
Projections of Future Suitable Bioclimatic Conditions of Parthenogenetic Whiptails


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
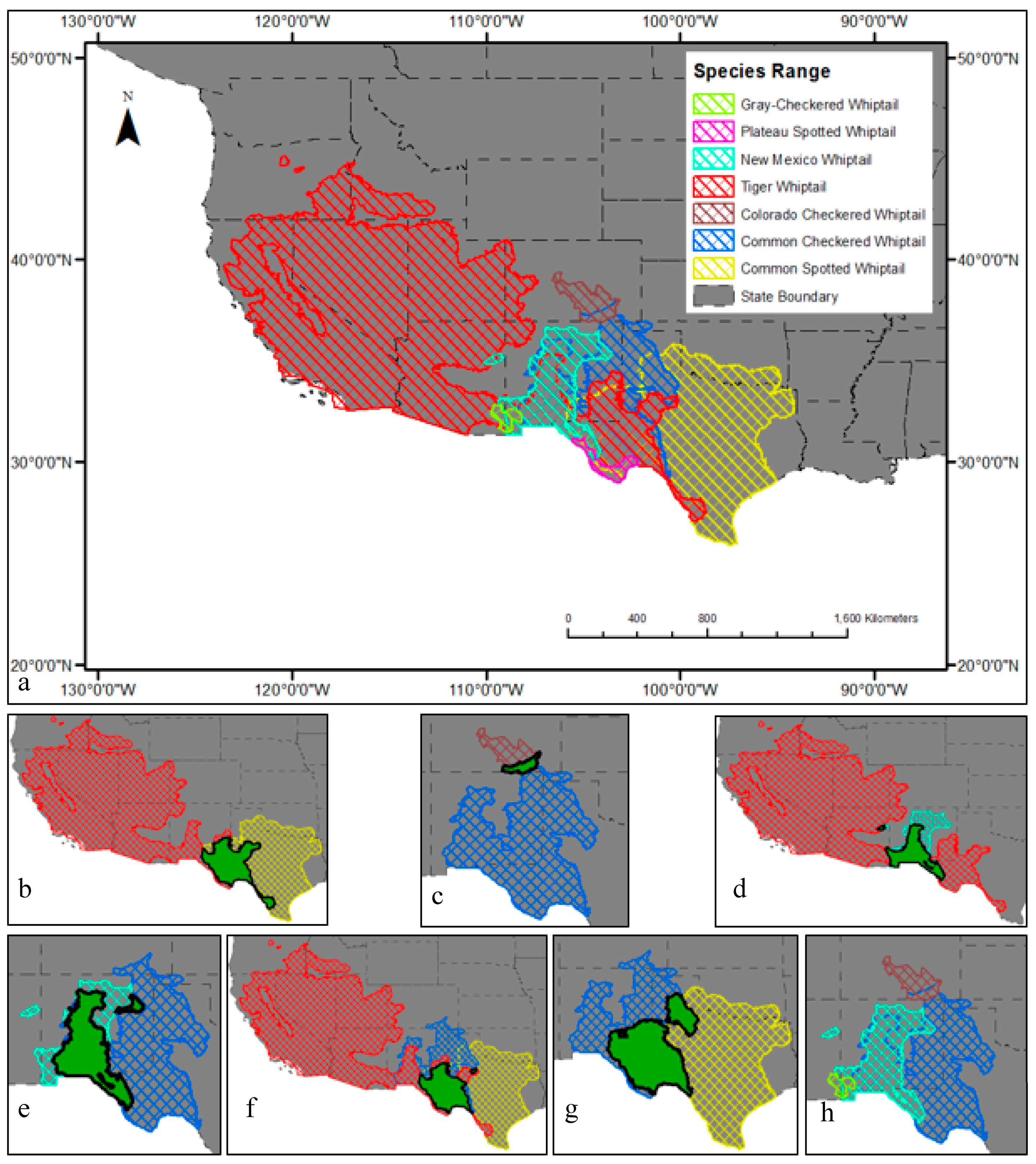
2.2. Occurence Datasets
2.3. Species Distribution Modeling
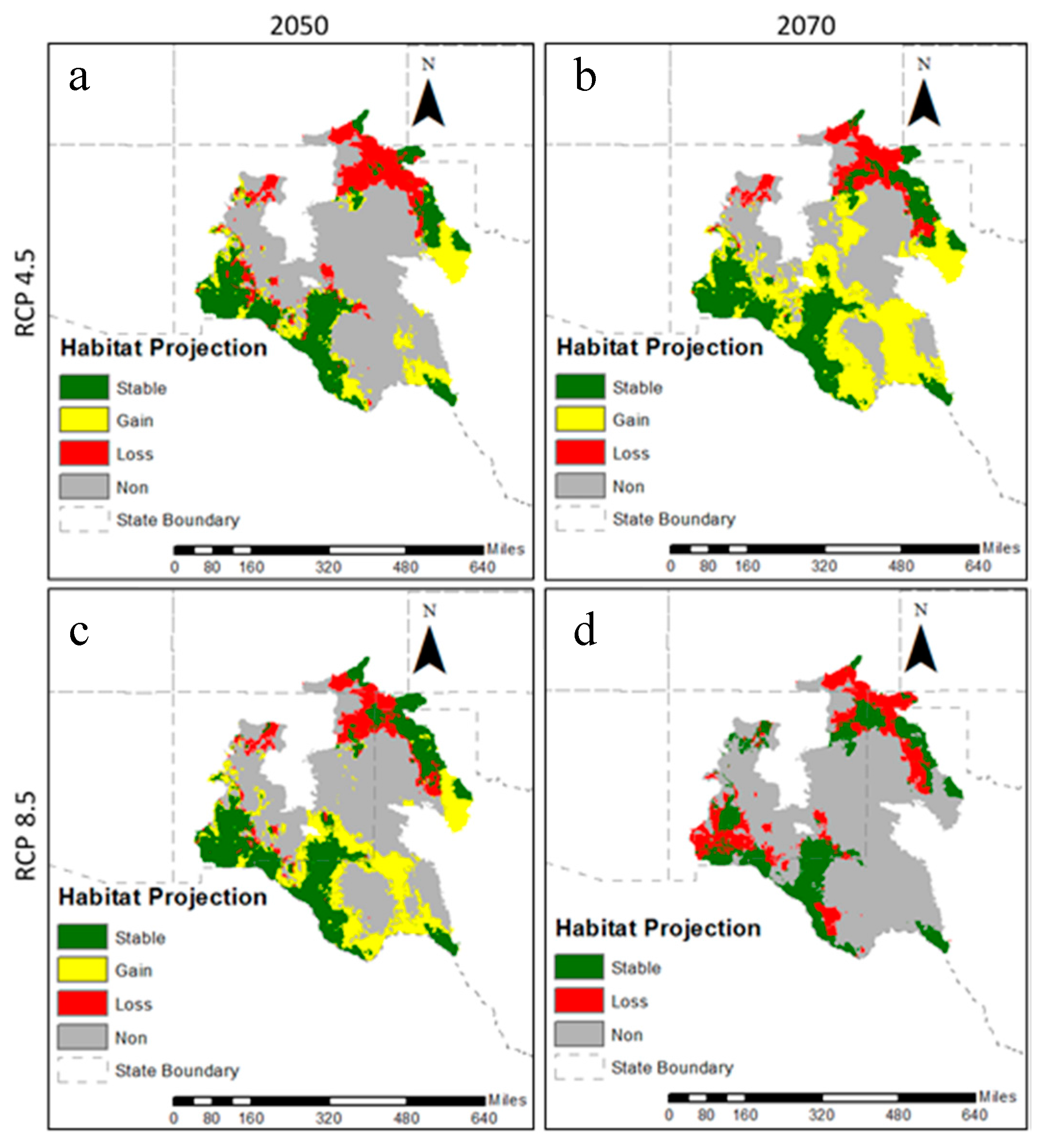
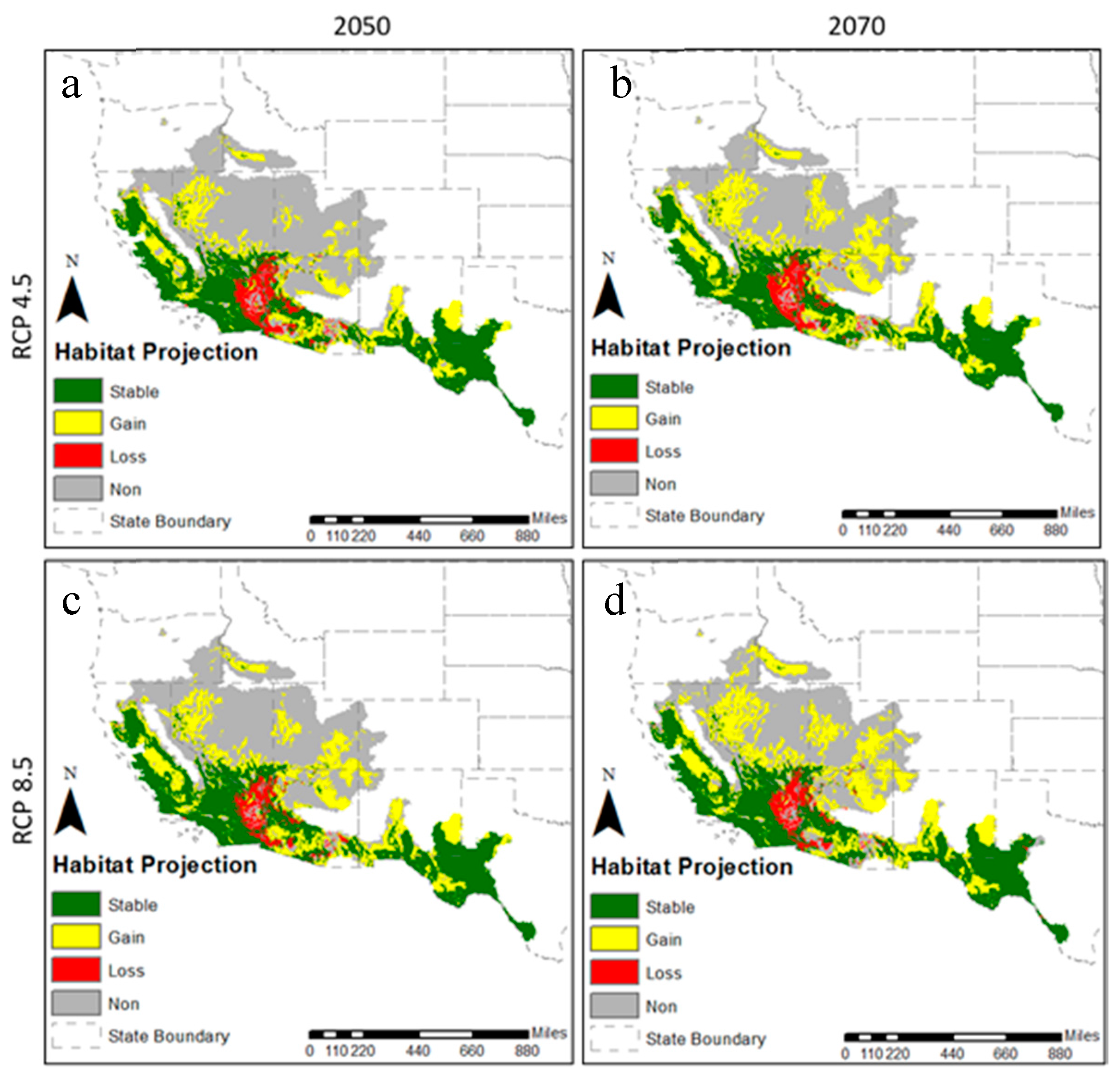
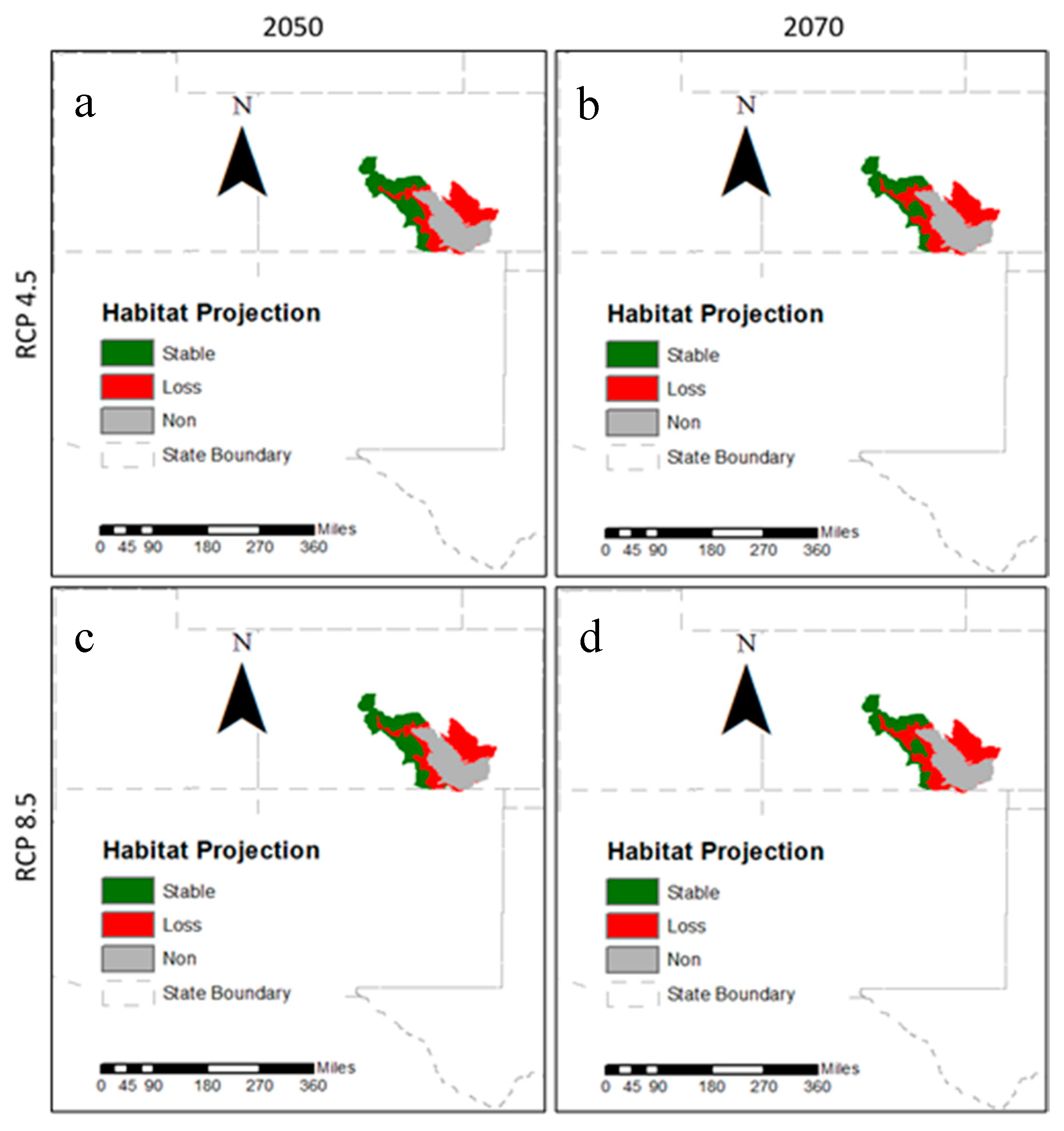
2.4. Projection to Future Conditions
3. Results
3.1. Individual Model Performance
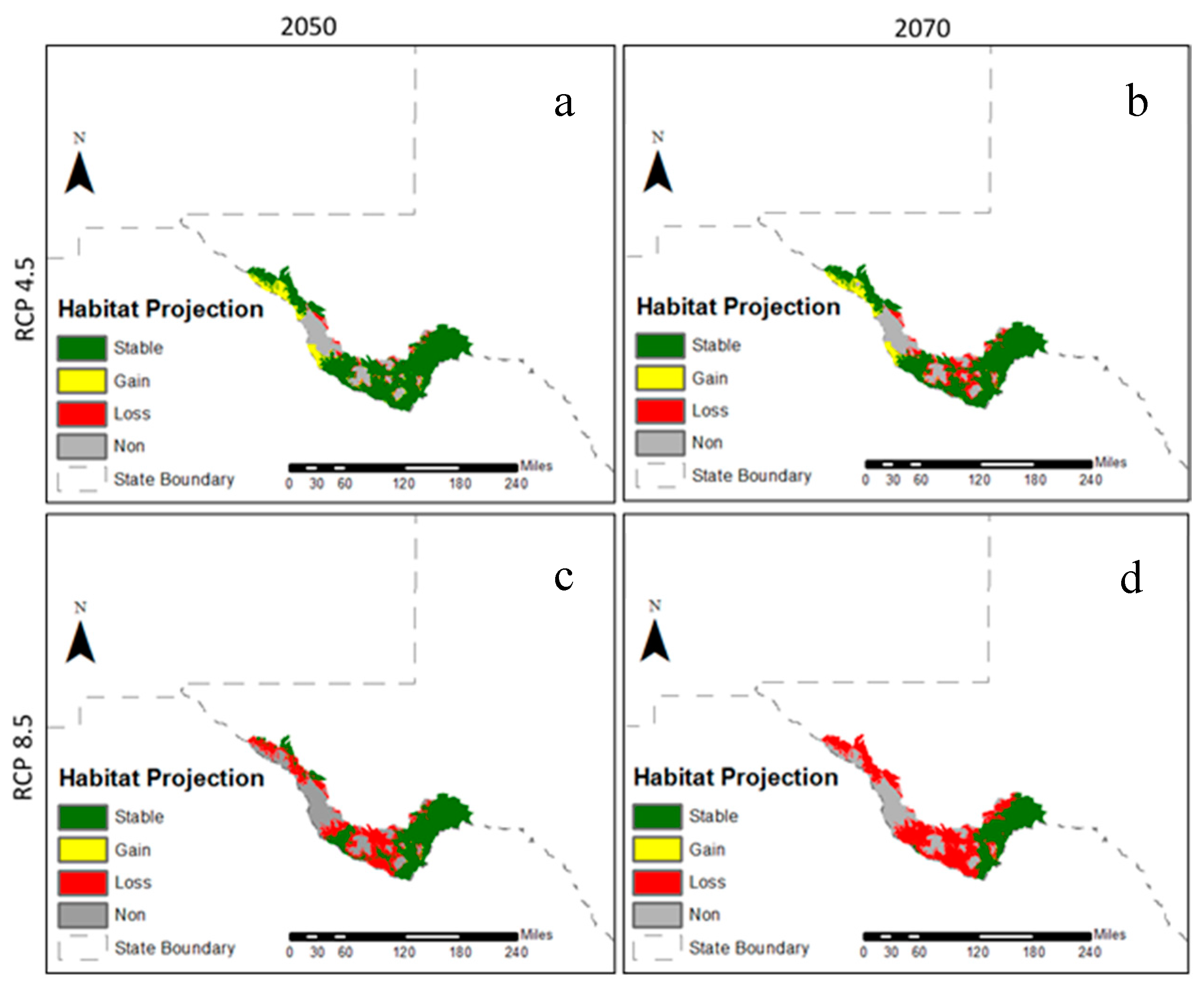
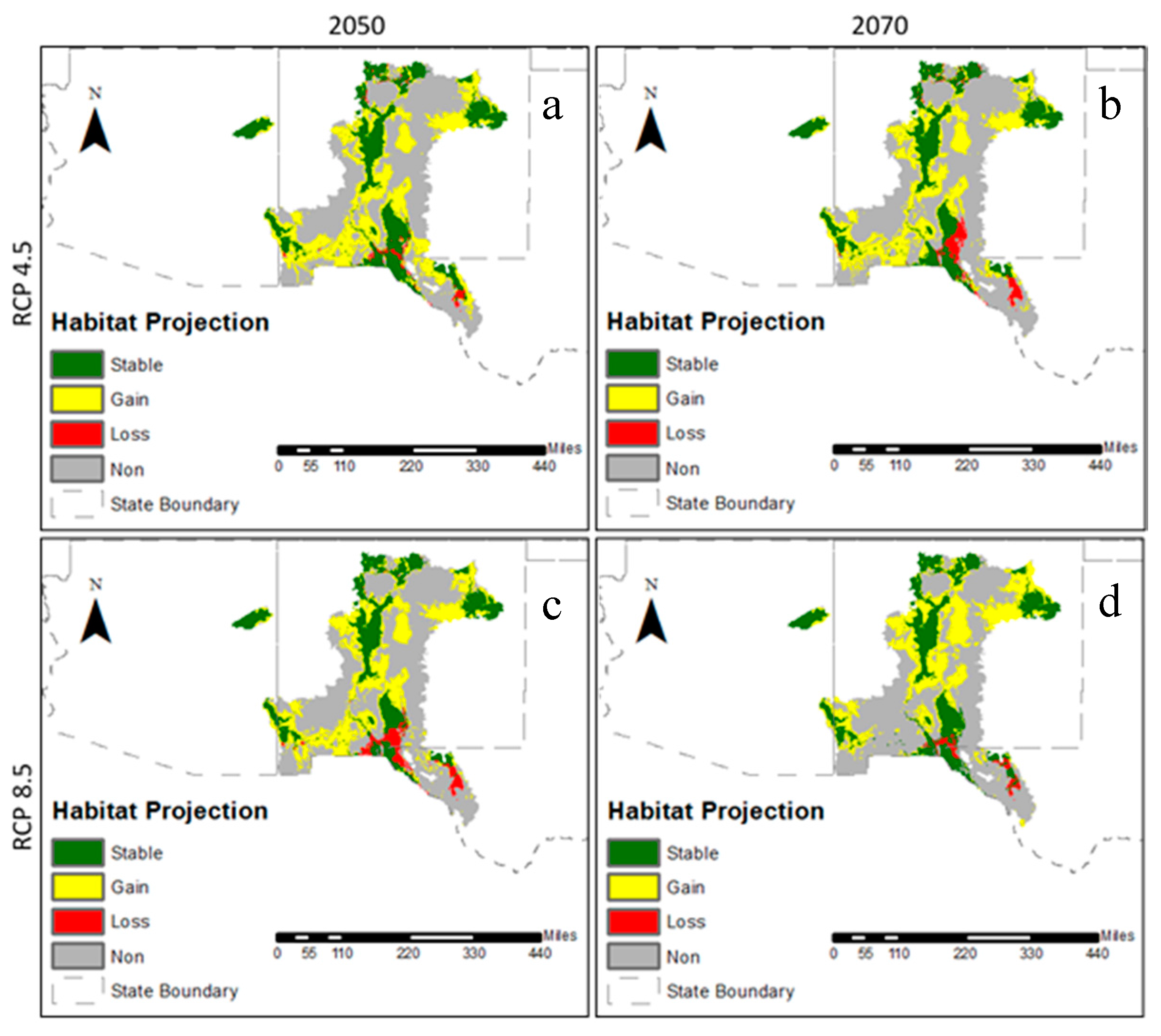
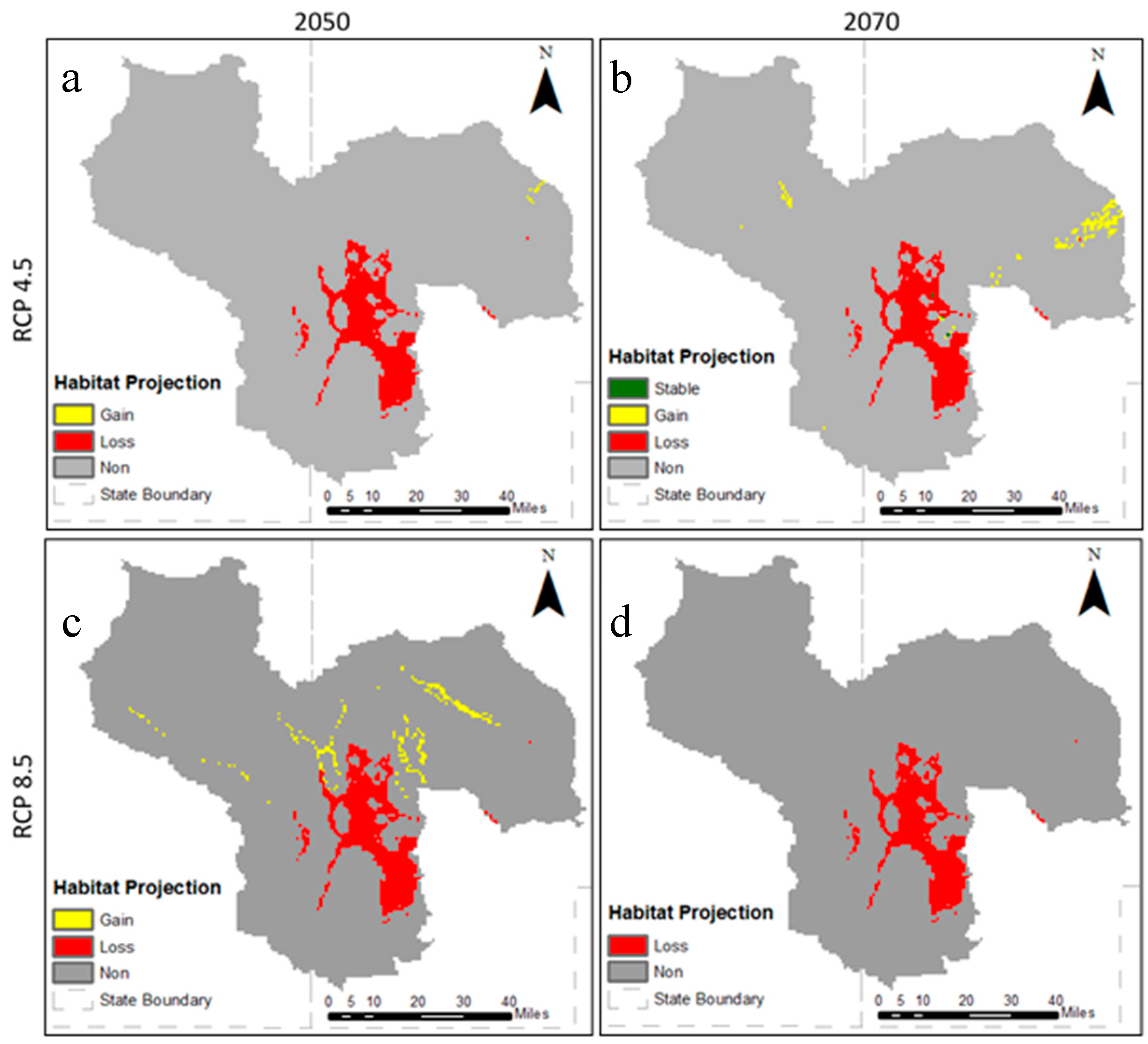
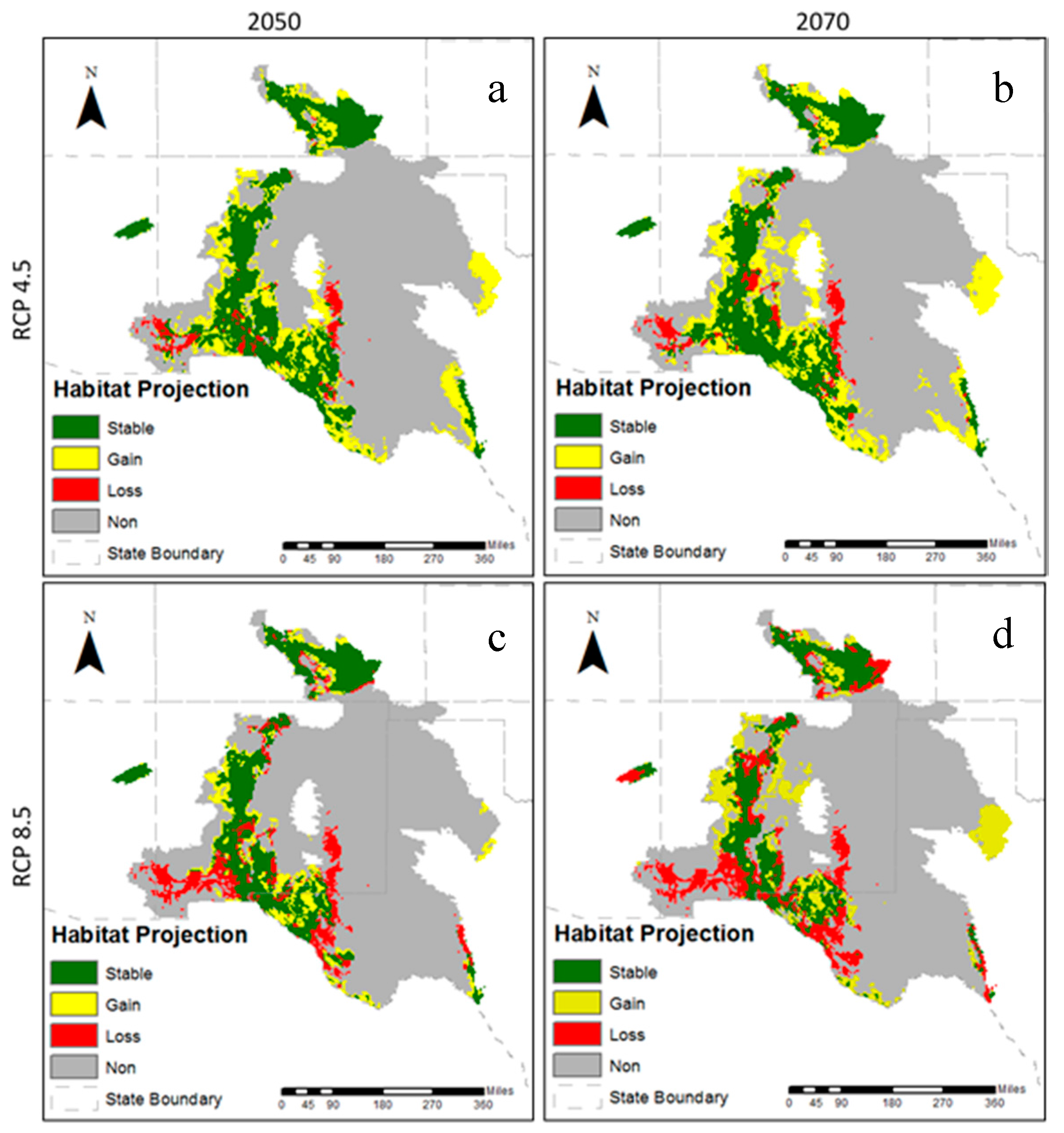
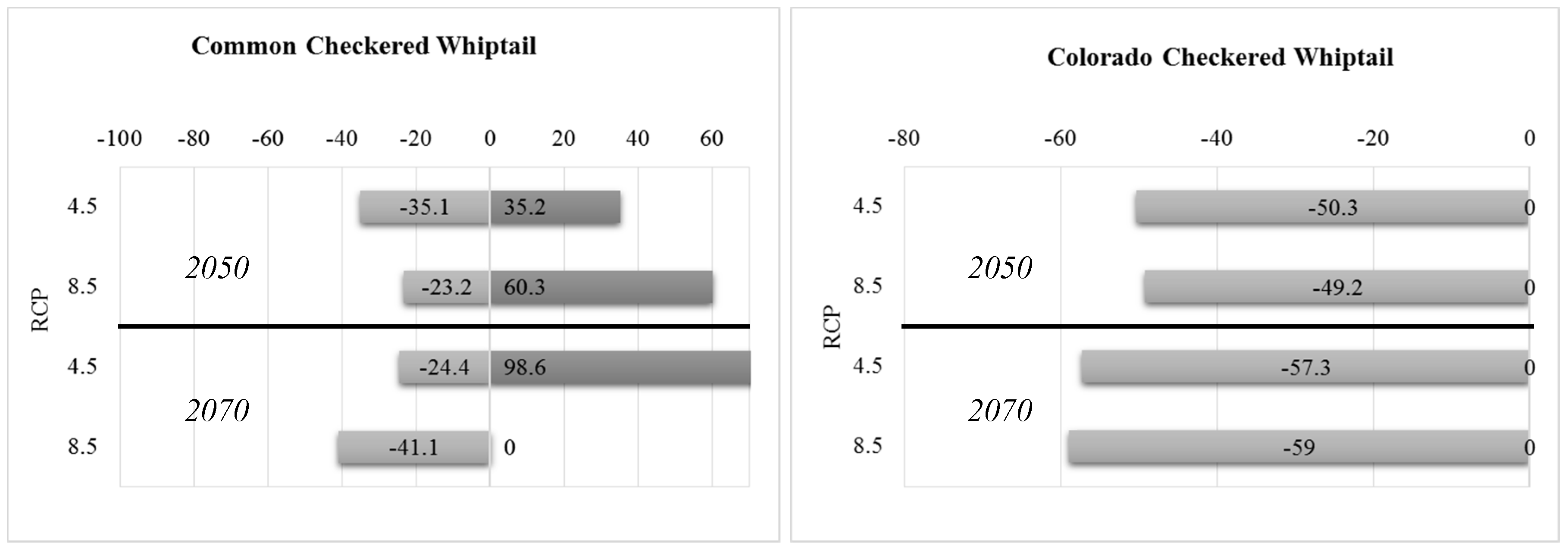
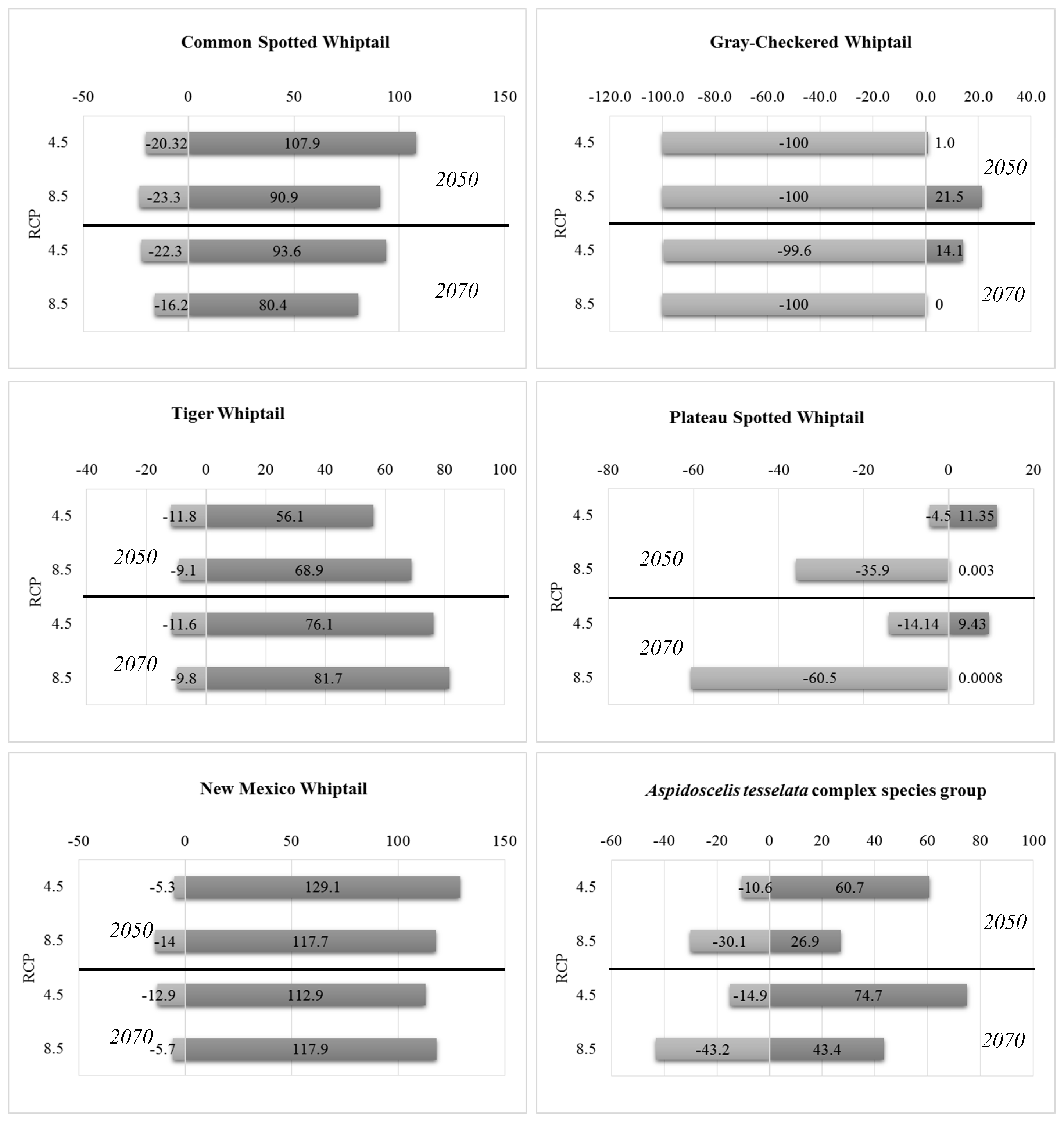
3.2. Future Climatic Envelope
4. Discussion
Caveats
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Species Use |
|---|---|---|
| Bioclim 1 | Annual Mean Temperature | |
| Bioclim 2 | Mean Diurnal Range | |
| Bioclim 3 | Isothermality | CW, TW, CCW, GCW |
| Bioclim 4 | Temperature Seasonality | |
| Bioclim 5 | Maximum Temperature Warmest Month | |
| Bioclim 6 | Minimum Temperature Coldest Month | CCW, PSW, CSW |
| Bioclim 7 | Temperature Annual Range | NMW |
| Bioclim 8 | Mean Temperature of Wettest Quarter | TW, CSW, NMW |
| Bioclim 9 | Mean Temperature of Driest Quarter | CW, TW |
| Bioclim 10 | Mean Temperature of Warmest Quarter | CW, TW, CSW |
| Bioclim 11 | Mean Temperature of Coldest Quarter | GCW |
| Bioclim 12 | Annual Precipitation | CW, TW, CSW |
| Bioclim 13 | Precipitation of Wettest Month | |
| Bioclim 14 | Precipitation of Driest Month | CCW |
| Bioclim 15 | Precipitation Seasonality | CW, PSW, PSW, NMW, GCW |
| Bioclim 16 | Precipitation of Wettest Quarter | |
| Bioclim 17 | Precipitation of Driest Quarter | |
| Bioclim 18 | Precipitation of Warmest Quarter | TW, PSW, NMW |
| Bioclim 19 | Precipitation of Coldest Quarter | GCW |
References
- Buckley, L.B.; Jetz, W. Environmental and historical constraints on global patterns of amphibian richness. Proc. R. Soc. B Biol. Sci. 2007, 274, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Sexton, J.P.; McIntyre, P.J.; Angert, A.L.; Rice, K.J. Evolution and ecology of species range limits. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 415–436. [Google Scholar] [CrossRef]
- Thomas, C.D. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Thuiller, W.; Pearson, R.G. Climate warming and the decline of amphibians and reptiles in Europe. J. Biogeogr. 2006, 33, 1712–1728. [Google Scholar] [CrossRef]
- Lannoo, M.J. Amphibian Declines: The Conservation Status of United States Species; University of California Press: Berkeley, CA, USA, 2005. [Google Scholar]
- Poiani, K.A.; Johnson, W.C. Global warming and prairie wetlands: Potential consequences for waterfowl habitat. BioScience 1991, 41, 8. [Google Scholar] [CrossRef]
- Halpin, P.N. Global climate change and natural-area protection: Management responses and research directions. Ecol. Appl. 1997, 7, 828–843. [Google Scholar] [CrossRef]
- Daszak, P.; Scott, D.E.; Kilpatrick, A.M.; Faggioni, C.; Gibbons, J.W.; Porter, D. Amphibian population declines at Savannah River site are linked to climate, not chytridiomycosis. Ecology 2005, 86, 3232–3237. [Google Scholar] [CrossRef]
- Raxworthy, C.J. Extinction vulnerability of tropical montane endemism from warming and upslope displacement: A preliminary appraisal for the highest massif in Madagascar. Glob. Chang. Biol. 2008, 14, 1703–1720. [Google Scholar] [CrossRef]
- Janzen, F.J. Climate change and temperature-dependent sex determination in reptiles. Proc. Natl. Acad. Sci. USA 1994, 91, 7487–7490. [Google Scholar] [CrossRef] [PubMed]
- Pounds, A.J.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate warming and disease risks for terrestrial and marine biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Kubisch, A.; Holt, R.D.; Poethke, H.J.; Fronhofer, E.A. Where am I and why? Synthesizing range biology and the eco-evolutionary dynamics of dispersal. Oikos 2014, 123, 5–22. [Google Scholar] [CrossRef]
- Pough, F.H.; Andrews, R.M.; Cadle, J.E.; Crump, M.L.; Savitsky, A.H.; Wells, K.D. Herpetology, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Brown, W.S. Biology, Status, and Management of the Timber Rattlesnake; Society for the Study of Amphibians & Reptiles: Topeka, KS, USA, 1993. [Google Scholar]
- Sinervo, B.; Méndez-de-la-Cruz, F.; Miles, D.B.; Heulin, B.; Bastiaans, E.; Villagrán-Santa Cruz, M.; Lara-Resendiz, R.; Martínez-Méndez, N.; Calderón-Espinosa, M.L. Erosion of lizard diversity by climate change and altered thermal niches. Science 2010, 328, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Gibbon, J.W.; Scott, D.E.; Ryan, T.J.; Buhlmann, K.A.; Tuberville, T.D.; Metts, B.S. The global decline of reptiles, déjà vu amphibians. BioScience 2000, 50, 653–666. [Google Scholar] [CrossRef]
- Shine, R.; Harlow, P.S. Maternal manipulation of offspring phenotypes via nest-site selection in an oviparous lizard. Ecology 1996, 77, 1808–1817. [Google Scholar] [CrossRef]
- Shine, R. A new hypothesis for the evolution of viviparity in reptiles. Am. Nat. 1995, 145, 809–823. [Google Scholar] [CrossRef]
- Shine, R. Seasonal shifts in nest temperature can modify the phenotypes of hatchling lizards, regardless of overall mean incubation temperature. Funct. Ecol. 2004, 18, 43–49. [Google Scholar] [CrossRef]
- Valenzuela, N.; Lance, V.A. Temperature-Dependent Sex Determination in Vertebrates; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2005. [Google Scholar]
- Van Damme, R.; Bauwens, D.; Braña, F.; Verheyen, R.F. Incubation temperature differentially affects hatching time, egg survival, and hatchling performance in the lizard podarcis muralis. Herpetologica 1992, 48, 220–228. [Google Scholar]
- Rhen, T.; Lang, J.W. Phenotypic plasticity for growth in the common snapping turtle: Effects of incubation temperature, clutch, and their interaction. Am. Nat. 1995, 146, 726–747. [Google Scholar] [CrossRef]
- Turner, D.S.; Holm, P.A.; Wirt, E.B.; Schwalbe, C.R. Amphibians and reptiles of the Whetstone Mountains, Arizona. Southwest. Nat. 2003, 48, 347–355. [Google Scholar] [CrossRef]
- Flesch, A.D.; Swann, D.E.; Turner, D.S.; Powell, B.F. Herpetofauna of the Rincon Mountains, Arizona. Southwest. Nat. 2010, 55, 240–253. [Google Scholar] [CrossRef]
- Boeing, W.J.; Griffis-Kyle, K.L.; Jungels, J.M. Anuran habitat associations in the northern Chihuahuan desert, USA. J. Herpetol. 2014, 48, 103–110. [Google Scholar] [CrossRef]
- Harings, N.M.; Boeing, W.J. Desert anuran occurrence and detection in artificial breeding babitats. Herpetologica 2014, 70, 123–134. [Google Scholar] [CrossRef]
- Rogers, J.S. Species density and taxonomic diversity of Texas amphibians and reptiles. Syst. Biol. 1976, 25, 26–40. [Google Scholar] [CrossRef]
- Owen, J.G. Patterns of herpetofaunal species richness: Relation to temperature, precipitation, and variance in elevation. J. Biogeogr. 1989, 16, 141–150. [Google Scholar] [CrossRef]
- Bogosian, V.; Hellgren, E.C.; Moody, R.W. Assemblages of amphibians, reptiles, and mammals on an urban military base in Oklahoma. Southwest. Nat. 2012, 57, 277–284. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Reeder, T.W.; Cole, C.J.; Dessauer, H.C. Phylogenetic relationships of whiptail lizards of the genus Cnemidophorus (Squamata: Teiidae): A test of monophyly, reevaluation of Karyotypic evolution, and review of hybrid origins. Am. Mus. Novit. 2002, 3365, 1–61. [Google Scholar] [CrossRef]
- Lowe, C.J.; Wright, C.J.; Cole, C.J.; Bezy, R.L. Chromosomes and evolution of the species groups of Cnemidophorus (Reptilia: Teiidae). Syst. Biol. 1970, 19, 128–141. [Google Scholar] [CrossRef]
- Crother, B.I. Standard common and current scientific names for North American amphibians, turtles, reptiles, and crocodilians. Herpetol. Circ. 2012, 39, 1–92. [Google Scholar]
- Maslin, T.P. The sex of hatchlings of five apparently unisexual species of whiptail lizards (Cnemidophorus, Teiidae). Am. Midl. Nat. 1966, 76, 369–378. [Google Scholar] [CrossRef]
- Walker, J.M.; Taylor, H.L.; Cordes, J.E.; Manning, G.J.; Cole, C.J. Comparative Meristic Variability in Whiptail Lizards (Teiidae, Aspidoscelis): Samples of Parthenogenetic A. tesselata Versus Samples of Sexually Reproducing A. sexlineata, A. marmorata, and A. gularis septemvittata; American Museum Novitates, No. 3744; American Museum of Natural History: New York, NY, USA, 2012. [Google Scholar]
- Maslin, T.P.; Secoy, D.M. A Checklist of the Lizard Genus Cnemidophorus (Teiidae), 1st ed.; University of Colorado Museum: Boulder, CO, USA, 1986. [Google Scholar]
- Peterson, A.T.; Soberon, J.; Pearson, G.; Anderson, R.P. Ecological Niches and Geographic Distributions (MPB-49); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- South Central Climate Science Center South-Central U.S. Available online: http://www.southcentralclimate.org/index.php/pages/scus (accessed on 16 December 2016).
- Arizona Game and Fish Department—Wildlife. Available online: https://www.azgfd.com/Wildlife/HeritageFund (accessed on 16 December 2016).
- Colorado Natural Heritage Program. Available online: http://www.cnhp.colostate.edu (accessed on 16 December 2016).
- Natural Heritage New Mexico. Available online: https://nhnm.unm.edu (accessed on 16 December 2016).
- Texas Parks and Wildlife Department, Wildlife Diversity. Available online: http://tpwd.texas.gov/huntwild/wild/wildlife_diversity (accessed on 16 December 2016).
- Biodiversity Information Serving Our Nation. Available online: https://bison.usgs.gov/#home (accessed on 16 December 2016).
- Herpetology Network. Available online: http://www.herpnet.org (accessed one 16 December 2016).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Leathwick, J.R.; Elith, J.; Hastie, T. Comparative performance of generalized additive models and multivariate adaptive regression splines for statistical modelling of species distributions. Ecol. Model. 2006, 199, 188–196. [Google Scholar] [CrossRef]
- Talbert, C. Software for Assisted Habitat Modeling Package for VisTrails (SAHM: VisTrails) v.1; U.S. Geological Survey: Fort Collins, CO, USA, 2012.
- Morisette, J.T.; Jarnevich, C.S.; Holcombe, T.R.; Talbert, C.B.; Ignizio, D.I.; Talbert, M.K.; Silva, C.; Koop, D.; Swanson, A.; Young, N.E. VisTrails SAHM: Visualization and workflow management for species habitat modeling. Ecography 2013, 36, 129–135. [Google Scholar] [CrossRef]
- National Gap Analysis Program. Available online: https://gapanalysis.usgs.gov (accessed on 16 December 2016).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carre, G. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Gama, M.; Crespo, D.; Dolbeth, M.; Anastácio, P. Predicting global habitat suitability for Corbicula fluminea using species distribution models: The importance of different environmental datasets. Ecol. Model. 2016, 319, 163–169. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Ma, P.; Kumar, S.; Rocca, M.; Morisette, J.T.; Jarnevich, C.S.; Benson, N. Ensemble habitat mapping of invasive plant species. Risk Anal. Off. Publ. Soc. Risk Anal. 2010, 30, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Manel, S.; Williams, S.C.; Ormerod, S.J. Evaluating presence–absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Crookston, N.L.; Sáenz-Romero, C.; Campbell, E.M. North American vegetation model for land-use planning in a changing climate: A solution to large classification problems. Ecol. Appl. Publ. Ecol. Soc. Am. 2012, 22, 119–141. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1998, 240, 1285–1293. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2011, 93, 485–498. [Google Scholar] [CrossRef]
- Sheffield, J.; Barrett, A.; Colle, B.; Fu, R.; Geil, K.L.; Hu, Q.; Kinter, J.; Kumar, S.; Langenbrunner, B.; Lombardo, K.; et al. North American climate in CMIP5 experiments. Part I: Evaluation of historical simulations of continental and regional climatology. J. Clim. 2013, 26, 9209–9245. [Google Scholar] [CrossRef]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Collins, W.J.; Bellouin, N.; Doutriaux-Boucher, M.; Gedney, N.; Halloran, P.; Hinton, T.; Hughes, J.; Jones, C.D.; Joshi, M.; Liddicoat, S.; et al. Development and evaluation of an Earth-System model—HadGEM2. Geosci. Model Dev. 2011, 4, 1051–1075. [Google Scholar] [CrossRef]
- Watanabe, M.; Suzuki, T.; O’Ishi, R.; Komuro, Y.; Watanabe, S.; Emori, S.; Takemura, T.; Chikira, M.; Ogura, T.; Sekiguchi, M.; et al. Improved climate simulation by MIROC5: Mean states, variability, and climate sensitivity. J. Clim. 2010, 23, 6312–6335. [Google Scholar] [CrossRef]
- Block, K.; Mauritsen, T. Forcing and feedback in the MPI-ESM-LR coupled model under abruptly quadrupled CO2. J. Adv. Model. Earth Syst. 2013, 5, 676–691. [Google Scholar] [CrossRef]
- Roeckner, E.; Giorgetta, M.A.; Crueger, T.; Esch, M.; Pongratz, J. Historical and future anthropogenic emission pathways derived from coupled climate–carbon cycle simulations. Clim. Chang. 2011, 105, 91–108. [Google Scholar] [CrossRef]
- Arora, V.K.; Scinocca, J.F.; Boer, G.J.; Christian, J.R.; Denman, K.L.; Flato, G.M.; Kharin, V.V.; Lee, W.G.; Merryfiel, W.G. Carbon emission limits required to satisfy future representative concentration pathways of greenhouse gases. Geophys. Res. Lett. 2011, 38, L05805. [Google Scholar] [CrossRef]
- Chalmers, N.; Highwood, E.J.; Hawkins, E.; Sutton, R.; Wilcox, L.J. Aerosol contribution to the rapid warming of near-term climate under RCP 2.6. Geophys. Res. Lett. 2012, 39, L18709. [Google Scholar] [CrossRef]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5. [Google Scholar] [CrossRef]
- Knutti, R.; Sedláček, J. Robustness and uncertainties in the new CMIP5 climate model projections. Nat. Clim. Chang. 2013, 3, 369–373. [Google Scholar] [CrossRef]
- Edenhofer, O.; Pichs-Madruga, R.; Sokona, Y.; Farahani, E. PCC 2014: Summary for policymakers. In Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2014. [Google Scholar]
- Rosenzweig, C.; Karoly, D.; Vicarelli, M.; Neofotis, P.; Wu, Q.; Casassa, G.; Menzel, A.; Root, T.L.; Estrella, N.; Seguin, B.; et al. Attributing physical and biological impacts to anthropogenic climate change. Nature 2008, 453, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.E.; Bolitho, E.E.; Fox, S. Climate change in Australian tropical rainforests: An impending environmental catastrophe. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Willette, D.A.; Tucker, J.K.; Janzen, F.J. Linking climate and physiology at the population level for a key life-history stage of turtles. Can. J. Zool. 2005, 83, 845–850. [Google Scholar] [CrossRef]
- Wapstra, E.; Uller, T.; Sinn, D.L.; Olsson, M.; Mazurek, K.; Joss, J.; Shine, R. Climate effects on offspring sex ratio in a viviparous lizard. J. Anim. Ecol. 2009, 78, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Stenseth, N.C.; Mysterud, A. Climate, changing phenology, and other life history traits: Nonlinearity and match–mismatch to the environment. Proc. Natl. Acad. Sci. USA 2002, 99, 13379–13381. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.E.; Both, C.; Lambrechts, M.M. Global climate change leads to mistimed avian reproduction. Adv. Ecol. Res. 2004, 35, 89–110. [Google Scholar]
- Hammerson, G.A. Amphibians and Reptiles in Colorado; University Press of Colorado: Boulder, CO, USA, 2000. [Google Scholar]
- Degenhardt, W.G.; Painter, C.W.; Price, A.H. Amphibians and Reptiles of New Mexico; University of New Mexico Press: Albuquerque, NM, USA, 2005. [Google Scholar]
- Dixon, J.R. Amphibians and Reptiles of Texas, 2nd ed.; Texas A&M University Press: College Station, TX, USA, 2000. [Google Scholar]
- Parker, E.D.; Selander, R.K. Low clonal diversity in the parthenogenetic lizard Cnemidophorus neomexicanus (Sauria: Teiidae). Herpetologica 1984, 40, 245–252. [Google Scholar]
- Walker, J.M.; Cordes, J.E.; Taylor, H.L. Parthenogenetic Cnemidophorus tesselatus complex (Sauria: Teiidae): A neotype for Diploid C. tesselatus (Say, 1823), redescription of the taxon, and description of a new triploid species. Herpetologica 1997, 53, 233–259. [Google Scholar]
- Stebbins, R.C. A Field Guide to Western Reptiles and Amphibians, 3rd ed.; Houghton Mifflin Harcourt: Boston, MA, USA, 2003. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org (accessed on 16 December 2016).
- Taylor, H.L.; Cole, C.J.; Hardy, L.M.; Cordes, J.E.; Walker, J.M.; Dessauer, H.C. Natural Hybridization between the Teiid Lizards Cnemidophorus tesselatus (parthenogenetic) and C. tigris marmoratus (bisexual): Assessment of Evolutionary Alternatives; American Museum Novitates, No. 3345; American Museum of Natural History: New York, NY, USA, 2001. [Google Scholar]
- Cole, C.J.; Painter, C.W.; Dessauer, H.C.; Taylor, H.L. Hybridization between the Endangered Unisexual Gray-Checkered Whiptail Lizard (Aspidoscelis dixoni) and the Bisexual Western Whiptail Lizard (Aspidoscelis tigris) in Southwestern New Mexico; American Museum Novitates, No. 3555; American Museum of Natural History: New York, NY, USA, 2007. [Google Scholar]
- Walker, J.M.; Lemos-Espinal, J.A.; Cordes, J.E.; Taylor, H.L.; Smith, H.M. Allocation of populations of whiptail lizards to septemvittatus Cope, 1892 (Genus Cnemidophorus) in Chihuahua, México, and the scalaris problem. Copeia 2001, 2001, 747–765. [Google Scholar] [CrossRef]
- Collins, J.T.; Conant, R. A Field Guide to Reptiles and Amphibians: Eastern and Central North America, 3rd ed.; Houghton Mifflin Harcourt: Boston, MA, USA, 1998. [Google Scholar]
- Bartlett, R.D.; Barlett, P.P. A Field Guide to Florida Reptiles and Amphibians; Gulf Publishing Company: Houston, TX, USA, 1999. [Google Scholar]
- Walker, J.M. The taxonomy of parthenogenetic species of hybrid origin: Cloned hybrid populations of Cnemidophorus (Sauria: Teiidae). Syst. Zool. 1986, 35, 427–440. [Google Scholar] [CrossRef]
- Forstner, M.R.J.; Dixon, J.R.; Forstner, J.M.; Davis, S.K. Apparent hybridization between Cnemidophorus gularis and Cnemidophorus septemvittatus from an area of sympatry in southwest Texas. J. Herpetol. 1998, 32, 418–425. [Google Scholar] [CrossRef]
- Committee on Standard English and Scientific. Scientific and Standard English Names of Amphibians and Reptiles of North America North of Mexico, with Comments Regarding Confidence in Our Understanding, 5th ed.; Society for the Study of Amphibians and Reptiles: Topeka, KS, USA, 2000. [Google Scholar]
- Scudday, J.F. A new species of lizard of the Cnemidophorus tesselatus group from Texas. J. Herpetol. 1973, 7, 363–371. [Google Scholar] [CrossRef]
- Painter, C.W. Status of the Gray-Checkered Whiptail (Cnemidophorus dixoni) on Bureau of Land Management Lands in Southwestern New Mexico; New Mexico Department of Game and Fish: Santa Fe, NM, USA, 1995. [Google Scholar]
- New Mexico Game and Fish. Available online: http://www.wildlife.state.nm.us (accessed on 16 December 2016).
- Zweifel, R.G. Variation in and Distribution of the Unisexual Lizard, Cnemidophorus tesselatus; American Museum Novitates, No. 2235; American Museum of Natural History: New York, NY, USA, 1965. [Google Scholar]
- Walker, J.M.; Taylor, H.L.; Cordes, J.E. Hybrid Cnemidophorus (Sauria: Teiidae) in Ninemile Valley of the Purgatoire River, Colorado. Southwest. Nat. 1994, 39, 235–240. [Google Scholar] [CrossRef]
- Persons, T.; Wright, J.W. Discovery of Cnemidophorus neomexicanus in Arizona. Herpetol. Rev. 1999, 30, 3. [Google Scholar]
- Taylor, H.L.; Parker, E.D.; Dessauer, H.C.; Cole, C.J. Congruent Patterns of Genetic and Morphological Variation in the Parthenogenetic lizard Aspidoscelis tesselata (Squamata, Teiidae) and the Origins of Color Pattern Classes and Genotypic Clones in Eastern New Mexico; American Museum Novitates, No. 3424; American Museum of Natural History: New York, NY, USA, 2003. [Google Scholar]
- Taylor, H.L.; Walker, J.M.; Cordes, J.E.; Manning, G.J. Application of the evolutionary species concept to parthenogenetic entities: Comparison of postformational divergence in two clones of Aspidoscelis tesselata and between Aspidoscelis cozumela and Aspidoscelis maslini (Squamata: Teiidae). J. Herpetol. 2005, 39, 266–277. [Google Scholar] [CrossRef]
- Bateman, B.L.; Murphy, H.T.; Reside, A.E.; Mokany, K.; VanDerWal, J. Appropriateness of full-, partial- and no-dispersal scenarios in climate change impact modelling. Divers. Distrib. 2013, 19, 1224–1234. [Google Scholar] [CrossRef]
- Salas, E.A.L.; Seamster, V.A.; Boykin, K.G.; Harings, N.M.; Dixon, K.W. Modeling the impacts of climate change on Species of Concern (birds) in South Central U.S. based on bioclimatic variables. AIMS Environ. Sci. 2017, 4, 358–385. [Google Scholar] [CrossRef]
- iNaturalist. Available online: https://www.inaturalist.org (accessed on 7 April 2017).











| Variable | Description | Temporal Scale |
|---|---|---|
| Bioclim 1 | Annual Mean Temperature | Annual |
| Bioclim 2 | Mean Diurnal Range | Variation |
| Bioclim 3 | Isothermality | Variation |
| Bioclim 4 | Temperature Seasonality | Variation |
| Bioclim 5 | Maximum Temperature of the Warmest Month | Month |
| Bioclim 6 | Minimum Temperature of the Coldest Month | Month |
| Bioclim 7 | Temperature Annual Range | Annual |
| Bioclim 8 | Mean Temperature of Wettest Quarter | Quarter |
| Bioclim 9 | Mean Temperature of Driest Quarter | Quarter |
| Bioclim 10 | Mean Temperature of Warmest Quarter | Quarter |
| Bioclim 11 | Mean Temperature of Coldest Quarter | Quarter |
| Bioclim 12 | Annual Precipitation | Annual |
| Bioclim 13 | Precipitation of Wettest Month | Month |
| Bioclim 14 | Precipitation of Driest Month | Month |
| Bioclim 15 | Precipitation Seasonality | Variation |
| Bioclim 16 | Precipitation of Wettest Quarter | Quarter |
| Bioclim 17 | Precipitation of Driest Quarter | Quarter |
| Bioclim 18 | Precipitation of Warmest Quarter | Quarter |
| Bioclim 19 | Precipitation of Coldest Quarter | Quarter |
| Species | GLM | MARS | BRT | RF | Maxent | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| AUC | %Co | AUC | %Co | AUC | %Co | AUC | %Co | AUC | %Co | |
| A. tesselata | 0.77 | 69.4 | 0.78 | 71.8 | 0.94 | 86.3 | 0.80 | 72.3 | 0.83 | 75.6 |
| A. tigris | 0.77 | 69.6 | 0.79 | 71.2 | 0.89 | 80.4 | 0.82 | 75.0 | 0.80 | 73.3 |
| A. neotesselata | NA | NA | 0.76 | 65.3 | NA | NA | NA | NA | 0.77 | 67.5 |
| A. scalaris | NA | NA | 0.78 | 78.4 | NA | NA | 0.65 | 63.1 | 0.58 | 61.8 |
| A. gularis | 0.55 | 49.5 | 0.66 | 61.7 | 0.87 | 78.3 | 0.71 | 66.8 | 0.70 | 63.7 |
| A. neomexicana | 0.77 | 66.5 | 0.80 | 70.0 | 0.94 | 85.9 | 0.81 | 76.8 | 0.81 | 72.2 |
| A. dixoni | 0.81 | 77.9 | 0.76 | 68.4 | NA | NA | 0.76 | 74.7 | NA | NA |
| Species | GLM | MARS | BRT | RF | Maxent |
|---|---|---|---|---|---|
| TSS | TSS | TSS | TSS | TSS | |
| A. tesselata | 0.41 | 0.43 | 0.73 | 0.44 | 0.51 |
| A. tigris | 0.39 | 0.42 | 0.61 | 0.51 | 0.47 |
| A. neotesselata | NA | 0.37 | NA | NA | 0.12 |
| A. scalaris | NA | 0.36 | NA | 0.27 | 0.22 |
| A. gularis | 0.05 | 0.24 | 0.56 | 0.33 | 0.28 |
| A. neomexicana | 0.33 | 0.43 | 0.72 | 0.53 | 0.46 |
| A. dixoni | 0.58 | 0.14 | NA | 0.73 | NA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, G.; Salas, E.A.L.; Harings, N.M.; Boykin, K.G. Projections of Future Suitable Bioclimatic Conditions of Parthenogenetic Whiptails. Climate 2017, 5, 34. https://doi.org/10.3390/cli5020034
Alvarez G, Salas EAL, Harings NM, Boykin KG. Projections of Future Suitable Bioclimatic Conditions of Parthenogenetic Whiptails. Climate. 2017; 5(2):34. https://doi.org/10.3390/cli5020034
Chicago/Turabian StyleAlvarez, Guillermo, Eric Ariel L. Salas, Nicole M. Harings, and Kenneth G. Boykin. 2017. "Projections of Future Suitable Bioclimatic Conditions of Parthenogenetic Whiptails" Climate 5, no. 2: 34. https://doi.org/10.3390/cli5020034
APA StyleAlvarez, G., Salas, E. A. L., Harings, N. M., & Boykin, K. G. (2017). Projections of Future Suitable Bioclimatic Conditions of Parthenogenetic Whiptails. Climate, 5(2), 34. https://doi.org/10.3390/cli5020034