
Effects of Extreme Temperature and Precipitation Events on Daily CO2 Fluxes in the Tropics

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
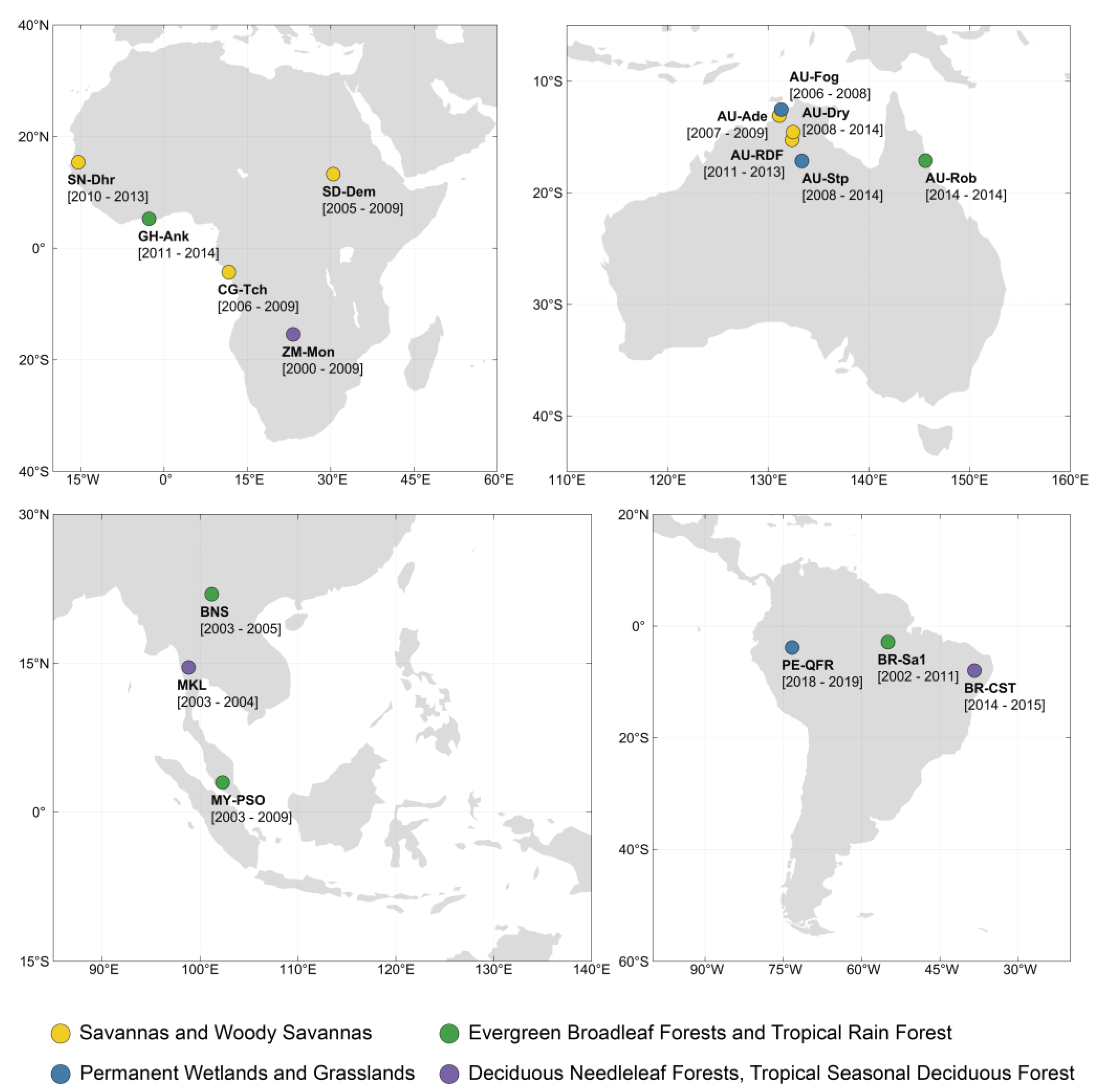
2.1. Meteorological and CO2 Flux Data Sets
2.2. Data Analysis
- −
- as a period when the daily mean temperature anomaly/daily precipitation amount exceeded 95% (90%) quantile (for extreme high temperature and precipitation) or did not reach 5% (10%) quantile (for extreme low temperature and precipitation) of probability density function (PDF);
- −
- as a period when the daily mean temperature anomaly exceeded one standard deviation (STD) calculated for each calendar month over the whole time series;
- −
- as the precipitation was not normally distributed, the threshold definition based on the STD exceeding was less informative. Therefore, only the quantile thresholds were applied for precipitation.
3. Results and Discussion
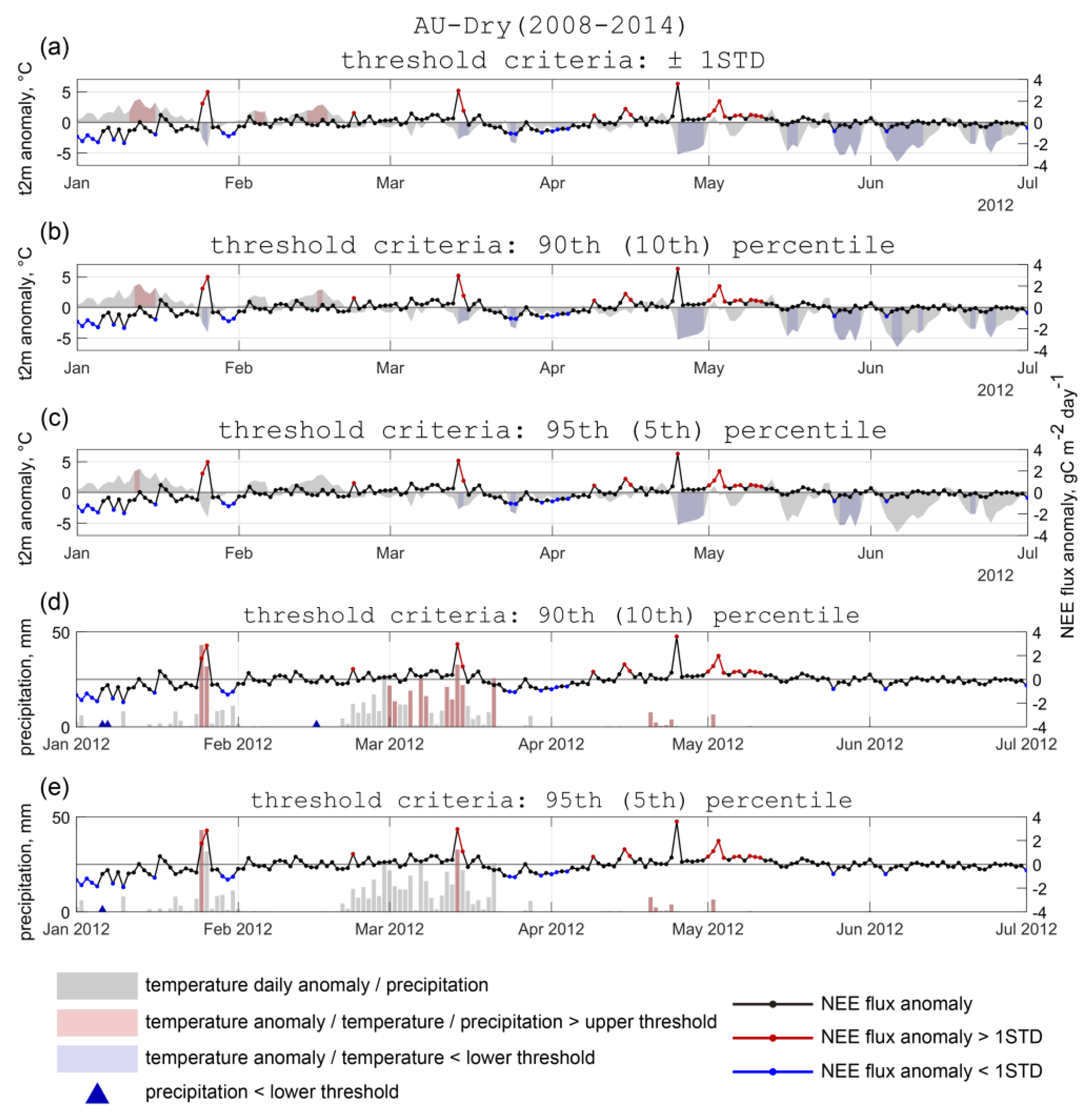
3.1. Temporal Variability of Daily Temperature, Precipitation, and CO2 Fluxes in Various Tropical Terrestrial Ecosystems
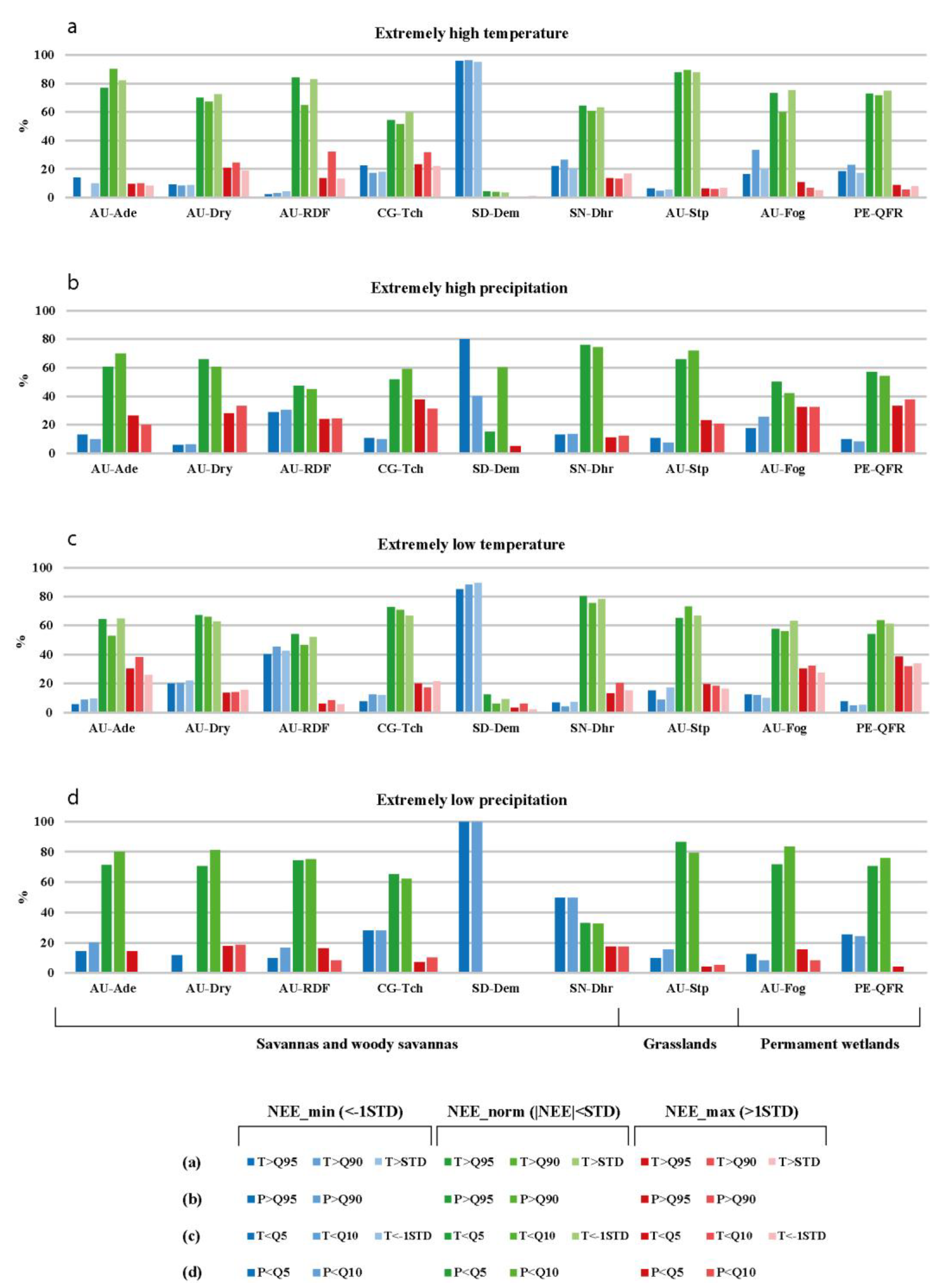
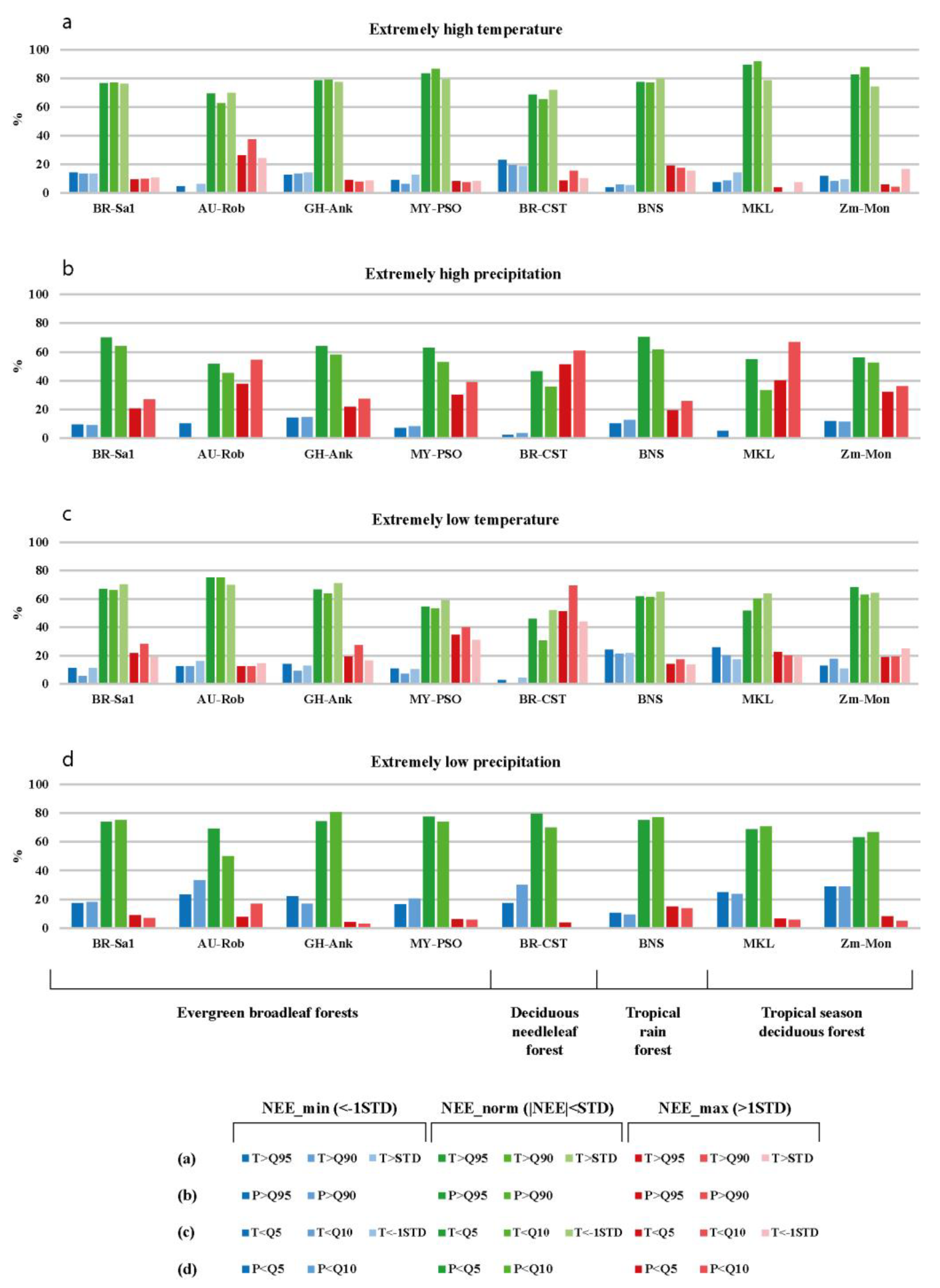
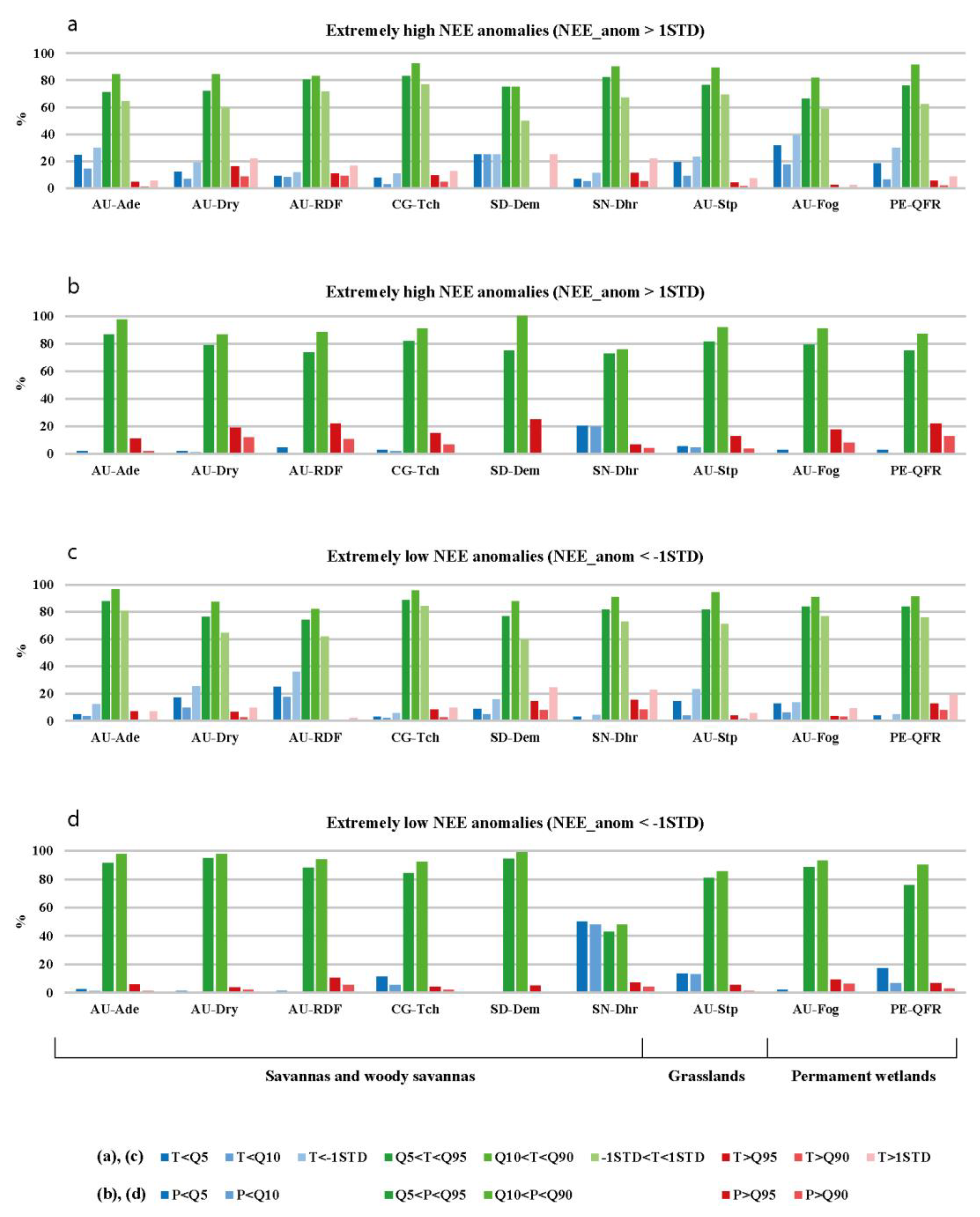
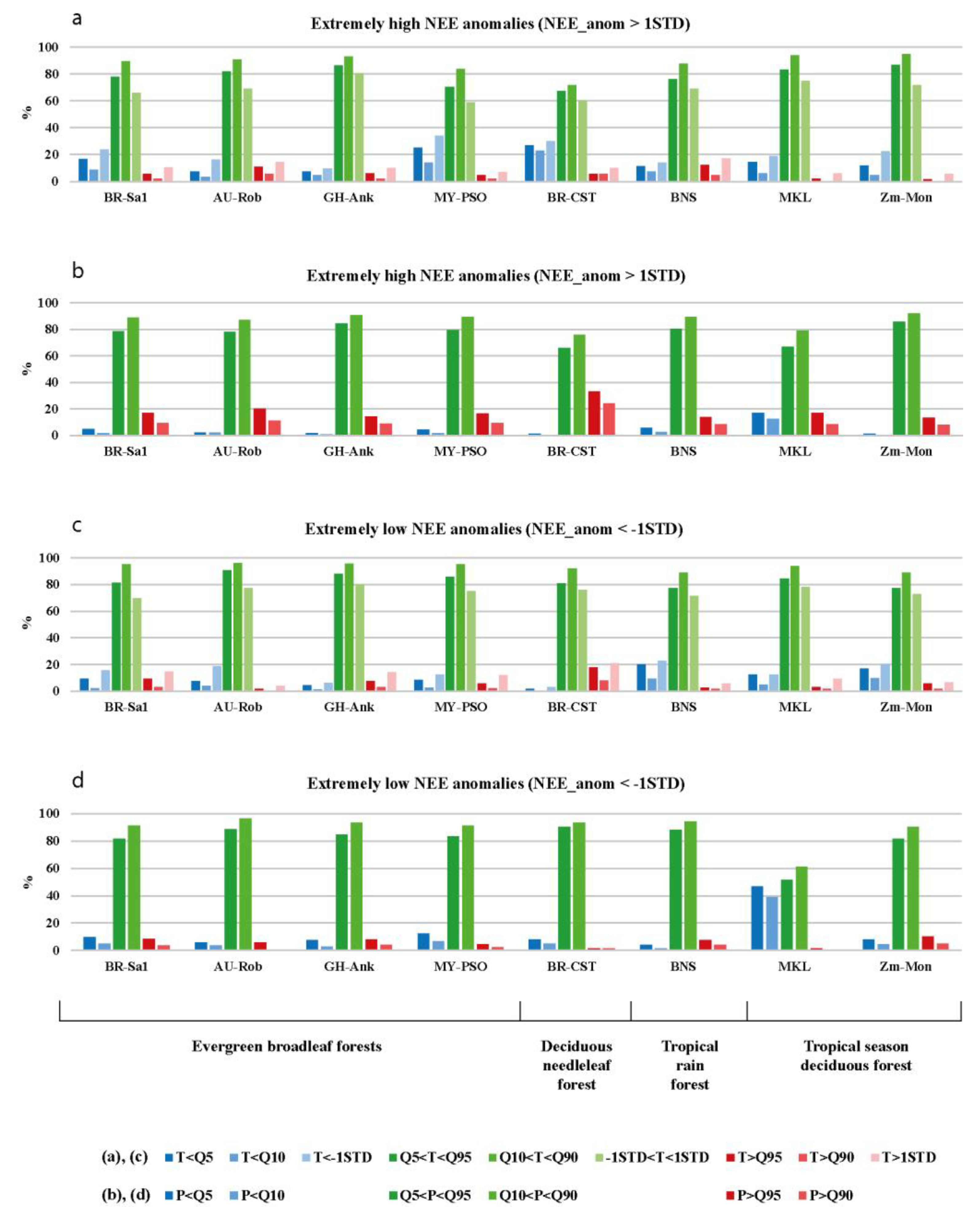
3.2. The Statistics of Relationships between Extreme Weather Conditions and Daily CO2 Flux Anomalies
3.2.1. Relationships between Daily Precipitation and CO2 Flux Anomalies
3.2.2. Relationships between Daily Air Temperature and CO2 Flux Anomalies
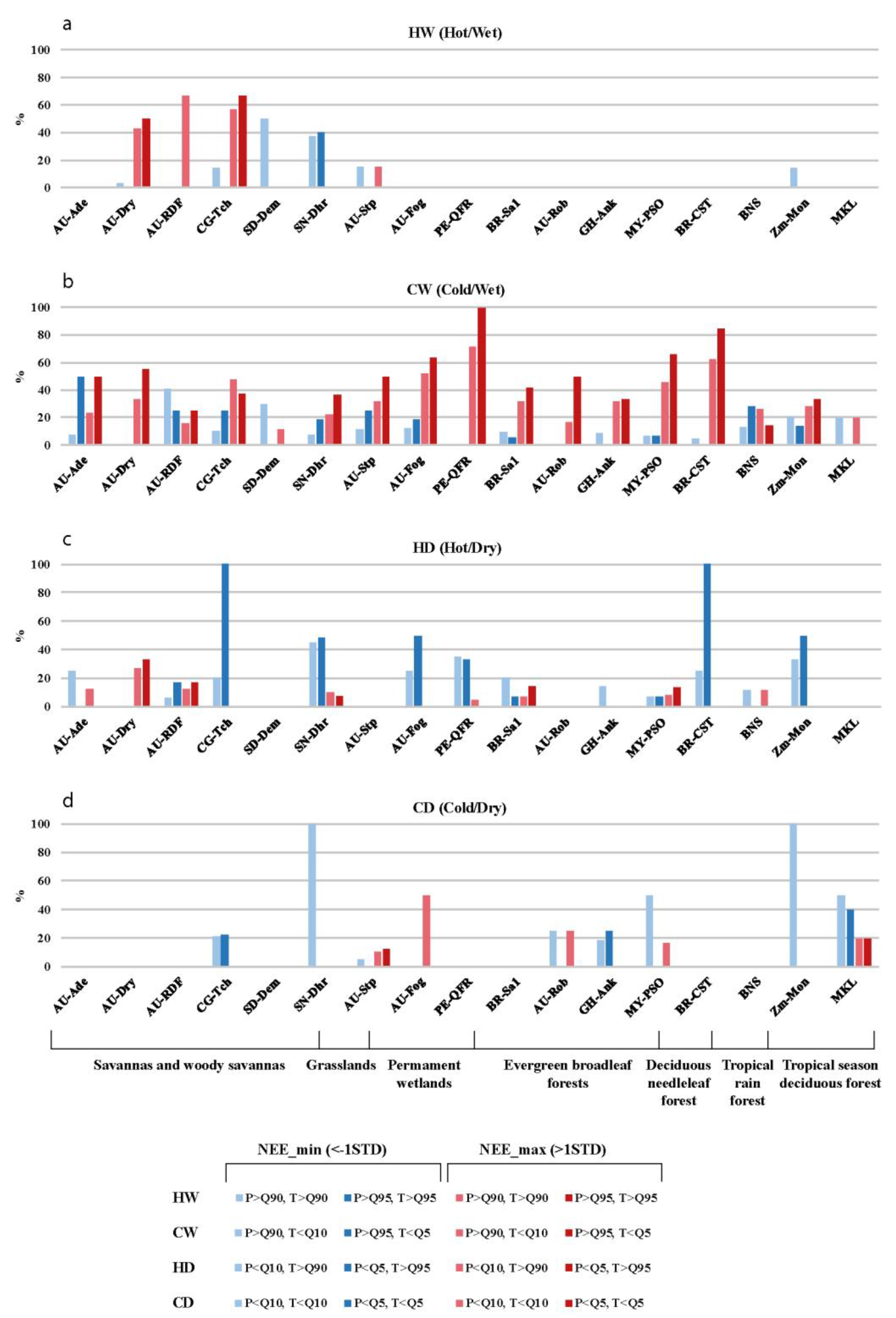
3.2.3. The Correlation between Daily CO2 Flux Anomalies and Temperature/Precipitation Extremes
3.2.4. Combined Effect of Temperature and Precipitation Extremes on Daily CO2 Fluxes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gulev, S.K.; Thorne, P.W.; Ahn, J.; Dentener, F.J.; Domingues, C.M.; Gerland, S.; Gong, D.; Kaufman, D.S.; Nnamchi, H.C.; Quaas, J.; et al. The Changing State of the Climate. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 287–422. ISBN 9781009157896. [Google Scholar]
- Seneviratne, S.I.; Zhang, X.; Adnan, M.; Badi, M.; Dereczynski, C.; Di Luca, A.; Ghosh, S.; Iskandar, I.; Kossin, J.; Lewis, S.; et al. Weather and Climate Extreme Events in a Changing Climate. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 1513–1766. ISBN 9781009157896. [Google Scholar]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef]
- Forzieri, G.; Girardello, M.; Ceccherini, G.; Spinoni, J.; Feyen, L.; Hartmann, H.; Beck, P.S.A.; Camps-Valls, G.; Chirici, G.; Mauri, A.; et al. Emergent vulnerability to climate-driven disturbances in European forests. Nat. Commun. 2021, 12, 1081. [Google Scholar] [CrossRef] [PubMed]
- Pugnaire, F.I.; Morillo, J.A.; Peñuelas, J.; Reich, P.B.; Bardgett, R.D.; Gaxiola, A.; Wardle, W.H.; van der Putten, W. Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci. Adv. 2019, 5, eaaz1834. [Google Scholar] [CrossRef] [PubMed]
- Ummenhofer, C.C.; Meeh, I.G.A. Extreme weather and climate events with ecological relevance: A review. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160135. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Root, T.L.; Willig, M.R. Impacts of extreme weather and climate on terrestrial biota. Bull. Am. Meteorol. Soc. 2000, 81, 443–450. [Google Scholar] [CrossRef]
- Zscheischler, J.; Michalak, A.M.; Schwalm, C.; Mahecha, M.D.; Huntzinger, D.N.; Reichstein, M.; Berthier, G.; Ciais, P.; Cook, R.B.; El-Masri, B.; et al. Impact of Large-Scale Climate Extremes on Biospheric Carbon Fluxes: An Intercomparison Based on MsTMIP Data. Glob. Biogeochem. Cycles 2014, 28, 585–600. [Google Scholar] [CrossRef]
- Forzieri, G.; Dakos, V.; McDowell, N.G.; Ramdane, A.; Cescatti, A. Emerging signals of declining forest resilience under climate change. Nature 2022, 608, 534–539. [Google Scholar] [CrossRef]
- Russell, A.E.; Parton, W.J., Jr. Modeling the Effects of Global Change on Ecosystem Processes in a Tropical Rainforest. Forests 2020, 11, 213. [Google Scholar] [CrossRef]
- Grace, J.; Lloyd, J.; McIntyre, J.; Miranda, A.; Meir, P.; Miranda, H.; Nobre, C.; Moncrieff, J.B.; Massheder, J.M.; Malhi, Y.; et al. Carbon dioxide uptake by an undisturbed tropical rain forest in south-west Amazonia, 1992 to 1993. Science 1995, 270, 778–780. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragao, L.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; Lopez-Gonzalez, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef]
- Ciais, P.; Piao, S.L.; Cadule, P.; Friedlingstein, P.; Chedin, A. Variability and recent trends in the African terrestrial carbon balance. Biogeosciences 2009, 6, 1935–1948. [Google Scholar] [CrossRef]
- Olchev, A.; Ibrom, A.; Panferov, O.; Gushchina, D.; Kreilein, H.; Popov, V.; Propastin, P.; June, T.; Rauf, A.; Gravenhorst, G.; et al. Response of CO2 and H2O fluxes in a mountainous tropical rainforest in equatorial Indonesia to El Niño events. Biogeosciences 2015, 12, 6655–6667. [Google Scholar] [CrossRef]
- Malhi, Y. The carbon balance of tropical forest regions, 1990–2005. Curr. Opin. Environ. Sustain. 2010, 2, 237–244. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Busman, N.A.; Melling, L.; Goh, K.J.; Imran, Y.; Sangok, F.E.; Watanabe, A. Soil CO2 and CH4 fluxes from different forest types in tropical peat swamp forest. Sci. Total Environ. 2023, 858, 159973. [Google Scholar] [CrossRef]
- Malhi, Y.; Baldocchi, D.D.; Jarvis, P.G. The carbon balance of tropical, temperate and boreal forests. Plant Cell Environ. 1999, 22, 715–740. [Google Scholar] [CrossRef]
- Wood, T.E.; Cavaleri, M.A.; Reed, S.C. Tropical Forest carbon balance in a warmer world: A critical review spanning microbial- to ecosystem-scale processes. Biol. Rev. 2012, 87, 912–927. [Google Scholar] [CrossRef]
- Alkama, R.; Cescatti, A. Biophysical climate impacts of recent changes in global forest cover. Science 2016, 351, 600–604. [Google Scholar] [CrossRef]
- Artaxo, P.; Hansson, H.C.; Machado, L.A.T.; Rizzo, L.V. Tropical forests are crucial in regulating the climate on Earth. PLoS Clim. 2022, 1, e0000054. [Google Scholar] [CrossRef]
- Andersen, A. Net photosynthesis as an indicator of plant response to environmental influence. Acta Hortic. 1976, 64, 133–138. [Google Scholar] [CrossRef]
- Schurr, U.; Walter, A.; Rascher, U. Functional dynamics of plant growth and photosynthesis—From steady-state to dynamics—From homogeneity to heterogeneity. Plant Cell Environ. 2006, 29, 340–352. [Google Scholar] [CrossRef] [PubMed]
- The Data Portal Serving the FLUXNET Community. Available online: https://fluxnet.org/data/ (accessed on 5 October 2022).
- Granier, A.; Reichstein, M.; Bréda, N.; Janssens, I.A.; Falge, E.; Ciais, P.; Grünwald, T.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; et al. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003. Agric. For. Meteorol. 2007, 143, 123–145. [Google Scholar] [CrossRef]
- Hiyama, T.; Ueyama, M.; Kotani, A.; Iwata, H.; Nakai, T.; Okamura, M.; Ohta, T.; Harazono, Y.; Petrov, R.E.; Maximov, T.C. Lessons learned from more than a decade of greenhouse gas flux measurements at boreal forests in eastern Siberia and interior Alaska. Polar Sci. 2021, 27, 100607. [Google Scholar] [CrossRef]
- Gatti, L.V.; Gloor, M.; Miller, J.B.; Doughty, C.E.; Malhi, Y.; Domingues, L.G.; Basso, L.S.; Martinewski, A.; Correia, C.S.C.; Borges, V.F.; et al. Drought sensitivity of Amazonian carbon balance revealed by atmospheric measurements. Nature 2014, 506, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.; Metcalfe, D.B.; Girardin, C.A.J.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.E.; Araujo-Murakami, A.; da Costa, M.C.; Rocha, W.; et al. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Gushchina, D.; Zheleznova, I.; Osipov, A.; Olchev, A. Effect of various types of ENSO events on moisture conditions in the humid and sub-humid tropics. Atmosphere 2020, 11, 1354. [Google Scholar] [CrossRef]
- Lindroth, A.; Holst, J.; Linderson, M.-L.; Aurela, M.; Biermann, T.; Heliasz, M.; Chi, J.; Ibrom, A.; Kolari, P.; Klemedtsson, L.; et al. Effects of drought and meteorological forcing on carbon and water fluxes in Nordic forests during the dry summer of 2018. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190516. [Google Scholar] [CrossRef]
- Gushchina, D.; Heimsch, F.; Osipov, A.; June, T.; Rauf, A.; Kreilein, H.; Panferov, O.; Olchev, A.; Knohl, A. Effects of the 2015–2016 El Niño event on energy and CO2 fluxes of a tropical rainforest in Central Sulawesi. Geogr. Environ. Sustain. 2019, 12, 183–196. [Google Scholar] [CrossRef]
- Zscheischler, J.; Reichstein, M.; Harmeling, S.; Rammig, A.; Tomelleri, E.; Mahecha, M.D. Extreme Events in Gross Primary Production: A Characterization across Continents. Biogeosciences 2014, 11, 2909–2924. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial Gross Carbon Dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Mahecha, M.D.; Reichstein, M.; Carvalhais, N.; Lasslop, G.; Lange, H.; Seneviratne, S.I.; Vargas, R.; Ammann, C.; Arain, M.A.; Cescatti, A.; et al. Global Convergence in the Temperature Sensitivity of Respiration at Ecosystem Level. Science 2010, 329, 838–840. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D. An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Pennington, R.T.; Lehmann, C.E.R.; Rowland, L.M. Tropical savannas and dry forests. Curr. Biol. 2018, 28, R541–R545. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Medium-Range Weather Forecasts. Available online: https://www.ecmwf.int/en/forecasts/datasets/reanalysis-datasets/era5 (accessed on 23 September 2022).
- Aubinet, M.; Vesala, T.; Papale, D. Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Springer: Dordrecht, The Netherlands, 2012; p. 438. ISBN 9789400723504. [Google Scholar]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Zheleznova, I.V.; Gushchina, D.Y. Variability of extreme air temperatures and precipitation in different natural zones in late XX and early XXI centuries according to ERA5 reanalysis data. Izv. Atmos. Ocean. Phys. 2023; in press. [Google Scholar]
- McCulley, R.L.; Boutton, T.W.; Archer, S.R. Soil Respiration in a Subtropical Savanna Parkland: Response to Water Additions. Soil Sci. Soc. Am. J. 2007, 7, 820–828. [Google Scholar] [CrossRef]
- Birch, H.F. Mineralisation of plant nitrogen following alternate wet and dry conditions. Plant Soil 1964, 20, 43–49. [Google Scholar] [CrossRef]
- Jarvis, P.; Rey, A.; Petsikos, C.; Wingate, L.; Rayment, M.; Pereira, J.; Banza, J.; David, J.; Miglietta, F.; Borghetti, M.; et al. Drying and Wetting of Mediterranean Soils Stimulates Decomposition and Carbon Dioxide Emission: The “Birch Effect”. Tree Physiol. 2007, 27, 929–940. [Google Scholar] [CrossRef]
- Roby, M.C.; Scott, R.L.; Biederman, J.A.; Smith, W.K.; Moore, D.J.P. Response of soil carbon dioxide efflux to temporal repackaging of rainfall into fewer, larger events in a semiarid grassland. Front. Environ. Sci. 2022, 10, 940943. [Google Scholar] [CrossRef]
- Devi Kanniah, K.; Beringer, J.; Hutley, L.B. The comparative role of key environmental factors in determining savanna productivity and carbon fluxes: A review, with special reference to northern Australia. Prog. Phys. Geogr. Earth Environ. 2010, 34, 459–490. [Google Scholar] [CrossRef]
- Hutley, L.B.; Beringer, J.; Fatichi, S.; Schymanski, S.J.; Northwood, M. Gross primary productivity and water use efficiency are increasing in a high rainfall tropical savanna. Glob. Chang. Biol. 2022, 28, 2360–2380. [Google Scholar] [CrossRef] [PubMed]
- Kutsch, W.L.; Hanan, N.; Scholes, B.; McHugh, I.; Kubheka, W.; Eckhardt, H.; Williams, C. Response of carbon fluxes to water relations in a savanna ecosystem in South Africa. Biogeosciences 2008, 5, 1797–1808. [Google Scholar] [CrossRef]
- Yu, J.-C.; Chiang, P.-N.; Lai, Y.-J.; Tsai, M.-J.; Wang, Y.-N. High Rainfall Inhibited Soil Respiration in an Asian Monsoon Forest in Taiwan. Forests 2021, 12, 239. [Google Scholar] [CrossRef]
- Stewart, I.T.; Maurer, E.P.; Stahl, K.; Joseph, K. Recent evidence for warmer and drier growing seasons in climate sensitive regions of Central America from multiple global datasets. Int. J. Climatol. 2022, 42, 1399–1417. [Google Scholar] [CrossRef]
- Ibrom, A.; Olchev, A.; June, T.; Ross, T.; Kreilein, H.; Falk, U.; Merklein, J.; Twele, A.; Rakkibu, G.; Grote, S.; et al. Effects of land-use change on matter and energy exchange between ecosystems in the rain forest margin and the atmosphere. In The Stability of Tropical Rainforest Margins-Linking Ecological, Economic and Social Constraints; Tscharntke, T., Leuschner, C., Zeller, M., Guhardja, E., Bidin, A., Eds.; Springer: Berlin, Germany, 2007; pp. 463–492. [Google Scholar] [CrossRef]
- Madani, N.; Kimball, J.S.; Parazoo, N.C.; Ballantyne, A.P.; Tagesson, T.; Jones, L.A.; Reichle, R.H.; Palmer, P.I.; Velicogna, I.; Bloom, A.A. Below-surface water mediates the response of African forests to reduced rainfall. Environ. Res. Lett. 2020, 15, 034063. [Google Scholar] [CrossRef]
- Li, L.; Fan, W.; Kang, X.; Wang, Y.; Cui, X.; Xu, C.; Griffin, K.L.; Hao, Y. Responses of Greenhouse Gas Fluxes to Climate Extremes in a Semiarid Grassland. Atmos. Environ. 2016, 142, 32–42. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of Plant Responses to Water Stress and Related Genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Miller, S.D.; Goulden, M.L.; Hutyra, L.R.; Keller, M.; Saleska, S.R.; Wofsy, S.C.; Figueira, A.M.S.; da Rocha, H.R.; de Camargo, P.B. Reduced impact logging minimally alters tropical rainforest carbon and energy exchange. Proc. Natl. Acad. Sci. USA 2011, 108, 9431–9435. [Google Scholar] [CrossRef]
- Mills, M.B.; Malhi, Y.; Ewers, R.M.; Kho, L.K.; Teh, Y.A.; Both, S.; Burslem, D.F.R.; Majalap, N.; Nilus, R.; Huasco, W.H.; et al. Tropical forests post-logging are a persistent net carbon source to the atmosphere. Proc. Natl. Acad. Sci. USA 2023, 120, e2214462120. [Google Scholar] [CrossRef]
- Wild, B.; Teubner, I.; Moesinger, L.; Zotta, R.-M.; Forkel, M.; van der Schalie, R.; Sitch, S.; Dorigo, W. VODCA2GPP—A new, global, long-term (1988--2020) gross primary production dataset from microwave remote sensing. Earth Syst. Sci. Data 2022, 14, 1063–1085. [Google Scholar] [CrossRef]
- Yang, Z.; Luo, X.; Shi, Y.; Zhou, T.; Luo, K.; Lai, Y.; Yu, P.; Liu, L.; Olchev, A.; Bond-Lamberty, B.; et al. Controls and variability of soil respiration temperature sensitivity across China. Sci. Total Environ. 2023, 871, 161974. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Eamus, D.; Hutley, L.B. Seasonal patterns of soil carbon dioxide efflux from a wet-dry tropical savanna of northern Australia. Aust. J. Bot. 2002, 50, 43–51. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Hutley, L.B.; Beringer, J.; Holtum, J.A.M.; Turner, B.L. Photosynthetic physiology of eucalypts along a sub-continental rainfall gradient in northern Australia. Agric. For. Meteorol. 2011, 151, 1462–1470. [Google Scholar] [CrossRef]
- Martínez, B.; Sanchez-Ruiz, S.; Gilabert, M.A.; Moreno, A.; Campos-Taberner, M.; García-Haro, F.J.; Trigo, I.F.; Aurela, M.; Brümmer, C.; Carrara, A.; et al. Retrieval of daily gross primary production over Europe and Africa from an ensemble of SEVIRI/MSG products. Int. J. Appl. Earth Obs. Geoinf. 2018, 65, 124–136. [Google Scholar] [CrossRef]
- Hanke, H.; Borjeson, L.; Hylander, K.; Enfors-Kautsky, E. Drought Tolerant Species Dominate as Rainfall and Tree Cover Returns in the West African Sahel. Land Use Policy 2016, 59, 111–120. [Google Scholar] [CrossRef]
- Beringer, J.; Livesley, S.J.; Randle, J.; Hutley, L.B. Carbon dioxide fluxes dominate the greenhouse gas exchanges of a seasonal wetland in the wet–dry tropics of northern Australia. Agric. For. Meteorol. 2013, 182–183, 239–247. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Tropical Ecosystems | Stations | Correlation Coefficients between Daily Temperature, Precipitation, and CO2 Fluxes | ||||
|---|---|---|---|---|---|---|
| T STD | T Q90 | T Q95 | P Q90 | P Q95 | ||
| Savannas | AU-Ade | −0.17 | −0.08 | 0.11 | 0.14 | 0.27 |
| AU-Dry | 0.12 | 0.04 | 0.11 | 0.14 | 0.17 | |
| AU-RDF | 0.33 | −0.40 | −0.32 | 0.16 | 0.17 | |
| CG-Tch | −0.07 | 0.02 | 0.05 | 0.14 | 0.20 | |
| SD-Dem | −0.07 | 0.25 | 0.34 | 0.06 | −0.03 | |
| SN-Dhr | −0.18 | −0.23 | −0.30 | 0.18 | 0.14 | |
| Evergreen broadleaf forests | BR-Sa1 | −0.14 | 0.04 | 0.04 | 0.16 | 0.23 |
| AU-Rob | 0.34 | 0.39 | 0.01 | 0.71 | 0.81 | |
| GH-Ank | −0.10 | 0.29 | 0.25 | −0.14 | −0.21 | |
| MY-PSO | −0.30 | −0.11 | −0.14 | 0.33 | 0.37 | |
| Grasslands | AU-Stp | 0.04 | −0.04 | 0.06 | −0.09 | −0.11 |
| Permanent wetlands | AU-Fog | −0.13 | −0.41 | −0.32 | 0.09 | 0.11 |
| PE-QFR | −0.33 | 0.07 | −0.17 | 0.17 | 0.23 | |
| Deciduous needleleaf forest | BR-CST | −0.51 | −0.17 | −0.17 | 0.16 | 0.31 |
| Tropical rain forest | BNS | 0.12 | −0.05 | 0.07 | 0.07 | −0.03 |
| Tropical season deciduous forest | MKL | −0.04 | −0.04 | 0.09 | −0.07 | −0.17 |
| Zm-Mon | −0.13 | −0.06 | −0.16 | 0.12 | 0.23 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gushchina, D.; Tarasova, M.; Satosina, E.; Zheleznova, I.; Emelianova, E.; Novikova, E.; Olchev, A. Effects of Extreme Temperature and Precipitation Events on Daily CO2 Fluxes in the Tropics. Climate 2023, 11, 117. https://doi.org/10.3390/cli11060117
Gushchina D, Tarasova M, Satosina E, Zheleznova I, Emelianova E, Novikova E, Olchev A. Effects of Extreme Temperature and Precipitation Events on Daily CO2 Fluxes in the Tropics. Climate. 2023; 11(6):117. https://doi.org/10.3390/cli11060117
Chicago/Turabian StyleGushchina, Daria, Maria Tarasova, Elizaveta Satosina, Irina Zheleznova, Ekaterina Emelianova, Elena Novikova, and Alexander Olchev. 2023. "Effects of Extreme Temperature and Precipitation Events on Daily CO2 Fluxes in the Tropics" Climate 11, no. 6: 117. https://doi.org/10.3390/cli11060117
APA StyleGushchina, D., Tarasova, M., Satosina, E., Zheleznova, I., Emelianova, E., Novikova, E., & Olchev, A. (2023). Effects of Extreme Temperature and Precipitation Events on Daily CO2 Fluxes in the Tropics. Climate, 11(6), 117. https://doi.org/10.3390/cli11060117