F. Accumulating mechanism of γ-aminobutyric acid in soybean (Glycine max L.) during germination. Int. J. Food Sci. Technol. 2018, 53, 106–111. [Google Scholar] [CrossRef]
- Taylor, A.G.; Allen, P.S.; Bennett, M.A.; Bradford, K.J.; Burris, J.S.; Misra, M.K. Seed enhancements. Seed Sci. Res. 1998, 8, 245–256. [Google Scholar] [CrossRef]
- Reddy, P.P. Climate Resilient Agriculture for Ensuring Food Security; Springer: Berlin, Germany, 2015; Volume 373. [Google Scholar]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.J.P.; Buitink, J.; Lalanne, D.; Rossi, R.F.; Pelletier, S.; Da Silva, E.A.A.; Leprince, O. Molecular characterization of the acquisition of longevity during seed maturation in soybean. PLoS ONE 2017, 12, e0180282. [Google Scholar]
- Ali, Q.; Perveen, R.; El-Esawi, M.A.; Ali, S.; Hussain, S.M.; Amber, M.; Iqbal, N.; Rizwan, M.; Alyemeni, M.N.; El-Serehy, H.A.; et al. Low Doses of Cuscuta reflexa Extract Act as Natural Biostimulants to Improve the Germination Vigor, Growth, and Grain Yield of Wheat Grown under Water Stress: Photosynthetic Pigments, Antioxidative Defense Mechanisms, and Nutrient Acquisition. Biomolecules 2020, 10, 1212. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities inwheat seedlings. Plant Physiol. Biochem. 2014, 74, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Savy, D.; Cozzolino, V.; Nebbioso, A.; Drosos, M.; Nuzzo, A.; Mazzei, P.; Piccolo, A. Humic-like bioactivity on emergence and early growth of maize (Zea mays L.) of water-soluble lignins isolated from biomass for energy. Plant Soil 2016, 402, 221–233. [Google Scholar] [CrossRef]
- Amirkhani, M.; Netravali, A.N.; Huang, W.; Taylor, A.G. Investigation of soy protein–based biostimulant seed coating for broccoli seedling and plant growth enhancement. HortScience 2016, 51, 1121–1126. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Deshmukh, R.; Muthamilarasan, M.; Rani, R.; Prasad, M. Versatile roles of aquaporin in physiological processes and stress tolerance in plants. Plant Physiol. Biochem. 2020, 149, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Fessehaie, A.; Arora, R. Aquaporin expression during seed osmopriming and post-priming germination in spinach. Biol. Plant 2013, 57, 193–198. [Google Scholar] [CrossRef]
- Noodén, L.D.; Blakley, K.A.; Grzybowski, J.M. Control of Seed Coat Thickness and Permeability in Soybean. Plant Physiol. 1985, 79, 543–545. [Google Scholar]
- Nair, P.; Kandasamy, S.; Zhang, J.; Ji, X.; Kirby, C.; Benkel, B.; Hodges, M.D.; Critchley, A.T.; Hiltz, D.; Prithiviraj, B. Transcriptional and metabolomic analysis of Ascophyllum nodosum mediated freezing tolerance in Arabidopsis thaliana. BMC Genom. 2012, 13, 643. [Google Scholar] [CrossRef] [Green Version]
- Contartese, V.; Garabello, C.; Occhipinti, A.; Barbero, F.; Bertea, C.M. Effects of a new biostimulant on gene expression and metabolic responses of tomato plants. In Proceedings of the II World Congress on the Use of Biostimulants in Agriculture 1148, Florence, Italy, 16–19 November 2015; pp. 35–42. [Google Scholar]
- Tirnaz, S.; Batley, J. DNA Methylation: Toward Crop Disease Resistance Improvement. Trends Plant Sci. 2019, 24, 1137–1150. [Google Scholar] [CrossRef]
- Conrath, U. Molecular aspects of defence priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef]
- Kim, J.M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 2015, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Manduzio, S.; Kang, H. Epitranscriptomic RNA methylation in plant development and abiotic stress responses. Front. Plant Sci. 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genuth, N.R.; Barna, M. The discovery of ribosome heterogeneity and its implications for gene regulation and organismal life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, A.; Das, P.; Chaudhuri, S.; Bevilacqua, E.; Andrews, J.; Barik, S.; Hatzoglou, M.; Komar, A.A.; Mazumder, B. Requirement of rRNA methylation for 80S ribosome assembly on a cohort of cellular internal ribosome entry sites. Mol. Cell. Biol. 2011, 31, 4482–4499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schosserer, M.; Minois, N.; Angerer, T.B.; Amring, M.; Dellago, H.; Harreither, E.; Calle-Perez, A.; Pircher, A.; Gerstl, M.P.; Pfeifenberger, S. Methylation of ribosomal RNA by NSUN5 is a conserved mechanism modulating organismal lifespan. Nat. Commun. 2015, 6, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Liberman, N.; O’Brown, Z.K.; Earl, A.S.; Boulias, K.; Gerashchenko, M.V.; Wang, S.Y.; Fritsche, C.; Fady, P.-E.; Dong, A.; Gladyshev, V.N. N6-adenosine methylation of ribosomal RNA affects lipid oxidation and stress resistance. Sci. Adv. 2020, 6, eaaz4370. [Google Scholar] [CrossRef] [Green Version]
- Lisso, J.; Altmann, T.; Müssig, C. The AtNFXL1 gene encodes a NF-X1 type zinc finger protein required for growth under salt stress. FEBS Lett. 2006, 580, 4851–4856. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Pandey, G.K. Expansion and function of repeat domain proteins during stress and development in plants. Front. Plant Sci. 2016, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Laluk, K.; Abuqamar, S.; Mengiste, T. The arabidopsis mitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Physiol. 2011, 156, 2053–2068. [Google Scholar] [CrossRef] [Green Version]
- Li, P.S.; Yu, T.F.; He, G.H.; Chen, M.; Zhou, Y.-B.; Chai, S.C.; Xu, Z.S.; Ma, Y.Z. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genom. 2014, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis HsfB1 and HsfB2b act as repressors of the expression of heat-inducible Hsfs but positively regulate the acquired thermotolerance. Plant Physiol. 2011, 157, 1243–1254. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Torres, M.A.; Dangl, J.L.; Jones, J.D.G. Arabidopsis gp91phox homologues Atrbohd and Atrbohf are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Wojtyla, Ł.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different modes of hydrogen peroxide action during seed germination. Front. Plant Sci. 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, H.; Galant, A.; Ravilious, G.E.; Preuss, M.L.; Jez, J.M. Sensing sulfur conditions: Simple to complex protein regulatory mechanisms in plant thiol metabolism. Mol. Plant 2010, 3, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Szalai, G.; Kellős, T.; Galiba, G.; Kocsy, G. Glutathione as an antioxidant and regulatory molecule in plants under abiotic stress conditions. J. Plant Growth Regul. 2009, 28, 66–80. [Google Scholar] [CrossRef]
- Kocsy, G.; Szalai, G.; Galiba, G. Induction of glutathione synthesis and glutathione reductase activity by abiotic stresses in maize and wheat. Sci. World J. 2002, 2, 1699–1705. [Google Scholar] [CrossRef] [Green Version]
- Peltzer, D.; Dreyer, E.; Polle, A. Differential temperature dependencies of antioxidative enzymes in two contrasting species: Fagus sylvatica and Coleus blumei. Plant Physiol. Biochem. 2002, 40, 141–150. [Google Scholar] [CrossRef]
- Nieto-Sotelo, J.; Ho, T.-H.D. Effect of heat shock on the metabolism of glutathione in maize roots. Plant Physiol. 1986, 82, 1031–1035. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ. Exp. Bot. 2015, 112, 44–54. [Google Scholar] [CrossRef]
- Jain, M.; Bhalla-Sarin, N. Glyphosate-induced increase in glutathione S-transferase activity and glutathione content in groundnut (Arachis hypogaea L.). Pestic. Biochem. Physiol. 2001, 69, 143–152. [Google Scholar] [CrossRef]
- Zagorchev, L.; Seal, C.E.; Kranner, I.; Odjakova, M. A central role for thiols in plant tolerance to abiotic stress. Int. J. Mol. Sci. 2013, 14, 7405–7432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul. 2014, 74, 139–152. [Google Scholar] [CrossRef]
- Ali, M.; Hayat, S.; Ahmad, H.; Ghani, M.I.; Amin, B.; Atif, M.J.; Cheng, Z. Priming of Solanum melongena L. seeds enhances germination, alters antioxidant enzymes, modulates ROS, and improves early seedling growth: Indicating aqueous garlic extract as seed-priming bio-stimulant for eggplant production. Appl. Sci. 2019, 9, 2203. [Google Scholar] [CrossRef] [Green Version]
- Bellieny-Rabelo, D.; De Oliveira, E.A.G.; Da Silva Ribeiro, E.; Pessoa Costa, E.; Oliveira, A.E.A.; Venancio, T.M. Transcriptome analysis uncovers key regulatory and metabolic aspects of soybean embryonic axes during germination. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, H.; Huber, S.C. Regulation of sucrose metabolism in higher plants: Localization and regulation of activity of key enzymes. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 253–289. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, O.; Béthencourt, L.; Quero, A.; Sangwan, R.S.; Clément Christophe, C. Trehalose and plant stress responses: Friend or foe? Trends Plant Sci. 2010, 15, 409–417. [Google Scholar] [CrossRef]
- De La Torre, F.; Cañas, R.A.; Pascual, M.B.; Avila, C.; Cánovas, F.M. Plastidic aspartate aminotransferases and the biosynthesis of essential amino acids in plants. J. Exp. Bot. 2014, 65, 5527–5534. [Google Scholar] [CrossRef] [Green Version]
- Balestrazzi, A.; Confalonieri, M.; Macovei, A.; Donà, M.; Carbonera, D. Genotoxic stress and DNA repair in plants: Emerging functions and tools for improving crop productivity. Plant Cell Rep. 2011, 30, 287–295. [Google Scholar] [CrossRef]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- Shuai, H.; Meng, Y.; Luo, X.; Chen, F.; Zhou, W.; Dai, Y.; Qi, Y.; Du, J.; Yang, F.; Liu, J.; et al. Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henning, A.A. Patologia e Tratamento de Sementes: Noções Gerais; Embrapa Soja: Brasília, Brazil, 2005. [Google Scholar]
- Dos Santos, S.F.; Carvalho, E.R.; Rocha, D.K.; Nascimento, R.M. Composition and volumes of slurry in soybean seeds treatment in the industry and physiological quality during storage. J. Seed Sci. 2018, 40, 67–74. [Google Scholar] [CrossRef]
- Tanabata, T.; Shibaya, T.; Hori, K.; Ebana, K.; Yano, M. SmartGrain: High-throughput phenotyping software for measuring seed shape through image analysis. Plant Physiol. 2012, 160, 1871–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannino, G.; Perrone, A.; Campobenedetto, C.; Schittone, A.; Margherita Bertea, C.; Gentile, C. Phytochemical profile and antioxidative properties of Plinia trunciflora fruits: A new source of nutraceuticals. Food Chem. 2020, 307, 125515. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, G.; Zhang, X.; Wang, F.; Song, R. Isolation of high quality RNA from cereal seeds containing high levels of starch. Phytochem. Anal. 2012, 23, 159–163. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Krishnan, N.; Chattopadhyay, S.; Kundu, J.K.; Chaudhuri, A. Superoxide dismutase activity in haemocytes and haemolymph of Bombyx mori following bacterial infection. Curr. Sci. 2002, 321–325. [Google Scholar]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Seed Parameters | Untreated | Biostimulant-Treated | %Δ |
|---|---|---|---|
| Area | 84.46 ± 4.91 a | 134.80 ± 8.90 b | 59.55% ± 1.26% |
| Perimeter | 40.71 ± 2.72 a | 58.99 ± 2.71 b | 45.04% ± 3.04% |
| Length | 55.62 ± 2.74 a | 76.01 ± 3.35 b | 36.68% ± 0.71% |
| Width | 13.08 ± 1.55 a | 27.82 ± 3.17 b | 112.77% ± 0.98% |
| Weight | 101.86 ± 2.33 a | 102.50 ± 3.49 a | 0.61% ± 0.02% |
| Untreated | Biostimulant-Treated | %Δ | |
|---|---|---|---|
| 24 h | n.d. | n.d. | - |
| 48 h | 68.80 ± 1.16 a | 77.70 ± 0.58 b | 12.95% ± 1.06% |
| 72 h | 82.22 ± 0.58 a | 91.13 ± 0.55 b | 10.84% ± 0.11% |
| Pathway ID GO Category | Pathway Description | Count in Gene Set | FDR |
|---|---|---|---|
| Upregulated Genes | |||
| IPR029063 | SAM-dependent methyltransferase | 9 | 6.7 × e−08 |
| IPR040072 | Methyltransferases (Class A) | 3 | 8.8 × e−05 |
| IPR004383 | Ribosomal RNA large subunit methyltransferase RlmN/Cfr | 3 | 5.73 × e−06 |
| IPR027492 | Dual-specificity RNA methyltransferase RlmN | 3 | 0.332 |
| IPR015507 | Ribosomal RNA large subunit methyltransferase E | 2 | 0.00509 |
| IPR002877 | Ribosomal RNA methyltransferase FtsJ domain | 2 | 0.00847 |
| IPR023267 | RNA (C5-cytosine) methyltransferase | 2 | 0.0263 |
| IPF01728 | FtsJ-like methyltransferase | 2 | 0.0149 |
| IPR006638 | Elongator protein 3/MiaB/NifB | 2 | 0.0106 |
| IPR001678 | SAM-dependent methyltransferase RsmB/NOP2-type | 2 | 0.0304 |
| Downregulated Genes | |||
| GO:0042221 | Response to chemical | 90 | 8.83 × e−11 |
| GO:0050896 | Response to stimulus | 135 | 2.24 × e−10 |
| GO:0044699 | Single-organism process | 186 | 9.73 × e−09 |
| GO:0008152 | Metabolic process | 211 | 1.21 × e−07 |
| GO:0044763 | Single-organism cellular process | 154 | 2.49 × e−07 |
| GO:0009987 | Cellular process | 213 | 3.13 × e−06 |
| GO:0006950 | Response to stress | 80 | 5.49 × e−06 |
| GO:0010033 | Response to organic substance | 60 | 1.13 × e−05 |
| GO:0071704 | Organic substance metabolic process | 175 | 2.30 × e−05 |
| GO:0009628 | Response to abiotic stimulus | 52 | 3.51 × e−05 |
| GO:0044238 | Primary metabolic process | 160 | 0.000933 |
| GO:0005975 | Carbohydrate metabolic process | 35 | 0.00117 |
| GO:0009755 | Hormone-mediated signaling pathway | 34 | 0.00117 |
| GO:0009058 | Biosynthetic process | 100 | 0.00211 |
| GO:0006952 | Defense response | 39 | 0.00228 |
| GO:1901698 | Response to nitrogen compound | 14 | 0.00244 |
| GO:0007165 | Signal transduction | 42 | 0.00993 |
| GO:0071554 | Cell wall organization or biogenesis | 21 | 0.00993 |
| Soybean Locus | Arabidopsis Locus | Category | Annotation |
|---|---|---|---|
| LOC100780381 | AT1G01860 | rRNA methylation | mRNA (2’-O-methyladenosine-N6-)-methyltransferase activity, rRNA (adenine-N6,N6-)-dimethyltransferase activity |
| LOC100796633 | AT1G54310 | rRNA methylation | RNA binding |
| LOC100813692 | AT1G60230 | rRNA methylation | rRNA base methylation, tRNA methylation |
| LOC100818638 | AT2G39670 | rRNA methylation | rRNA base methylation, tRNA methylation |
| LOC100779327 | AT3G13180 | rRNA methylation | rRNA (cytosine-C5-)-methyltransferase activity |
| LOC100804869 | AT3G19630 | rRNA methylation | rRNA base methylation, tRNA methylation |
| LOC100778294 | AT3G57000 | rRNA methylation | rRNA (pseudouridine) methyltransferase activity, rRNA binding |
| LOC100807160 | AT4G26600 | rRNA methylation | S-adenosylmethionine-dependent methyltransferase activity, rRNA (cytosine-C5-)-methyltransferase activity |
| LOC100809923 | AT5G01230 | rRNA/tRNA methylation | RNA methyltransferase activity, tRNA methyltransferase activity |
| LOC100787277 | AT5G10910 | rRNA methylation | Plastid rRNA methyltransferase involved in ribosome biogenesis and plant development. Accounts to the N4-methylation of C1352 in chloroplast 16S rRNA. |
| LOC100805818 | AT5G13830 | rRNA methylation | RNA methyltransferase activity, rRNA (uridine-2’-O-)-methyltransferase activity |
| LOC100818428 | AT5G50110 | rRNA methylation | rRNA (guanine-N7-)-methyltransferase activity, rRNA methyltransferase activity |
| LOC100804672 | AT1G12650 | Ribosome assembly | Cleavage involved in rRNA processing, maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) |
| LOC100798373 | AT5G20600 | Ribosome assembly | rRNA processing |
| LOC100802650 | AT5G38720 | Ribosome assembly | rRNA processing, ribosomal small subunit assembly |
| LOC100808860 | AT5G40530 | Ribosome assembly | Methylated histone binding involved in rDNA heterochromatin assembly |
| LOC100816856 | AT5G08420 | Ribosome assembly | Small subunit processome (nucleolus, nucleus) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campobenedetto, C.; Mannino, G.; Agliassa, C.; Acquadro, A.; Contartese, V.; Garabello, C.; Bertea, C.M. Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants 2020, 9, 1308. https://doi.org/10.3390/plants9101308
Campobenedetto C, Mannino G, Agliassa C, Acquadro A, Contartese V, Garabello C, Bertea CM. Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants. 2020; 9(10):1308. https://doi.org/10.3390/plants9101308
Chicago/Turabian StyleCampobenedetto, Cristina, Giuseppe Mannino, Chiara Agliassa, Alberto Acquadro, Valeria Contartese, Christian Garabello, and Cinzia Margherita Bertea. 2020. "Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean" Plants 9, no. 10: 1308. https://doi.org/10.3390/plants9101308
APA StyleCampobenedetto, C., Mannino, G., Agliassa, C., Acquadro, A., Contartese, V., Garabello, C., & Bertea, C. M. (2020). Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants, 9(10), 1308. https://doi.org/10.3390/plants9101308